玉米抗粗缩病材料产量性状的配合力分析
2016-05-30卫晓轶魏锋丁微魏芳王向阳张学舜
卫晓轶 魏锋 丁微 魏芳 王向阳 张学舜
摘要:采用NCII遗传交配设计方法,以郑58、昌7-2、PH6WC、PH4CV为测验种,通过田间试验,分析了7份新引抗粗缩病材料的穗部性状和产量配合力。结果显示,由7份新引材料穗部性状和产量一般配合力(GCA)综合分析来看,Y7和Y4易组配出高产组合,Y6易组配出稳产组合;从产量特殊配合力(SCA)分析得知,产量SCA效应值较大的组合有Y4×PH4CV、Y7×PH6WC、Y6×PH4CV、Y7×郑58,其产量分别为11.3、11.4、11.2、11.1 t/hm2,均极显著地高于对照郑单958和先玉335。综合来看,Y4×PH4CV、Y7×PH6WC、Y6×PH4CV、Y7×郑58等组合表现较好。
关键词:玉米;产量;一般配合力;特殊配合力
中图分类号:S513.034文献标识号:A文章编号:1001-4942(2016)10-0020-04
玉米是我国种植面积最大的粮食作物,玉米的产量直接关系着国家的粮食安全[1]。然而,现在我国存在着玉米种质资源缺乏和遗传基础狭窄问题,这已成为玉米生产可持续发展的首要限制因素[2-4]。引进外来种质可以丰富现有种质的遗传多样性,可以拓宽玉米杂交种的遗传模式[5-7]。在作物杂交育种中,配合力是至关重要的[8-10]。配合力的高低是衡量一个玉米自交系优劣的重要条件之一[11-13]。杂交种选育的主要步骤,其一是自交系的选育,其二是选择适合的自交系亲本来组配优良的杂交种[14]。因此,在杂交种的选育过程中,选择高配合力的自交系作为亲本至关重要。
玉米粗缩病是由灰飞虱传播粗缩病毒(MRDV)引起的病毒病。近年来,该病在我国东北、华北、西北、华中及长江以南部分地区蔓延流行,并有逐年加重的趋势,是一种严重的玉米病害,给玉米生产带来了巨大的损失[15-17]。由于目前大面积种植的玉米品种多数表现感或高感玉米粗缩病,缺乏抗病品种,生产上主要采用化学农药来防治此病,不仅成本高、污染重,而且防治效果较差[18]。因此,在拓宽玉米种质基础的前提下,充分挖掘抗病基因,培育高抗粗缩病的玉米新品种是减轻产量损失最经济有效的措施[19]。
为了分析新引抗粗缩病材料与我国生产中主要应用种质之间的杂种优势潜力及遗传关系,本研究利用NCII遗传交配设计方法,以国内目前大面积推广的杂交种郑单958和先玉335双亲本(郑58、昌7-2、PH6WC、PH4CV)为测验种,对7份新引抗粗缩病材料进行配合力鉴定,以明确其产量性状的一般配合力效应(GCA)和特殊配合力效应(SCA),旨在从供试材料中筛选强优势组合和一般配合力较高的自交系,并对新引材料进行遗传分析、评价,以了解新引材料的性状表现及产量潜力。
1材料与方法
1.1试验材料
以7份新引自交系材料Y1、Y2、Y3、Y4、Y5、Y6、Y7为被测系,以国内骨干自交系郑58、PH6WC、昌7-2、PH4CV为测验种组配出28个组合。以郑单958和先玉335作对照种。
1.2试验设计
试验于2015年在新乡市农业科学院试验基地(辉县)进行。6月14日播种,10月11日收获。采用随机区组试验设计,郑单958(CK1)和先玉335(CK2)作对照。每个组合2行区,行长5 m,行距0.6 m,株距0.25 m,种植密度为6.75万株/hm2。重复3次。按常规大田措施进行田间管理。
1.3调查项目
于每个小区内选择有代表性的10个果穗进行考种,测量穗长、穗粗、穗行数、行粒数、百粒重,同时测定籽粒含水量。收获小区鲜穗,计算折合14%水分后的产量[20]。
配合力分析采用Griffing IV分析方法计算[21],方法如下:
一般配合力效应值:GCAgi=(nTi-2T)/n(n-2);
特殊配合力效应值:SCAsij=Yij -(Ti+Tj)/(n-2)+2T/(n-1)(n-2)。
式中,n为待测群体数,Ti、Tj分别为第i、j个群体的所有组合的总和,T为所有参试双列杂交组合之总和,Yij为第i和第j群体的组合值。
2结果与分析
2.1一般配合力效应分析
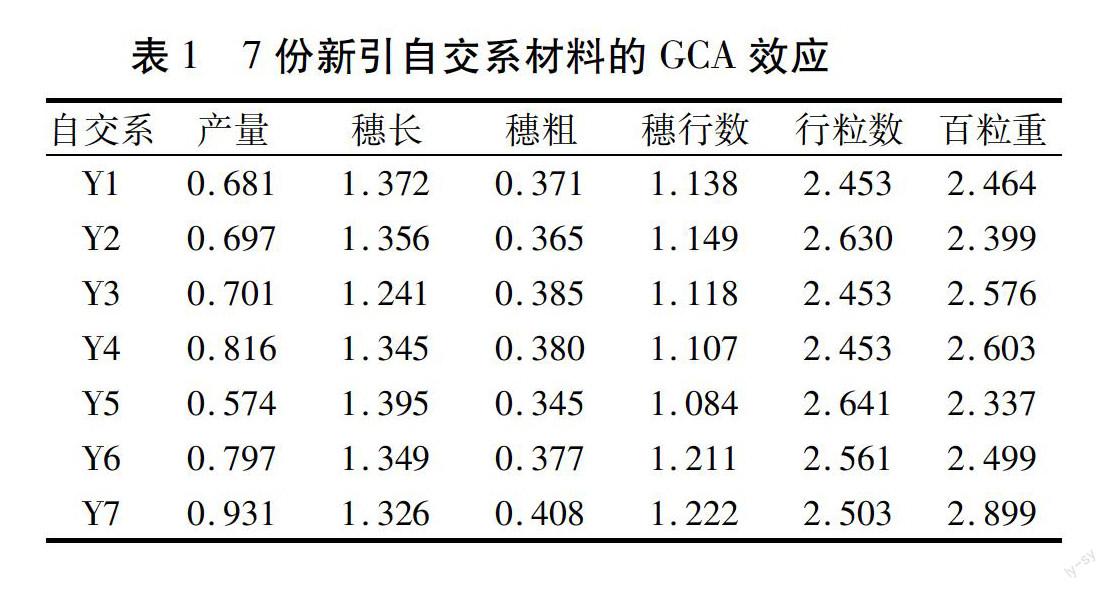
对7份新引材料一般配合力进行分析,结果见表1。从中可以看出,Y7产量的一般配合力效应值最高(0.931),Y4次之(0.816),Y5最低(0.574)。
穗长的一般配合力效应:Y5、Y1的GCA较高,分别为1.395和1.372;Y3最低(1.241)。
穗粗的一般配合力效应:Y7的GCA最高(0.408);Y3、Y4次之,分别为0.385和0.380;Y5最低(0.345)。
穗行数的一般配合力效应:Y7的GCA最高(1.222),Y6次之(1.211),Y5最低(1.084)。
行粒数的一般配合力效应:Y5的GCA最高(2.641),Y1、Y3、Y4的GCA较低。
百粒重的一般配合力效应:Y7的GCA最高(2.899),Y4次之(2.603),Y5最低(2.337)。
2.2产量特殊配合力效应分析
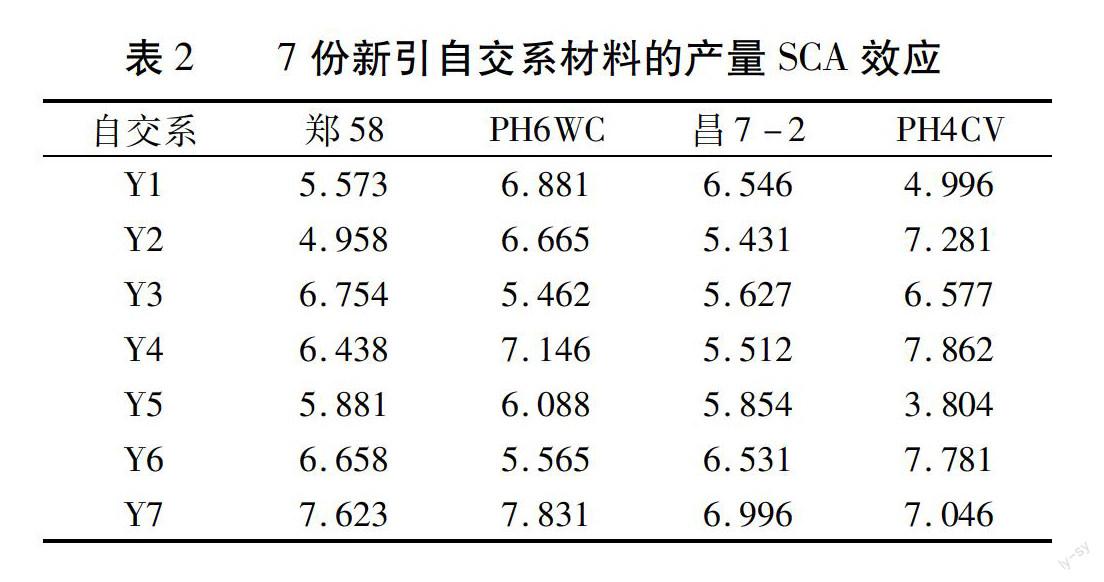
对28个组合产量特殊配合力效应结果(表2)进行分析可知,7份新引材料之间相比,Y7的特殊配合力最高,Y4次之,Y5最低。其中,Y7与郑58、PH6WC、昌7-2、PH4CV四个测验种组配组合的产量SCA效应值分别为7.623、7.831、6.996、7.046。Y4与郑58、PH6WC、昌7-2、PH4CV四个测验种组配组合的产量SCA效应值分别为6.438、7.146、5.512、7.862。Y5与郑58、PH6WC、昌7-2、PH4CV四个测验种组配组合的产量SCA效应值分别为5.881、6.088、5.854、3.804。
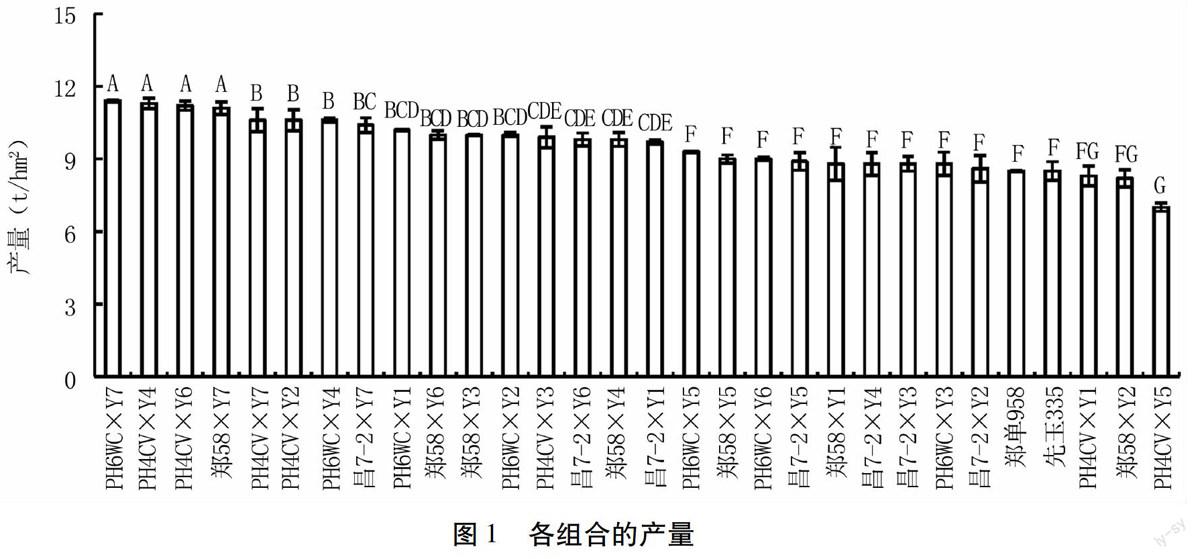
对28个组合的产量性状进行方差分析,结果见图1。其中,PH6WC×Y7的产量为11.4 t/hm2,比对照郑单958产量高33.5%,比先玉335产量高33.8%,且差异均达极显著水平。PH4CV×Y4的产量为11.3 t/hm2,比郑单958产量高32.8%,比先玉335产量高33.1%,且差异均达极显著水平。PH4CV×Y5的产量为7.0 t/hm2,比对照郑单958产量低17.6%,比先玉335产量低17.3%,且差异均达极显著水平。
3讨论与结论
配合力主要分为一般配合力(GCA)和特殊配合力(SCA)。其中,一般配合力是由加性效应引起的,特殊配合力是由非加性效应引起的[22-24]。一个自交系的GCA是根据该自交系作为亲本所组配的所有杂交种的平均表现来衡量的。一个组合的SCA是在已知双亲的GCA的基础上根据杂交种的表现差异来衡量的[25]。
对穗部性状和产量来说,它们的GCA 相对效应值越大越好,有利于改善杂交组合的穗部性状和产量,比较容易组配出高产杂交组合[26]。对7份新引抗粗缩病材料的穗部性状和产量一般配合力综合分析可以看出,由Y7配出的组合穗粗,穗行数多,百粒重高,小区产量高,易组配出高产组合。由Y4配出的组合穗较粗,百粒重高,小区产量高,也易组配出高产组合。由Y6配出的组合穗较长而粗,穗行数、行粒数较多,百粒重和小区产量较高,易组配出稳产组合。
特殊配合力是指两亲本所组配组合的水平,它的高低决定于亲本基因型的非加性基因效应,是杂交组合与其双亲平均表现基础上的预期结果的偏差,受外界环境条件的影响较大,不能在上下代间稳定遗传,但它可以指导杂种优势的利用和杂交种的选育[26]。本研究中,在组配的28个杂交组合中,产量SCA效应值变化范围是3.804~7.862。其中,效应值较大的组合有Y4×PH4CV、Y7×PH6WC、Y6×PH4CV、Y7×郑58,其效应值分别为7.862、7.831、7.781、7.623。同时,Y4×PH4CV、Y7×PH6WC、Y6×PH4CV、Y7×郑58的产量分别为11.3、11.4、11.2、11.1 t/hm2,均极显著地高于对照郑单958和先玉335。
杂交组合的优劣,不仅取决于亲本自交系的GCA相对效应值的大小,还取决于其组配的杂交组合的SCA相对效应值的大小,GCA和SCA的综合表现是玉米杂交种选育中的一个重要参数。在组配的组合中,至少要有一个亲本的一般配合力相对效应值高,而且特殊配合力相对效应值也比较高,才能组配出强优势的组合。本试验结果显示28个杂交组合中,从GCA和SCA效应值来看,Y4×PH4CV、Y7×PH6WC、Y6×PH4CV、Y7×郑58等组合表现较好。
参考文献:
[1]徐云姬,顾道健,秦昊,等. 玉米灌浆期果穗不同部位籽粒碳水化合物积累与淀粉合成相关酶活性变化[J]. 作物学报,2015,41(2):297-307.
[2]张世煌,彭泽斌,李新海. 玉米杂种优势与种质扩增、改良和创新[J]. 中国农业科学,2000,33(增刊):34-39.
[3]王懿波,王振华,王永普,等. 中国玉米主要种质杂交优势利用模式研究[J]. 中国农业科学,1997,30(4):16-24.
[4]曾三省. 中国玉米杂交种的种质基础[J]. 中国农业科学,1990,23(4):1-9.
[5]番兴明,谭静,杨峻芸. 热带、亚热带外来玉米种质利用[J]. 西南农业学报,2000,13(1):107-111.
[6]王河成,段运平,石红卫. 热带亚热带种质不同导入量对玉米自交系配合力的影响[J].玉米科学,1995,3(3):9-11.
[7]Hallauer A R.玉米轮回选择的理论与实践[M]. 中国农业科学院作物育种栽培研究所.北京:农业出版社,1989.
[8]Qi H H,Huang J,Zheng Q,et al. Identification of combining ability loci for five yield-related traits in maize using a set of testcrosses with introgression lines[J]. Theor. Appl. Genet.,2013,126(2):369-377.
[9]Hallauer A R. Methods used in developing maize inbreds[J]. Maydica,1990,35(1):1-6.
[10]Huang J,Qi H H,Feng X M,et al. General combining ability of most yield-related traits had a genetic basis different from their corresponding traits per se in a set of maize introgression lines[J]. Genetica,2013,141(10/11/12):453-461.
[11]杨引福,郭强,钱劲华. 8个玉米自交系主要穗部性状配合力的遗传分析[J]. 玉米科学,2008,16(3):30-33.
[12]李明顺,张世煌,李新海,等. 根据产量特殊配合力分析玉米自交系杂种优势群[J].中国农业科学,2002,35(6):600-605.
[13]王有芳,李少勇,王凌汉. 玉米果穗性状的配合力分析[J]. 玉米科学,2003,11(2):37-40,65.
[14]Lee E A,Tollenaar M. Physiological basis of successful breeding strategies for maize grain yield[J]. Crop Sci.,2007,47:202-215.
[15]王安乐,赵德法,陈朝辉,等. 玉米自交系抗粗缩病特性的遗传基础及轮回选择效应研究[J]. 玉米科学,2000,8(1):80-82.
[16]邸垫平,易晓云,苗洪芹,等. 玉米粗缩病抗性遗传研究[J]. 植物病理学报,2012,42(4):404-410.
[17]薛林,张丹,徐亮,等. 玉米抗粗缩病自交系种质的发掘和遗传多样性及其在育种中的应用[J]. 作物学报,2011,37(12):2123-2129.
[18]蒋飞. 玉米抗粗缩病材料的鉴定与筛选[D]. 泰安:山东农业大学,2011.
[19]陶永富,刘庆彩,徐明良. 玉米粗缩病研究进展[J]. 玉米科学,2013,21(1):149-152.
[20]李明顺. 13个玉米群体的配合力和遗传多样性分析[D]. 北京:中国农业科学院,2005.
[21]郭平仲. 数量遗传分析[M]. 北京:首都师范大学出版社,1993.
[22]Sprague G F,Tatum L A. General vs. specific combining ability in single crosses of corn[J]. J. Am. Soc. Agron.,1942,34:923-932.
[23]Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems[J]. Aust. J. Biol. Sci.,1956,9:463-493.
[24]Lv A Z,Zhang H,Zhang Z X,et al. Conversion of the statistical combining ability into a genetic concept[J]. Journal of Integrative Agriculture,2012,11(1):43-52.
[25]Xu Y B. Molecular plant breeding[M/OD]. http://www.cabi.org,2010.
[26]祁志云,杨华,李淑君,等. 优异玉米种质产量性状配合力分析[J]. 湖北农业科学,2014,53(7):1497-1501.
