5种川西北沙化地草本植物生态适应策略的差异性
2016-06-14曾晓琳游明鸿刘金平
李 莹,曾晓琳,游明鸿,刘金平,蔡 捡
(1.西华师范大学生命科学院,四川 南充 637009; 2.四川省草原科学研究院,四川 成都 611731)
5种川西北沙化地草本植物生态适应策略的差异性
李 莹1,曾晓琳1,游明鸿2,刘金平1,蔡 捡1
(1.西华师范大学生命科学院,四川 南充 637009; 2.四川省草原科学研究院,四川 成都 611731)
摘要:以5种川西北沙化草地自然留存植物沙生苔草(Carex praeclara)、赖草(Leymus secalinus)、紫花列当(Orobanche coerulescens)、藏茴香(Carum carvi)和地八角(Astragalus bhotanensis)为研究对象,通过测定5种草地植物的表观性状、构件含水量和生物量及其分配,分析5种植物的生长发育、资源分配对沙化生境的响应差异,探讨5种植物对沙化地生境的应对策略与适应机理。结果表明,1)5种植物的表观性状具有极显著差异(P<0.01),沙生苔草通过冠幅,赖草通过根茎,地八角通过深根系,紫花列当和藏茴香通过扩大有性生殖来适应沙化生境。2)5种植物的含水量及其配比均存在极显著差异(P<0.01),沙生苔草、赖草和地八角的水分优先分配到生殖枝,紫花列当优先分配到茎叶,藏茴香则采用相对均衡的水分分配方式。3)5种植物的生物量及其分配也存在极显著差异(P<0.01),沙生苔草生物量优先分配到根茎,赖草优先分配到地上茎叶,紫花列当优先分配到生殖枝,地八角优先分配到根部,而藏茴香则相对均衡的分配生物量到各构件。5种沙生植物对资源权衡分配的方式不同,但均能通过表型可塑性、水分利用能力和生物量分配等应对策略来获取或分配有限的可利用资源,增加其对生境的适应性,得以在沙化草地中留存下来。
关键词:沙化草地;沙生植物;表观性状;水分分配;适应策略
川西北高寒草原地处青藏高原东缘,是全国五大牧区之一,草地面积820余万hm2[1]。随着全球气候变暖及人类掠夺式放牧、滥垦和乱挖,致使草原植被覆盖度急剧下降,生态系统极度恶化,退化沙化草地面积逐年扩大,形成斑块状固定、半固定和流动沙丘,川西北高寒草原生态系统的自调节功能严重丧失[2]。近年来,虽通过种植高山柳(Salixcupularis)和沙棘(Hippophaerhamnoides)等本土灌木作为生物围栏和防风林,并结合燕麦(Avenasativa)、披碱草(Elymusdahuricus)等优良草种来进行防沙治沙和恢复植被[3],但由于所选草本植物生态适应性差,植株存活率和植被保存率极低[4],3~5年后仅剩灌木,引进草本植物荡然无存。因此,筛选当地野生耐沙抗沙草种,进行合理驯化与利用是该区防沙治沙必由之路。
在沙化过程中通过适者生存的自然选择,高寒草原留存有多种适应沙化环境的野生植物,这些植物长期经受高寒缺氧、强烈太阳辐射、大量紫外线、低CO2以及急剧气候变化的严重胁迫[5],从生理、形态、能量分配等方面形成一系列生态适应对策[6]。诸多学者对沙化草地植物群落[7-8]、抗逆生理[9]、水分利用[10-11]、生物量分配[12]等方面进行了研究,如王钰和宝音陶格涛[8]研究了荒漠草原沙生针茅(Stipaglareosa)植物群落的组成及生物量结构;左力翔等[11]研究了小叶杨(Populussimonii)不同树龄的水分利用策略;周瑞莲等[13]研究了4种沙生植物的生理调控机制及适应对策。但对川西北沙化草地自然存留植物生态适应策略[14-15]的报道较少。筛选出适应能力强、分生再生能力好的本土植物,对解决该区生态治理植物材料极度缺乏[16]的问题具有极其重要的现实意义。本研究选取沙生苔草(Carexpraeclara)、赖草(Leymussecalinus)、紫花列当(Orobanchecoerulescens)、藏茴香(Carumcarvi)和地八角(Astragalusbhotanensis)5种常见的沙化草地留存植物为试验材料,通过测定生长特征、表观性状、水分含量和生物量分配等指标,分析5种沙生植物的形态结构、水分和生物量及其分配对沙化生境的适应性差异,探讨不同沙生植物的生态适应策略,旨在研究不同沙生植物对沙化地生境的应对策略与适应机理,为川西北高寒草原地区防风治沙植物的选择利用提供依据。
1材料与方法
1.1试验地概况
研究于四川省阿坝州阿坝县贾洛乡沙化地进行,地理坐标为33°10.725′ N,102°37.170′ E,海拔3 462 m,属高原寒温带半湿润季风气候,年平均气温3.3 ℃,1月平均气温7.9 ℃,7月平均气温12.5 ℃,≥10 ℃的年活动积温788.0 ℃·d,持续天数63 d;年均降水量717.8 mm,降水集中在5―9月,占全年降水量的81%[17],全年无绝对无霜期,相对无霜期33 d。
试验区内分布着3年前人工种植株距10~12 m的高山柳,以该灌木为中心演替出呈斑块状规律性分布的沙生植物,距高山柳0~2 m(A)分布有紫花列当,2~4 m(B)分布有藏茴香和地八角,4~6 m(C)分布有苔草和赖草。样点生境如表1。
1.2试验方法
于2014年7月,在沙化地内随机选取高山柳为圆心,以距高山柳0~2 m(A)、2~4 m(B)、4~6 m(C)环形范围为样圆,在各样圆上随机选取目标植物各5株,连根挖取备用。设10次重复。
1.3测定指标
将挖取的植物清洗、拭干,用直尺测植株最大根长、根茎长、地上茎长、花穗柄长、花穗轴长、根幅和冠幅后,将花、茎叶、根茎和根分离称鲜重,分别装袋,105 ℃杀青0.5 h后,在80 ℃下烘至恒重,称干重为生物量。计算下列参数:
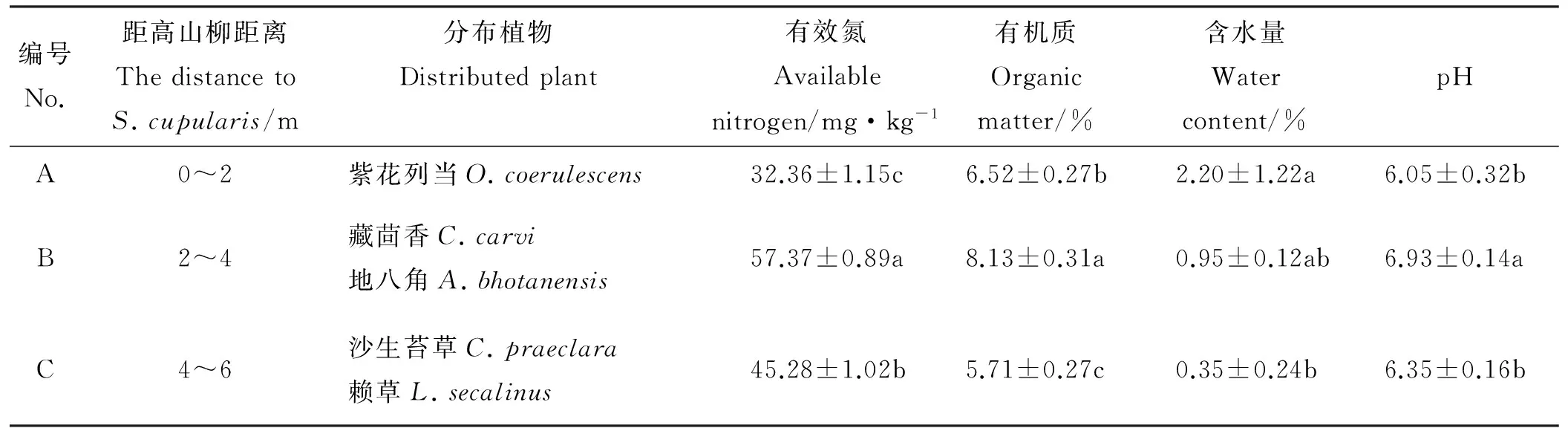
表1 采样地生境概况
注:同列不同小写字母表示不同样地间差异显著(P<0.05)。
Note: Different lower case letters within the same column indicate significant difference among different samplings at 0.05 level.
构件含水量=(鲜重-干重)/鲜重×100%;
构件水分配比=构件含水量/总含水量×100%;
构件生物量配比=构件生物量/总生物量×100%;
根冠比=(根+根茎)生物量/(地上茎+生殖枝)生物量;
根冠水分比=(根+根茎)含水量/(地上茎+生殖枝)含水量;
根茎的比茎长(cm·g-1)=根茎长/根茎生物量。
1.4数据分析
采用SPSS 19.0对数据进行方差分析(ANOVA)和多重比较(SNK)。
2结果与分析
2.1表观性状差异
5种沙生植物根、茎、花等构件的表观性状有极显著差异(P<0.01)(表2)。多重比较表明,地八角最大根长和根幅显著大于其它植物(P<0.05),分别为1 059.97和239.33cm;沙生苔草次之,显著大于赖草、藏茴香和紫花列当,紫花列当最小。5种植物茎性状均具有显著差异(P<0.05),其根茎长大小顺序为赖草>紫花列当>地八角>沙生苔草(藏茴香不具根茎),地上茎长大小顺序为赖草>沙生苔草>藏茴香>地八角>紫花列当。5种植物花性状均有显著差异,藏茴香花穗柄长显著大于其它植物(P<0.05),为35.43 cm,随后依次为沙生苔草>赖草>地八角>紫花列当;紫花列当花穗轴长与藏茴香差异不大,但均显著大于沙生苔草、赖草和地八角,赖草花穗轴长略大于沙生苔草,地八角花穗轴长最小。沙生苔草冠幅显著大于其它4种植物,其它植物冠幅大小顺序为藏茴香>赖草>紫花列当>地八角。
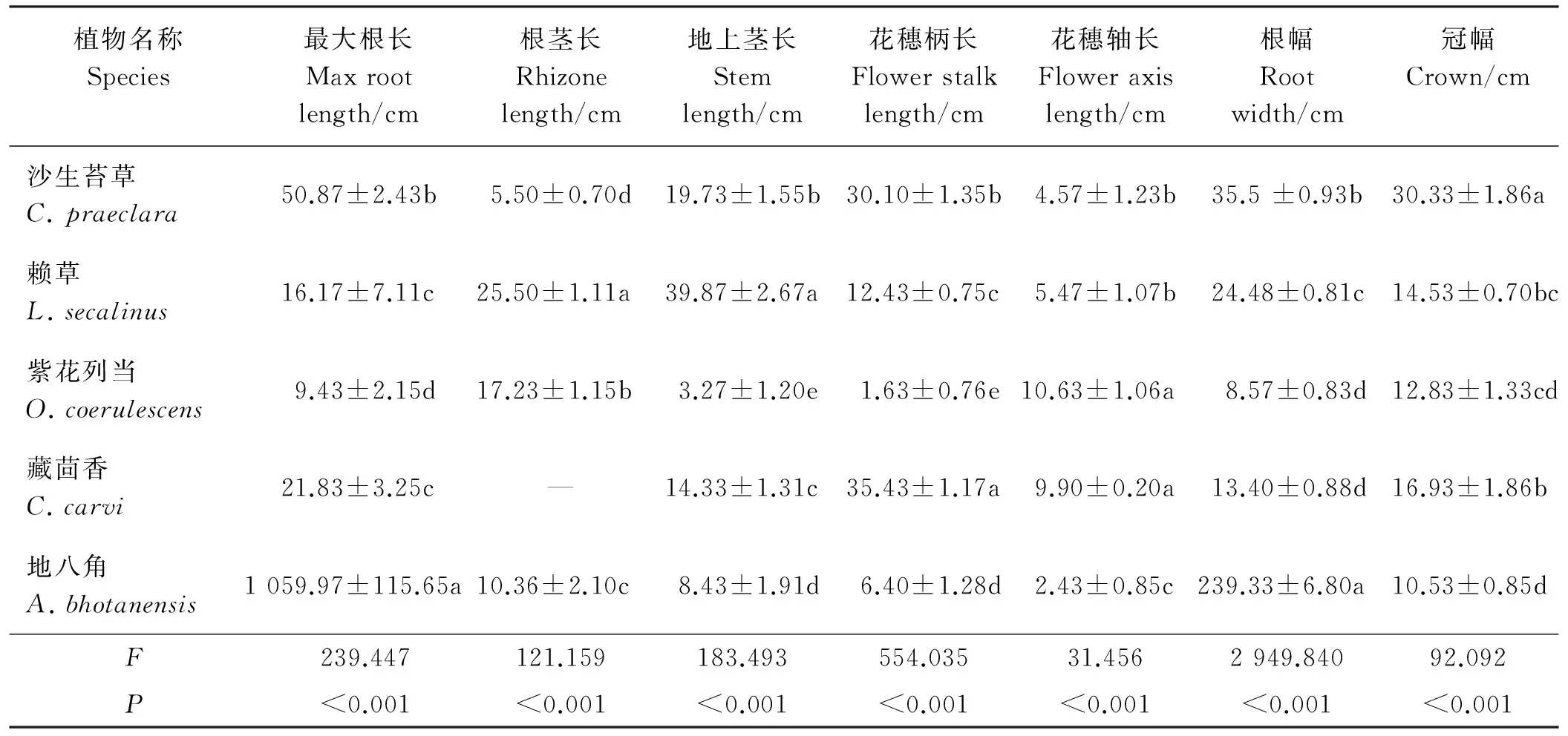
表2 5种沙生植物各构件表观性状差异分析
注:同列不同小写字母表示不同植物间差异显著(P<0.05)。下同。
Note: Different lower case letters within the same column indicate significant difference among different species. The same below.
2.2含水量差异
2.2.1单株含水量差异5种植物单株含水量存在着极显著差异(P<0.01)(表3)。单株含水量高低顺序均表现为紫花列当>藏茴香>地八角>沙生苔草>赖草。紫花列当单株含水量高达67.23%,显著高于其它4种植物(P<0.05),而沙生苔草和赖草单株含水量较低,约为紫花列当的40%。
2.2.2构件含水量差异5种植物构件含水量均存在极显著差异(P<0.01),F值显示,构件含水差异为根茎>根>茎叶>生殖枝。沙生苔草、赖草和地八角的生殖枝含水量均高于营养器官。沙生苔草的茎叶含水量次之,根茎含水量略大于根含水量;赖草根茎含水量高于茎叶含水量和根含水量,根含水量最低;地八角茎叶含水量稍高于根含水量,两者均高于根茎含水量。紫花列当和藏茴香茎叶含水量均最高,紫花列当各构件含水量高低顺序为地上茎叶>根茎>根>生殖枝;藏茴香没有根茎,其根含水量和生殖枝含水量相差甚微,仅略小于茎叶含水量。
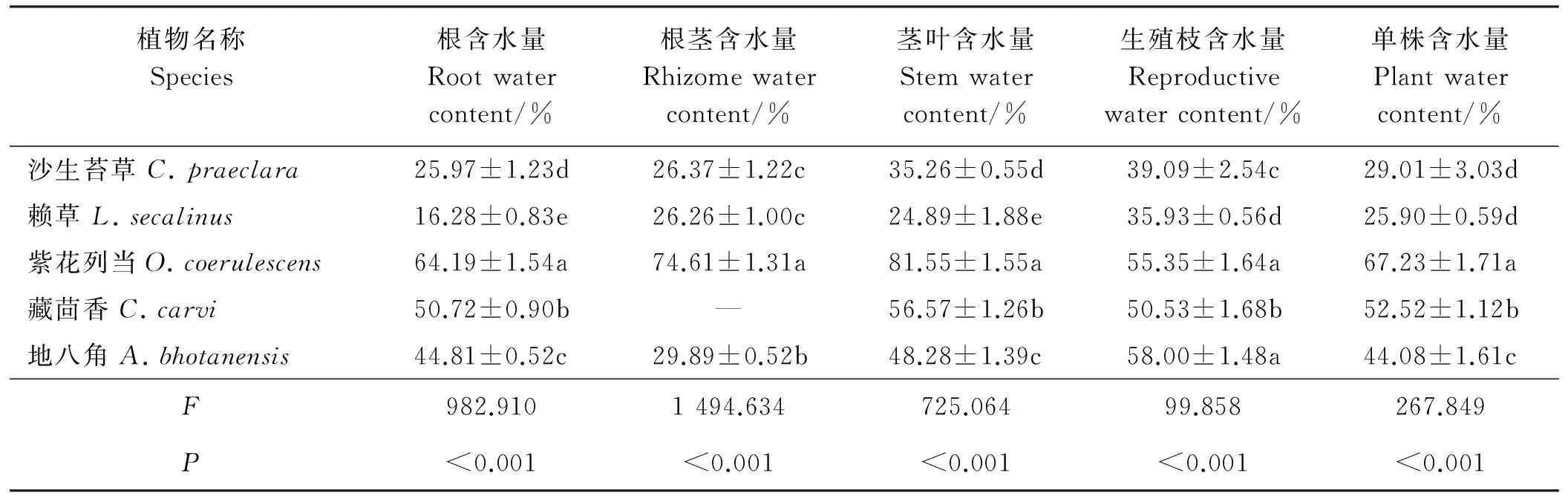
表3 5种沙生植物各构件含水量差异分析
2.2.3同一构件含水量差异同一构件含水量在不同植物间均有极显著差异(P<0.01)。5种植物根含水均有极显著差异(P<0.05),变化范围为16.28%~64.19%,由高到低顺序为紫花列当>藏茴香>地八角>沙生苔草>赖草,其中紫花列当根含水量是赖草的3.94倍。根茎含水量顺序为紫花列当>地八角>沙生苔草>赖草,沙生苔草和赖草间无显著差异(P>0.05),仅约为紫花列当根茎含水量的35%。地上茎叶含水量在不同植物间均存在显著差异,与根含水量大小顺序一致。紫花列当和地八角生殖枝含水量差异较小,但均显著高于其它植物(P<0.05),藏茴香、沙生苔草和赖草间生殖枝含水量也有显著差异,赖草最小。
2.3水分分配差异
2.3.1根冠水分比差异5种植物间根冠水分比存在极显著差异(P<0.01)(表4),紫花列当根冠水分比最大,为1.01 g·g-1,沙生苔草、赖草和地八角之间无明显差异(P>0.05),但均显著高于藏茴香(P<0.05),藏茴香根冠水分比仅为紫花列当的46.53%。
2.3.2构件水分分配差异构件水分分配在5种沙生植物间均有极显著差异(P<0.01),F值显示,构件水分分配差异大小为根茎>根>地上茎叶>生殖枝。沙生苔草、赖草和地八角水分优先分配到生殖枝,紫花列当则优先分配到茎叶,藏茴香则相对均衡的分配水分。
2.3.3同一构件水分分配差异同一构件水分分配在不同植物间也有显著差异(P<0.05)。5种植物的地下构件水分配比差异显著(P<0.05),根水分配比顺序为藏茴香>地八角>紫花列当>沙生苔草>赖草,藏茴香根水分配比是赖草的2.04倍;根茎水分配比顺序为紫花列当>赖草>沙生苔草>地八角。地上构件水分配比在不同植物间有显著差异,地上茎叶水分配比高低顺序为藏茴香>紫花列当>沙生苔草>地八角>赖草,赖草生殖枝水分配比明显高于其它4种植物,为34.79%。
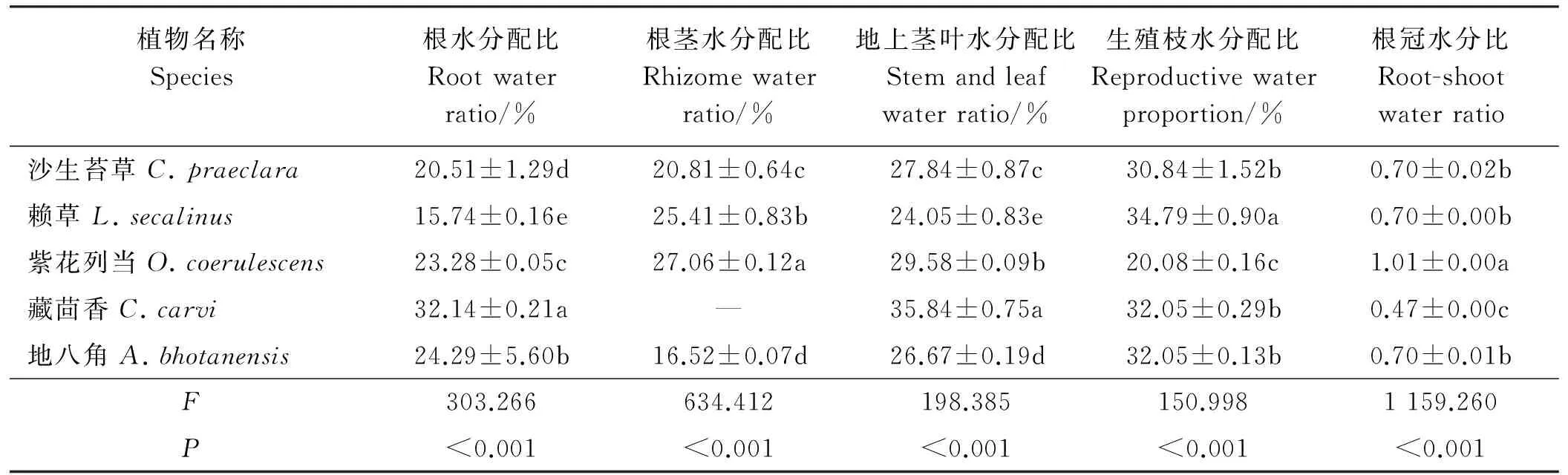
表4 5种沙生植物各构件水分分配差异分析
2.4生物量累积的差异
2.4.1单株生物量差异5种植物单株生物量存在极显著差异(P<0.01)(表5),大小顺序为沙生苔草>地八角>紫花列当>赖草>藏茴香。沙生苔草单株生物量高达31.25 g,显著高于其它植物(P<0.05),藏茴香单株生物量最小,沙生苔草是其生物量的18.83倍。
2.4.2构件生物量差异5种沙生植物构件生物量均具有极显著差异(P<0.01)。F值显示,构件生物量差异顺序为根>根茎>地上茎叶>生殖枝。同一植物不同构件生物量存在明显差异(P<0.05),沙生苔草单株根茎生物量最大,为15.01 g,单株地上茎叶生物量也最大,为6.83 g,紫花列当单株生殖生物量最大,为6.87 g,地八角单株根生物量均最大,为18.16 g。
2.4.3同一构件生物量差异不同植物同一构件比较可知,地八角根单株生物量显著大于其它植物(P<0.05),沙生苔草单株根生物量显著大于紫花列当,也均显著大于赖草和藏茴香,赖草和藏茴香的单株根生物量相差不显著(P<0.05)。单株根茎生物量大小顺序为沙生苔草>紫花列当>地八角>赖草,地上茎叶生物量大小顺序为沙生苔草>地八角>赖草>紫花列当>藏茴香。单株生殖生物量以紫花列当最大,显著大于其它植物,沙生苔草、赖草、藏茴香和地八角单株生殖生物量没有明显差异。
2.5生物量分配的差异
2.5.1根冠比和根茎比茎长差异5种植物根冠比、根茎比茎长均表现出极显著差异(P<0.01)(表6)。地八角根冠比最大,沙生苔草次之,根冠比值均大于1,显著大于赖草、紫花列当和藏茴香(P<0.05),后3种植物根冠比相差不显著(P>0.05)。赖草的根茎比茎长达29.31cm·g-1,显著高于其它3种植物(P<0.05),紫花列当和地八角根茎比茎长高于沙生苔草,沙生苔草仅为赖草的1.3%。可见,沙生苔草和地八角生物量主要投资于地下部分,赖草、紫花列当和藏茴香生物量则主要投资于地上茎叶和生殖枝。
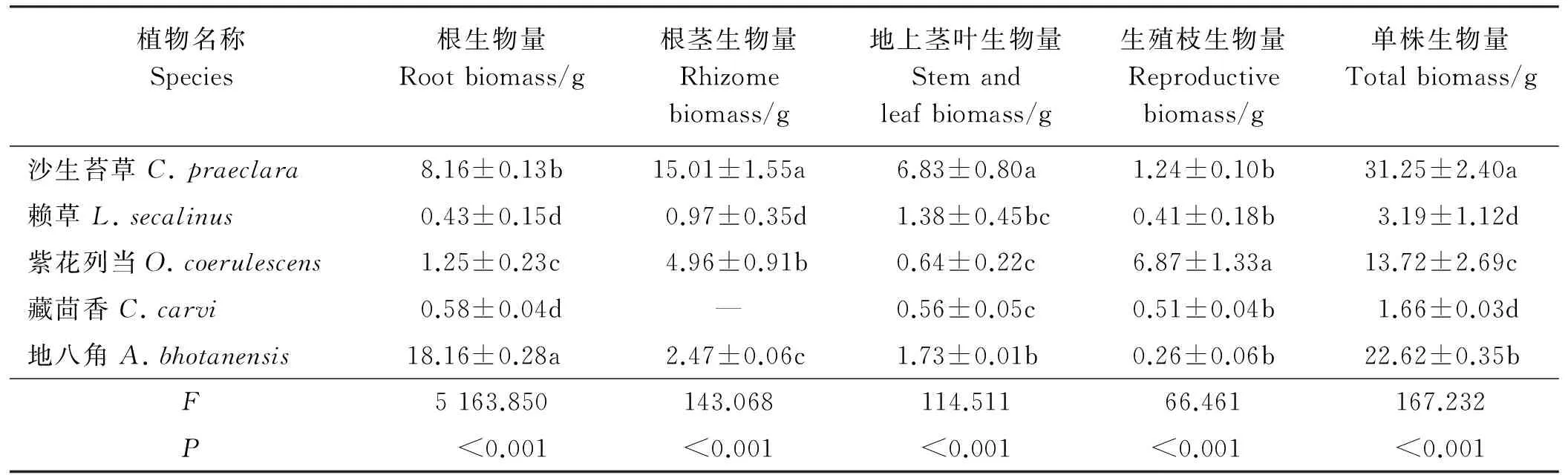
表5 5种沙生植物各构件单株生物量差异分析
2.5.2生物量分配比差异5种植物构件生物量分配比有极显著差异(P<0.01),F值显示,差异大小顺序为根>根茎>生殖枝>茎叶。根生物量配比变化范围为9.10%~80.28%,大小表现为地八角>藏茴香>沙生苔草>赖草>紫花列当,地八角是紫花列当的8.82倍;根茎生物量配比大小表现为沙生苔草>紫花列当>赖草>地八角;地上茎叶生物量配比大小表现为赖草>藏茴香>沙生苔草>地八角>紫花列当;生殖配比大小表现为紫花列当>藏茴香>赖草>沙生苔草>地八角,紫花列当生殖配比高达50.14%,沙生苔草仅为它的8%左右。可见,沙生苔草生物量优先向根茎分配,赖草优先向地上茎叶分配,紫花列当优先向生殖枝分配,地八角则优先向根部分配,藏茴香则采用了相对均衡分配策略。
3讨论与结论
水分短缺和养分贫瘠是沙化地生境的基本特征[18],植物必须经受养分匮乏和干旱胁迫的双重考验,才能在沙化地中存活。5种沙生植物生物学特征和生态学特点差异较大,沙生苔草具有较强的分蘖再生能力,赖草有发达的根茎,紫花列当为寄生型植物,藏茴香具有肉质根,地八角根系发达且固氮能力强,它们在沙地环境中表现出了与其生物学特征相对应的生态适应性。通过对比5种植物的表观性状,研究其在沙化环境下形态适应策略表明,5种植物的构件性状均具有极显著差异(P<0.01)。沙生苔草和赖草均具有极强的克隆生长潜力,能不断分生更新地下根茎系统,充分利用风蚀形成的裸沙斑块作为根茎扩展的空间资源[19],通过形态可塑性和繁殖对策,以克隆生长策略去适应或逃离胁迫环境[20]。
沙生苔草为游击型(Guerilla)克隆植物,因分枝强度大兼具密集型(Phalanx)生长特点[19],根茎上形成发达须根,极大增加了觅养水分的能力。赖草是典型的游击型克隆植物,通过强大的根茎和匍匐茎拓展能力,地上、地下协同作用寻觅水分或逃离胁迫生境。紫花列当作为寄生植物,受沙生旱生寄主植物的限制,通过退化叶减少水分散失,膨大茎贮藏水分的节水策略适应环境,且通过增长花穗轴,增加授粉、结实和种子散布的能力,利于后代逃离胁迫。藏茴香通过膨大的肉质根贮藏水分,通过高挺花穗增加种子散布范围。地八角采用相对保守的生存策略,通过根长和根幅增加获取水分的能力。5种植物采用不同的适应策略,增加获取潜在资源或利用有限资源的能力,得以在高寒沙化环境中长期留存和不断繁衍。
植物资源分配格局反映了植物发育对逆境的响应规律和适应对策[21],将更多资源投资于地下部分是对逆境的一种适应[22]。沙生苔草对地下构件投资大于地上部分,根茎含水量和生物量配比显著高于其它器官,将水分和养分蓄积在根茎或基茎,为促进分枝和增大冠幅奠定了物质基础。赖草水分和生物量优先向地上构件分配,小根冠比有利于逃离胁迫生境,这与任安芝等[20]的研究一致。紫花列当将水分更多向根茎和茎叶分配,将生物量优先向生殖分配,表明紫花列当在营养生长与生殖生长间采用了非平衡的分配策略。藏茴香采用了相对平衡的水分、生物量分配,营养生长与生殖生长协同适应了沙化生境。地八角生物量优先向根部分配,减少对其它构件的投入,是植物应对干旱胁迫普遍存在的适应策略[23]。由此可知,紫花列当在平衡生长和生殖、水分分配和生物量配比方面表现出较强的能力,具有极强抗旱和适应沙化环境的潜力。
本研究表明,在长期适应过程中,5种沙生植物形成了不同表观性状、水分分配和生物量分配的生态适应策略,并以此获取、权衡和分配环境资源,用于应对恶劣环境来保证植株的存活和繁衍。适应策略的差异决定了5种植物的生态适应能力的强弱,综合5种植物表观性状、水分和生物量及其分配差异分析表明,其生态适应能力的大小顺序为紫花列当>地八角>沙生苔草>赖草>藏茴香。紫花列当适应能力虽然最强,但作为寄生植物,必须依靠寄主提供营养和水分,从而受到寄主适应能力的限制;地八角根系发达,根瘤菌多,能够提高对水分的吸收并改善土壤的含氮量;沙生苔草和赖草的适应能力虽然不如地八角,但因其无性系克隆植物的特性具有较强的生长潜力;藏茴香适应能力相对最差,可能会在适应沙地的过程中逐渐被淘汰。在实际工作中,可依据5种植物的生态适应策略,以高山柳等灌木治理沙化草地的同时,结合5种本土草本植物,采用灌草结合的立体治沙模式,以期达到较好的防沙治沙效果,促进沙化地植被的恢复与重建。
参考文献References:
[1]骆建国,郑文靖.川西北草地沙漠化现状与防治对策研究,四川林业科技,2006,27(1):63-66.
Luo J G,Zhen W J.Research on the present situation of desertification of grassland in northwest Sichuan and its control measures.Journal of Sichuan Forestry Science and Technology,2006,27(1):63-66.(in Chinese)
[2]何文兴,李洪梅,陈放.川西北草原沙化区多年禾草混播草地的群落学特征.生态学杂志,2008,27(12):2079-2082.
He W X,Li H M,Chen F.Community characteristics of grassland sowed with mixed prennial gramineous species on desertified areas of northwest Sicuan.Chinese Journal of Ecolog,2008,27(12):2079-2082.(in Chinese)
[3]蔡应君,蒙嘉文.高寒草地生态环境现状分析及治理对策探讨.阿坝科技,2009(2):43-45.
[4]邓东周,鄢武先,武碧先,张炜,黄茜,余凌帆.川西北防沙治沙试点示范工程成果巩固必要性分析.四川林业科技,2015,36(1):69-72.
Deng D Z,Yan W X,Wu B X,Zhang W,Huang Q,Yu L F.The necessary analysis of strengthening results of demonstration project of preventing and controlling desertification pilot in northwest Sichuan.Journal of Sichuan Forestry Science and Technology,2015,36(1):69-72.(in Chinese)
[5]叶煜辉,姜明峰,陈艳,杨满业.赖草属植物的抗逆性研究进展与应用前景.生物学杂志,2009,26(4):54-57.
Ye Y H,Jiang M F,Chen Y,Yang M Y.Research progress of stress resistance mechanisms and application prospect ofLeymusHochst.Journal of Biology,2009,26(4):54-57.(in Chinese)
[6]王天慧.植物表型可塑性及生活史对策研究.长春:东北师范大学博士学位论文,2006.
Wang T H.The study of plant phenotypic plasticity and life-history strategies.PhD Thesis.Changchun:Northeast Normal University,2006.(in Chinese)
[7]刘海江,郭柯.浑善达克沙地丘间低地植物群落的分类与排序.生态学报,2003,23(10):2163-2169.
Liu H J,Guo K.Classification and ordination analysis of plant communities in Inter-dune lowland in Hunshandak sandy land. Acta Ecologica Sinica,2003,23(10):2163-2169.(in Chinese)
[8]王钰,宝音陶格涛.荒漠草原沙生针茅群落组成及其地上生物量研究.中国草地学报,2014,36(3):108-111.
Wang Y,Baoyintaogetao.Species composition and the aboveground biomass ofStipaglareosacommunity in desert steppe.Chinese Journal of Grassland,2014,36(3):108-111.(in Chinese)
[9]许冬梅,王堃,龙澍普.宁夏中部干旱带沙质草地赖草和中亚白草的抗逆生理特性.中国草地学报,2007,29(2):31-34.
Xu D M,Wang K,Long S P.Physiological characteristics of drought-resistant forLeymussecalinusandPennisetumcentrasiaticumin sandy grassland of arid region in central Ningxia.Chinese Journal of Grassland,2007,29(2):31-34.(in Chinese)
[10]付涛,蒋志荣,鲍婧婷,朱瑞清,鲁玉超.沙坡头地区3种沙生植物水分及光能利用效率特征比较分析.甘肃农业大学学报,2015,50(2):106-110.
Fu T,Jiang Z R,Bao J T,Zhu R Q,Lu Y C.Water and light use efficiency of 3 psammophytes in Shapotou area.Journal of Gansu Agricultural University,2015,50(2):106-110.(in Chinese)
[11]左力翔,陈佳村,李秧秧.陕北沙地不同树龄小叶杨的水分利用策略.中国水土保持科学,2014,12(6):59-67.
Zuo L X,Chen J C,Li Y Y.Water use strategies of different ages ofPopulussimoniion sandy land of northern Shaanxi.Science of Soil and Water Conservation,2014,12(6):59-67.(in Chinese)
[12]安钰,安慧.宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应.西北植物学报,2015,35(2):0373-0378.
An Y,An H.Response of plant growth and biomass allocation to grazing disturbance in desert steppe of Ningxia.Acta Botanica Boreali-Occidentalia Sinica,2015,35(2):0373-0378.(in Chinese)
[13]周瑞莲,侯玉平,左进城,王艳芳.不同沙地共有种沙生植物对环境的生理适应机理.生态学报,2015,35(2):340-349.
Zhou R L,Hou Y P,Zuo J C,Wang Y F.The physiological adaptation mechanisms of four common desert species in response to desert environments.Acta Ecologica Sinica,2015,35(2):340-349.(in Chinese)
[14]王兰州.植物群落趋中生态适应对策的机理.西北师范大学学报:自然科学版,1996,32(2):46-51.
Wang L Z.On mechanism of a mediotaxis strategy of ecological adaptability about plant community(MSEP).Journal of Northwest Normal University:Natural Science Edition,1996,32(2):46-51.(in Chinese)
[15]梁金华.内蒙古荒漠化草原四种植物的根系构型特点及其生态适应性研究.呼和浩特:内蒙古农业大学硕士论文,2012.
Liang J H.Study on the root architecture of four plants and their ecological adaptability in desert steppe of Inner Mongolia.Master Thesis.Hohhot:Inner Mongolia Agricultural University,2012.(in Chinese)
[16]肖冰雪,陈琴,郑群英,杨满业,孙海燕.川西北草原沙化治理现状、存在的问题及建议.草业与畜牧,2012(12):21-23.
Xiao B X,Chen Q,Zheng Q Y,Yang M Y,Sun H Y.The situation,problems and suggestions of controlling grassland desertification in northwestern Sichuan.Prataculture & Animal Husbandry,2012(12):21-23.(in Chinese)
[17]林波,刘庆,游翔,庞学勇.川西地区退耕还林工程及其对农村经济发展的影响.山地学报,2002,20(4):438-444.
Lin B,Liu Q,You X,Pang X Y.Engineering of changing cultivation into tree planting and its effects to the development of rural economics in western Sichuan,China.Journal of Mountain Science,2002,20(4):438-444.(in Chinese)
[18]朱灵益,宝音.毛乌素沙地乔灌木立地质量评价.北京:中国林业出版社,1993.
Zhu L Y,Baoyin.The Quality Evaluation of Sites of Shrubs in Mu Us Sandy Land.Beijing:China Forestry Press,1993.(in Chinese)
[19]何文兴,杨志荣,曹毅,陈放.川西北高寒沙区切断根茎对赖草和沙生苔草克隆生长的影响.生态学杂志,2005,24(6):607-612.
He W X,Yang Z R,Cao Y,Chen F.Effects of severied rhizome on clonal growth ofLeymussecalinusandCarexpraeclaraof alpine desertification grassland in Northwestern Sichuan.Chinese Journal of Ecology,2005,24(6):607-612.(in Chinese)
[20]任安芝,高玉葆,梁宇,陈世苹,刘爽,刘宁.白草和赖草无性系生长对干旱胁迫的反应.中国沙漠,1999,19(1):30-34.
Ren Z A,Gao Y B,Liao Y,Chen S P,Liu S,Liu N.Effect of drought stress on clonal growth ofPennisetumcentrasiaticumandLeymussecalinus.Journal of Desert Research,1999,19(1):30-34.(in Chinese)
[21]Enquist B J,Niklas K J.Global allocation rules for patterns of biomass portioning in seed plant.Science,2002,29(5):1517-1520.
[22]刘庆,钟章成.斑苦竹无性系生长与水分供应及其适应对策的研究.植物生态学报,1996,20(3):245-254.
Liu Q,Zhong Z C.The effects of water resources supply on clonal growth inPleioblsatusmaculatapopulation.Acata Phytoecologica Sinica,1996,20(3):245-254.(in Chinese)
[23]刘金平.坡度对野生荩草分株特征及生殖分配的影响.草业科学,2013,30(10):1602-1607.
Liu J P.Effects of sectional slope on reproductive ramets quantitative characteristics and reproductive investment of wildArthraxonhispidus.Pratacultural Science,2013,30(10):1602-1607.(in Chinese)
(责任编辑王芳)
Differences of ecological adaptation strategies of 5 herbs from the desertified grassland in the northwest Sichuan
Li Ying1, Zeng Xiao-lin1, You Ming-hong2, Liu Jin-ping1, Cai Jian1
(1.College of Life Sciences, China West Normal University, Nanchong 637009, China;2.Sichuan Academy of Grassland Science, Chengdu 611743, China)
Abstract:In the study, five species of natural herbs in the desertified grassland of northwest Sichuan, including Carex praeclara, Leymus secalinus, Orobanche coerulescens, Carum carvi and Astragalus bhotanensis, were selected to determine their adaptation strategies and mechanisms to desertification habitat by measuring apparent traits, water content, biomass and its allocation ratio, and analyzing the response of growth and development and resource allocation of different herbs to desertified habitats. The results of this study showed that apparent traits were extremely significant difference among five species of herbs(P<0.01), in which O. coerulescens and C. carvi adapted to desertification habitats by encouraging the sexual reproduction, however C. praeclara, L. secalinus and A. bhotanensis adapted to desertification habitats through crowns, rhizomes, and deep roots, respectively. Water content and its allocation proportion also were extremely significant difference among five species of herbs(P<0.01), in which the water content of C. praeclara, L. secalinus and A. bhotanensis preferentially assigned to reproductive branches, but O. coerulescens had priority to stems and leaves, and C. carvi used the water allocation with relative balance. Biomass and its allocation proportion showed aextremely significant difference among five species(P<0.01), indicating that the biomass of C. praeclara, L. secalinus, O. coerulescens and A. bhotanensis preferentially allocated to rhizomes, stems and leave, reproductive branches, and roots, respectively; however, C. carvi allocated the biomass to each component with relative balance. These suggested that five species of psammophytesshowed a difference strategies to trade off resource allocation and they obtained or allocated the limited available resources to increase the adaptability to habitat through varieties of phenotypic plasticity, water using ability and biomass allocation, etc.
Key words:desertified grassland; psammophytes; apparent traits; water allocation; adaptation strategy
DOI:10.11829/j.issn.1001-0629.2015-0506
*收稿日期:2015-09-08
基金项目:四川省科技支撑计划(2011NZ0064)
通信作者:刘金平(1972-),男,山西临县人,副教授,博士,主要从事草地生态学研究。E-mail:jpgg2000@163.com
中图分类号:S812.29
文献标志码:A
文章编号:1001-0629(2016)5-0843-08*1
Corresponding author:Liu Jin-pingE-mail:jpgg2000@163.com
李莹,曾晓琳,游明鸿,刘金平,蔡捡.5种川西北沙化地草本植物生态适应策略的差异性.草业科学,2016,33(5):843-850.
Li Y,Zeng X L,You M H,Liu J P,Cai J.Differences of ecological adaptation strategies of 5 herbs from the desertified grassland in the northwest Sichuan.Pratacultural Science,2016,33(5):843-850.
接受日期:2015-11-03
第一作者:李莹(1991-),女,四川眉山人,在读硕士生,主要从事草地生态学研究。E-mail:568824801@qq.com
