西藏色季拉山高山松种群点格局分析
2016-06-06沈志强方江平
沈志强,卢 杰,华 敏,方江平
(1 西藏大学 农牧学院,西藏 林芝 860000;2 武汉大学 资源与环境科学学院,湖北 武汉 430079)
西藏色季拉山高山松种群点格局分析
沈志强1,卢杰1,华敏2,方江平1
(1 西藏大学 农牧学院,西藏 林芝 860000;2 武汉大学 资源与环境科学学院,湖北 武汉 430079)
[摘要]【目的】 研究高山松种群不同生长阶段的空间分布格局类型及其空间关联性。【方法】 2014年6-8月,在西藏色季拉山选择有代表性的区域设置50 m×50 m的高山松林样地4块,对样地内的高山松进行定位和每木检尺,按胸径<12 cm、≥12~<32 cm、≥32 cm将高山松分为小树、中树和大树,运用点格局分析方法中的Ripley’s L函数,对高山松种群不同发育阶段的空间格局及关联性进行分析。【结果】 (1)色季拉山高山松种群径级结构呈典型的“金字塔”型,种群自然更新良好,属增长型种群;(2)高山松不同发育阶段的分布格局各异,小树在小尺度范围内的聚集强度均明显强于中树和大树,中树主要表现为随机分布-集群分布-随机分布,而大树主要表现为随机分布;(3)样地1与样地2中,高山松小树与中树、大树之间的空间关联性基本上均呈负关联关系,但中树与大树之间基本上无关联;样地3和样地4中,高山松不同龄级的小树、中树和大树三者之间相互独立,关联性小。【结论】 在从小树发育到大树的过程中,高山松种群的空间分布格局演变过程基本表现为由集群分布到随机或均匀分布;高山松种群的更新不仅受种间竞争的影响,同样受种内竞争的制约;在环境条件相似的情况下,高山松种群自身的生物、生态学特性是影响其分布格局的最主要因素。
[关键词]高山松;空间分布;点格局分析;色季拉山
空间格局是植物种群的重要特征,是种群自身特性、种间关系及环境条件综合作用的结果[1]。对植物种群空间格局的研究一直是当代生态学研究的热点,对于认识植物种群的种内和种间关系、生态过程以及植物种群与生境的相互关系具有重要作用[2]。点格局分析方法首先由北京师范大学张金屯教授引入国内,推动了植物种群空间分布的研究[3-4]。点格局分析法能够最大限度地利用点与点之间的距离,并提供较为全面的空间尺度信息[5]。因此,它可以分析任意尺度上种群的空间格局和种间关系,同时还能够给出空间格局的最大聚集强度及其对应的尺度,为群落内种群空间格局的比较提供了方便。
高山松(Pinusdensata)为松科松属植物,是中国特有植物种,广泛分布于川滇和青藏高原地区,其属于强阳性树种,在西藏地区常生长在海拔2 600~3 500 m的山坡、河谷和山谷等的阳坡地带,是西藏色季拉山地区主要的植物种群之一[6],在保护生物多样性、涵养水源和水土保持等方面具有极为重要的生态价值。目前对该物种的研究主要集中在种群分布[7]、群落特性[8]、幼苗生长[9-10]以及天然更新[11-12]等方面,而对其种群格局的研究依然不够充分。为此,本研究采用点格局分析法对西藏色季拉山地区高山松种群进行调查,分析了该种群不同年龄结构的空间分布格局及关系,以期为当地合理利用生物资源和充分发挥其生态价值奠定理论基础。
1材料与方法
1.1研究区概况
色季拉山位于西藏东南部,是念青唐古拉山向南延伸的余脉,山脉走向由西北向东南延伸,主峰海拔5 200 m左右。植被从高海拔到低海拔主要分布有草甸、草地、高山寒带灌丛、亚高山寒温带暗针叶林、松林、落叶阔叶林和山地温带针阔混交林。研究地区主要位于29°45′22″~29°46′23″N,94°13′58″~94°15′54″E,海拔3 113~3 610 m,平均坡度28°。本区域气候属于湿润和半湿润的亚高山寒温带气候,春夏多雨,秋冬干燥少雨,大气相对湿度64%,年降水量800 mm,年平均气温-1.8 ℃,1月平均气温-7.8 ℃,7月平均气温9.8 ℃。土壤以山地棕壤和酸性棕壤为主[13]。研究地区乔木层树种主要有高山松、川滇高山栎(Quercusaquifolioides)、急尖长苞冷杉(Abiesgeorgeivar.smithii)和林芝云杉(Picealikiangensisvar.linzhiensis),灌木层主要有西南花楸(Sorbusrehderiana)、雅致山马蝗(Desmodiumelegans)、野丁香(Leptodermispotaninii)、林芝杜鹃(Rhododendronnyingchiense)等,草本层主要有桃儿七(Sinopodophyllumhexandrum)、五裂蟹甲草(Parasenecioquinquelobus)、宽叶兔儿风(Ainsliaealatifolia)、白花酢浆草(Oxalisacetosella)、高山露珠草(Circaeaalpina)等,另有藤本植物西藏铁线莲(Clematistenuifolia)等。
1.2样地调查
2014年6-8月,在实地踏查的基础上对西藏色季拉山地区的高山松林进行实地调查,在海拔 3 113~3 610 m选择能代表区域种群特性的典型样地进行调查,共设置4个50 m×50 m的固定样地,记录每个样地的海拔、坡向、坡度和经纬度等。然后对样地内的高山松种群进行定位和每木检尺,记录树高、胸径、冠幅等指标,并分别统计样地内胸径<12 cm(小树)、≥12~<32 cm(中树)、≥32 cm(大树)的高山松的株数。样地基本概况见表1。
1.3数据分析
1.3.1种群结构分析有研究表明,在相同环境条件下,同一树种龄级和胸径级对环境的反应规律具有一致性[14-16],因此以胸径(DBH)作为表征其年龄的指标来反映高山松的不同生长阶段。根据高山松的生活史特征及实地调查数据的特点,将胸径在12 cm以下的高山松划定为Ⅰ径级,然后以10 cm为步长增加一级,将色季拉山高山松种群共划分为13个径级(Ⅰ-ⅩⅢ),统计各个样地内各径级高山松的株数。依据径级特点,将色季拉山高山松种群分为小树(Ⅰ级)、中树(Ⅱ-Ⅲ级)和大树(Ⅳ-ⅩⅢ)3个年龄段。采用点格局分析方法,分析不同发育阶段个体的空间格局及关系,进而准确表达高山松的空间分布格局,对高山松种群格局关系进行详细分析与判定。
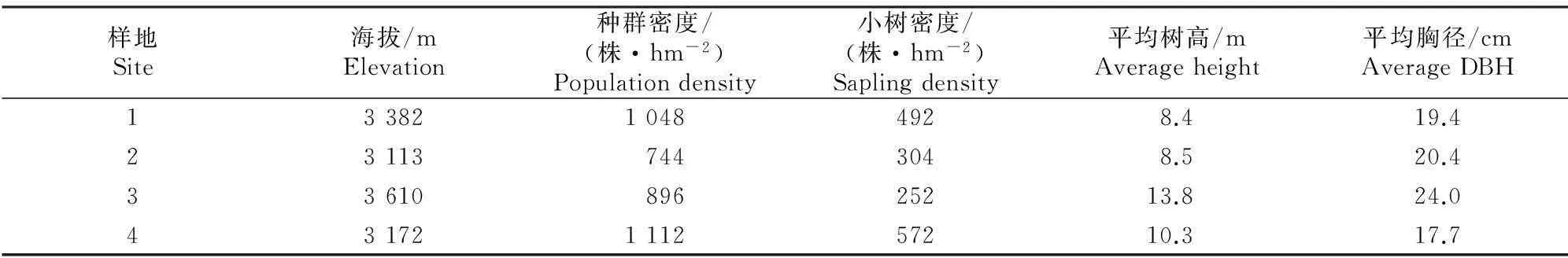
表 1 西藏色季拉山高山松林样地基本概况
注:小树指胸径<12 cm的高山松。
Note:Saplings refer to thePinusdensatawith DBH of <12 cm.
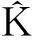
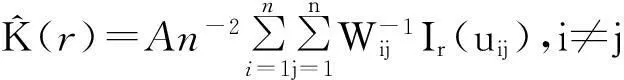
(1)
式中:A表示样地面积;n为样地中某一植物种群的个体总数;uij表示点i到j之间的距离;r表示空间尺度,当uij≤r时,Ir=1,否则Ir=0;Wij是以i为圆心、以uij为半径的圆落在面积A中的弧长与整个圆周的比值,其可以校正边界效应引起的误差。

(2)
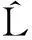
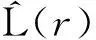
1.3.3不同龄级空间关联分析对2个种群之间关系的分析,其实质就是对两者的点格局分析,或者叫做多元点格局分析。其计算公式如下:
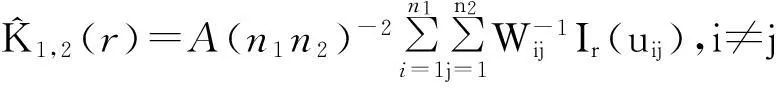
(3)
式中:n1、n2分别表示2个物种的个体数量,i、j则代表着种1和种2中的个体,uij、Ir、Wij、A的含义与公式(1)一致。现将公式(3)同样进行简化,得到如下公式:

(4)
2结果与分析
2.1高山松种群的径级结构
种群径级结构能够反映种群的生存状态[19]。由图1可知,色季拉山高山松种群径级结构呈典型的“金字塔”型,表现为增长型种群。样地1、2、4的各径级中Ⅰ级小树数量最多,分别为492,304和572株/hm2,样地3中Ⅲ级株数最多,为316株/hm2,并且在各样地中中树株数均明显多于大树,这说明种群的自然更新较好。然而由于自然环境和人为因素的干扰,不同样地间的种群径级结构也存在差异,样地1、4中,种群个体主要集中在Ⅰ级,分别占总个体数的46.9%和51.4%,为增长型;样地3中,Ⅱ、Ⅲ级(中树)株数最多,共384株/hm2,占总数的42.9%。样地1最大径级为Ⅹ、样地2为Ⅶ、样地3为ⅩⅢ、样地4为Ⅵ,并且样地1第Ⅷ级、样地3第Ⅶ-Ⅻ级处出现“断层”现象,这在某种意义上与种群“潮动”发展现象类似[20]。
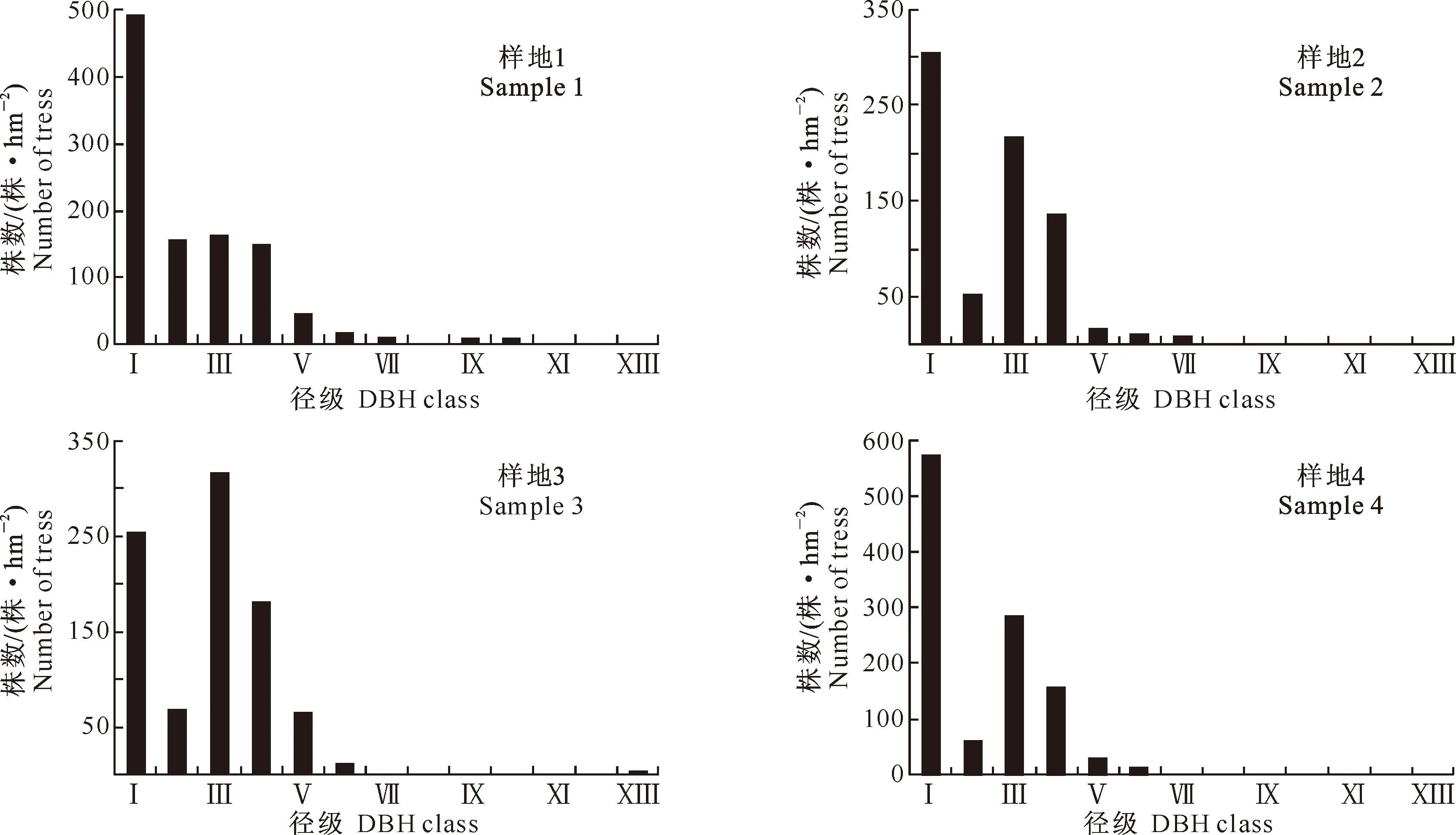
图 1 西藏色季拉山高山松种群的径级结构
2.2高山松种群的空间分布
西藏色季拉山高山松种群在4个50 m×50 m固定标准样地中的个体分布见图2。

图 2西藏色季拉山高山松个体的空间分布点图
Fig.2Space distribution ofPinusdensataindividuals in Sejila Mountains,Tibet
从图2可以看出,样地4中高山松的个体数最多,密度最大,为 1 112 株/hm2;而样地2中高山松的密度远远低于样地4,其密度为744株/hm2;4个样地内高山松个体密度大小顺序为样地4>样地1>样地3>样地2。
2.3高山松不同龄级种群的空间格局
不同龄级高山松种群在4个标准样地中的空间分布格局分析结果见图3。
图 3西藏色季拉山4块样地中高山松各发育阶段的空间分布格局
Fig.3Patterns ofPinusdensataat diffferent developmental stages in 4 plots of Sejila Mountains,Tibet
由此可见,4个样地中,小树在小尺度范围内的聚集强度均明显强于中树和大树,这可能是由于小树需要的资源较少、竞争相对较弱以及种子散布的不均匀性和萌发率的差异性引起的,因而呈现集群分布,这有利于个体的生长发展。中树在空间格局上表现为随机分布-集群分布-随机分布,而大树在4个样地中基本上均呈随机分布。也可以得出高山松种群在从小树、中树发育到大树的过程中,其空间格局表现基本上是由集群分布到随机或均匀分布的演变过程,这可能与高山松自身的生长特性有关。
2.4高山松不同龄级种群的空间关联性
对4块样地中高山松种群不同龄级间空间关联性的分析结果见图4。

图 4西藏色季拉山4块样地高山松各发育阶段间的空间关联性
Fig.4Spatial association ofPinusdensataat different developmental stages in 4 plots in Sejila Mountains,Tibet
样地1中,1~25 m尺度下,高山松小树和中树以及小树和大树基本上均呈显著负关联,小树和中树在0~1 m尺度下无关联;中树和大树在2~4 m尺度下呈负关联,其他尺度下无关联。样地2中,0~13 m尺度下,小树和中树呈明显负关联,在13~25 m尺度下无关联;小树和大树在15~18 m尺度下无关联,其他尺度下均呈显著负关联;中树和大树在0~25 m尺度下均无关联。样地3中,在0~25 m尺度下,小树、中树和大树间均无关联。样地4中,0~25 m尺度下,小树和中树以及小树和大树均无关联,而中树和大树在0~3 m尺度下呈正关联,其他尺度下无关联。
由此可以看出,在样地1和样地2中,高山松小树和中树、大树之间的空间关联性基本上呈负关联关系,但中树和大树之间基本上呈无关联关系,这说明小树和中树、大树间竞争激烈,不利于小树的成长,而中树和大树间则相对独立。样地3和样地4中,高山松不同龄级的小树、中树和大树三者之间基本上无关联关系,说明样地3和样地4中的高山松不同龄级间相互各自独立,这有利于高山松种群的稳定增长。
3讨论
西藏色季拉山高山松种群径级结构呈典型的“金字塔”型,幼苗补给相对充足,种群年龄属增长型,但在不同样地间种群的径级结构和个体数量存在较明显的差异。这可能与高山松种群分布的生境类型有关,即与海拔等自然因素有关,也可能与高山松自身的生物学特性有关[21]。随着年龄的增长,高山松个体数量逐渐减少,这表明其更新良好,为增长型种群。
影响植物种群空间分布格局的因素主要是其自身的生物学特性、环境及其相互间的作用[21-23],对空间分布格局的研究能够揭示种群的动态变化规律和未来的发展趋势[23]。在本研究中,高山松种群在从小树、中树发育到大树的过程中,其空间格局表现基本上是由集群分布到随机或均匀分布的演变过程,小树在小尺度上呈强烈的集群分布,而中树和大树基本上呈随机或均匀分布。这一研究结果与国内学者的一些研究结果[24-26]相一致。造成这种现象的原因可能有以下几点:(1)种子传播对早期种群空间分布格局具有重要的影响,同时种子萌发的不均匀性也可能会导致这种现象;(2)小树在小尺度上呈集群分布,因为早期种子会散落在母株周围,其能够相互庇护并以集群形式利用有利资源促进自身生长,提高其存活几率,更利于小树的生长[27];(3)随着种群的生长发育,不同龄级树木对光因子和营养元素的竞争日益激烈,种群内部自疏作用增强,而小树、中树在竞争中处于劣势,导致死亡,种群最终趋于均匀或随机分布[28]。
种内联结是对一定时期内植物种群组成个体之间相互关系的静态描述,表示同一物种不同龄级在空间分布上的相互关联性,其能够反映出种群的现状和种群内的相互关系[29]。通过对色季拉山高山松种群不同龄级间的空间关联性分析可以得出,在样地1和样地2中,小树与中树、大树之间呈明显的负关联,说明高山松中树、大树对小树的更新有强烈的抑制作用,高山松种群不同龄级间竞争激烈,小树在生长过程中与中树、大树争夺资源。而中树与大树之间基本上无关联,这表明中树和大树相互独立,也可能是保持有一定的距离,减轻了它们对资源的争夺,有利于种群的健康发展。样地3和样地4中,高山松小树、中树和大树三者之间基本均无明显的关联性,其中的原因可能是由于个体具有较大的生长空间,主要是因为高山松属于强阳性树种,小树的生长需要一定的光照条件,小树、中树在大树下难以获得充足的光照,在林冠下不能很好地存活和生长,其各自独立,降低了相互依赖性,这对不同龄级的高山松种群发展是有利的,对于整个种群的稳定增长具有重要意义[27]。
研究西藏色季拉山高山松种群结构和空间格局等,对正确认识和揭示高山松种群的动态变化、种群与环境之间的关系具有重要作用。在今后的工作中将进一步研究高山松种群与其他植物种群间的关系,以期进一步揭示植物种群间的维持和共存机制,为该地区合理利用生物资源奠定理论基础。
[参考文献]
[1]张金屯.数量生态学 [M].北京:科学出版社,2004.
Zhang J T.Quantitative ecology [M].Beijing:Science Press,2004.(in Chinese)
[2]杨洪晓,张金屯,吴波,等.毛乌素沙地油蒿种群点格局分析 [J].植物生态学报,2006,30(4):563-570.
Yang H X,Zhang J T,Wu B,et al.Point pattern analysis ofArtemisiaordosicapopulation of the Mu Us Sandy Land [J].Journal of Plant Ecology,2006,30(4):563-570.(in Chinese)
[3]张兴旺,张小平,郭传友,等.皖北石灰岩山地青檀种群不同发育阶段的点格局分析 [J].生态学杂志,2013,32(3):542-550.
Zhang X W,Zhang X P,Guo C Y,et al.Point pattern analysis ofPteroceltistatarinowiipopulation at its different development stages in limestone mountain area of north Anhui,East China [J].Chinese Journal of Ecology,2013,32(3):542-550.(in Chinese)
[4]杨晓凤,苗艳明,张钦弟,等.五鹿山白皮松林不同龄级立木的点格局分析 [J].植物研究,2013,33(1):24-30.
Yang X F,Miao Y M,Zhang Q D,et al.Spatial pattern analysis of individuals in different age-classes ofPinusbungeanainWulu Mountain Reserve,Shanxi,China [J].Bulletin of Botanical Research,2013,33(1):24-30.(in Chinese)
[5]王本洋,余世孝.种群分布格局的多尺度分析 [J].植物生态学报,2005,29(2):235-241.
Wang B Y,Yu S X.Multi-scale analyses of population distribution patterns [J].Acta Phytoecologica Sinica,2005,29(2):235- 241.(in Chinese)
[6]李文华.西藏森林 [M].北京:科学出版社,1985.
Li W H.Tibet forest [M].Beijing:Science Press,1985.(in Chinese)
[7]禄树晖,潘朝晖.藏东南高山松种群分布格局 [J].东北林业大学学报,2008,36(11):22-24.
Lu S H,Pan C H.Distribution pattern ofPinusdensatapopulation in the Southeast Tibet [J].Journal of Northeast Forestry University,2008,36(11):22-24.(in Chinese)
[8]卢杰,郭其强,郑维列,等.藏东南高山松种群结构及动态特征 [J].林业科学,2013,49(8):154-160.
Lu J,Guo Q Q,Zheng W L,et al.Population structure and dynamic characteristics ofPinusdensatain Southeast Tibet [J].Scientia Silvae Sinicae,2013,49(8):154-160.(in Chinese)
[9]何飞飞,李永梅,李翠萍,等.滇西北生态公益林区高山松容器育苗基质配方研究 [J].湖南农业大学学报(自然科学版),2008,34(6):631-634.
He F F,Li Y M,Li C P,et al.Study onPinusdensataseedling substrates of ecological forest in westnorth Yunnan [J].Journal of Hunan Agricultural University(Nat Sic Ed), 2008,34(6):631-634.(in Chinese)
[10]蔡年辉,许玉兰,白青松,等.不同种群高山松1年生播种苗木生长节律及其变异 [J].东北林业大学学报,2013,41(5):11-15.
Cai N H,Xu Y L,Bai Q S,et al.Annual seedling growth dynamic rhythm and variation of one-yearPinusdensataamong populations [J].Journal of Northeast Forestry University,2013,41(5):11-15.( in Chinese)
[11]杨冬.高山松天然更新特性 [J].林业科技,2005,30(6):15-17.
Yang D.The natural regeneration characteristics ofPinusdensata[J].Forestry Science and Technology,2005,30(6):15-17.(in Chinese)
[12]吴明山,许彦红,王俊峰,等.因子分析在林地更新评价中的应用:以香格里拉高山松为例 [J].西南林业大学学报,2011,31(6):36-44.
Wu M S,Xu Y H,Wang J F,et al.Application of factor analysis to evaluation of forest regeneration:a case study ofPinunsdensataforest in Shangri-la County [J].Journal of Southwest Forestry University,2011,31(6):36-44.(in Chinese)
[13]周晨霓,马和平.西藏色季拉山典型植被类型土壤活性有机碳分布特征 [J].土壤学报,2013,50(6):1246-1251.
Zhou C N,Ma H P.Distribution of labile organic carbon in soil as affected by vegetation typical of Sygera Mountains,Tibet,China [J].Acta Pedologica Sinica,2013,50(6):1246-1251.(in Chinese)
[14]龙成,杨小波,龙文兴,等.铜鼓岭热带常绿季雨矮林5种蒲桃属植物的种群结构及空间格局 [J].林业科学,2015,51(2):18-27.
Long C,Yang X B,Long W X,et al.Population structure and spatial pattern of fiveSyzygiumspecies in Tropical Evergreen Monsoom Elfin Forest,Tongguling [J].Scientia Silvae Sinicae,2015,51(2):18-27.(in Chinese)
[15]Frost I,Rydin H.Spatial pattern and size distribution of the animal-dispersed treeQuercusroburin two spruce-dominated forests [J].Ecoscience,2000,7(1):38-44.
[16]胡尔查,王晓江,张文军,等.乌拉山自然保护区白桦种群的年龄结构和点格局分析 [J].生态学报,2013,33(9):2867-2876.
Hu E C,Wang X J,Zhang W J,et al.Age structure and point pattern ofButulaplatyphyllain Wulashan Natural Reserve of Inner Mongolia [J].Acta Ecologica Sinica,2013,33(9):2867-2876.(in Chinese)
[17]Ripley B D.Modelling spatial patterns [J].Journal of the Royal Statistical Society:Series B,1977,39(2):172-212.
[18]常静,潘存德,师瑞锋.梭梭-白梭梭群落优势种种群分布格局及其种间关系分析 [J].新疆农业大学学报,2006,29(2):26-29.
Chang J,Pan C D,Shi R F.Analysis on dominant species distribution patterns and relation ofAss.Haloxylonpersicum+H.ammodendron[J].Journal of Xinjiang Agricultural University,2006,29(2):26-29.(in Chinese)
[19]Lan G Y,Getzin S,Wiegand T,et al.Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China [J].PLoS One,2010,7(11):4531-4575.
[20]阳含熙,伍业钢.长白山自然保护区阔叶红松林林木种属组成、年龄结构和更新策略的研究 [J].林业科学,1988,24(1):19-27.
Yang H X,Wu Y G.Tree composition,age structure and regeneration strategy of the mixed broadleafKoreanpineForest in Changbaishan Biosphere Reserve,China [J].Scientia Silvae Sinicae,1988,24(1):19-27.(in Chinese)
[21]Condit R,Ashton P S,Baker P,et al.Spatial patterns in the distribution of tropical species [J].Science,2000,288(5470):1414-1418.
[22]Crawley M J.Plant ecology [M].London:Blackwell Scientific Publications,1986:97-185.
[23]Wu X P,Zheng Y,Ma K P.Population distribution and dynamics ofQuercusliaotungensis,FraxinusrhynchophyllaandAcermonoin Dongling Mountain, Beijing [J].Acta Botanica Sinica,2002,44(2):212-223.
[24]黄明钗,史艳财,韦霄,等.珍稀濒危植物金花茶的点格局分析 [J].生态学杂志,2013,32(5):1127-1134.
Huang M C,Shi Y C,Wei X,et al.Point pattern analysis of rare and endangered plantCamellianitidissimaChi [J].Chinese Journal of Ecology,2013,32(5):1127-1134.(in Chinese)
[25]张健,郝占庆,宋波,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性 [J].应用生态学报,2007,18(8):1681-1687.
Zhang J,Hao Z Q,Song B,et al.Spatial distribution patterns and associations ofPinuskoraiensisandTiliaamurensisin broad-leaved Korean pine mixed forest in Changbai Mountains [J].Chinese Journal of Applied Ecology,2007,18(8):1681-1687.(in Chinese)
[26]郭垚鑫,胡有宁,李刚,等.太白山红桦种群不同发育阶段的空间格局与关联性 [J].林业科学,2014,50(1):9-14.
Guo Y X,Hu Y N,Li G,et al.Spatial pattern and spatial association ofBetulaalbosinensisat different developmental stages at Taibai Mountain [J].Scientia Silvae Sinicae,2014,50(1):9-14.(in Chinese)
[27]张震,刘萍,丁易,等.天山云杉林不同发育阶段种群分布格局研究 [J].北京林业大学学报,2010,32(3):75-79.
Zhang Z,Liu P,Ding Y,et al.Distribution patterns ofPiceaschrenkianavar.tianshanicapopulations at different developmental stages in the western Tianshan Mountain,northwestern China [J].Journal of Beijing Forestry University,2010,32(3):75-79.(in Chinese)
[28]闫海冰,韩有志,杨秀清,等.华北山地典型天然次生林群落的树种空间分布格局及其关联性 [J].生态学报,2010,30(9):2311-2321.
Yan H B,Han Y Z,Yang X Q,et al.Spatial distribution patterns and associations of tree species in typical natural secondary mountain forest communities of Northern China [J].Acta Ecologica Sinica,2010,30(9):2311-2321.(in Chinese)
[29]王磊,孙启武,郝朝运,等.皖南山区南方红豆杉种群不同龄级立木的点格局分析 [J].应用生态学报,2010,21(2):272-278.
Wang L,Sun Q W,Hao C Y,et al.Point pattern analysis of different age-classTaxuchinensisvar.maireiindividuals in mountainous area of southern Anhui Province [J].Chinese Journal of Applied Ecology,2010,21(2):272-278.(in Chinese)
Spatial point pattern ofPinusdensataforests of Sejila Mountains in Tibet
SHEN Zhi-qiang1,LU Jie1,HUA Min2,FANG Jiang-ping1
(1AgriculturalandAnimalHusbandryCollege,TibetUniversity,Linzhi,Tibet860000,China;2SchoolofResourceandEnvironmentalSciences,WuhanUniversity,Wuhan,Hubei430079,China)
Abstract:【Objective】 The spatial distribution pattern and spatial correlation of Pinus densata at different developmental stages were studied.【Method】 During June to August,2014,four 50 m×50 m P.densata plots were set in a representative area of Sejila Mountains.Location and tally of P.densata were conducted and the P.densata trees were divided into saplings,middle trees and large trees according to DBH values of <12 cm,≥12-<32 cm,and ≥32 cm,respectively.The spatial pattern and spatial correlation of P.densata at different developmental stages were analyzed using Ripley’s L function in point pattern analysis.【Result】 (1) The demographic structure of P.densata population in Sejila Mountains followed a typical “pyramid” shape,suggesting a high regeneration rate and good growth condition and belonging to expanding population.(2) The distribution patterns of P.densata varied a lot at different developmental stages.The aggregation intensities of saplings at small scales were significantly stronger than those of middle trees and large trees.The spatial distribution of middle trees changed from random,aggregated,to random,while large trees showed random distribution.(3) In plots 1 and 2,the spatial correlations between saplings and middle trees and between large trees were negative,and there was no correlation between middle trees and large trees.In plots 3 and 4,saplings,middle trees,and large trees at different ages were independent with less correlations.【Conclusion】 During the growth process of P.densata population from saplings to large trees,the spatial pattern developed from aggregated to random or uniform distribution.The regeneration of P.densata populations depended on interspecific relationship and intraspecific relationship.The spatial pattern of P.densata population in this area with uniform environment conditions was controlled by biotic interactions and ecological characteristics.
Key words:Pinus densata;spatial distribution;point pattern analysis;Sejila Mountain
DOI:网络出版时间:2016-04-0709:0010.13207/j.cnki.jnwafu.2016.05.010
[收稿日期]2015-09-06
[基金项目]国家科技支撑计划重点项目(2013BAC04B01);西藏大学农牧学院2014年提升综合实力建设项目(生态学科)
[作者简介]沈志强(1989-),男,安徽宿州人,在读硕士,主要从事森林生态系统研究。E-mail:syg521hm@163.com[通信作者]卢杰(1973-),男,四川安岳人,副教授,主要从事高原植物生态与植物保护研究。E-mail:tibetlj@163.com
[中图分类号]S718.54
[文献标志码]A
[文章编号]1671-9387(2016)05-0073-09
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20160407.0900.020.html
