‘石硖’龙眼果皮发育过程中糖代谢及相关酶活性变化
2016-05-30张红娜李建光舒波杨为海邓旭决登伟石胜友刘丽琴
张红娜 李建光 舒 波 杨为海 邓 旭 决登伟 石胜友 刘丽琴
摘 要 为探索龙眼果皮发育过程中糖分积累以及相关酶活性的变化规律,以‘石硖龙眼(Dimocarpus longan Lour. cv‘shixia)为试材,对花后45 d到果实成熟期间果皮质量、厚度,果糖、葡萄糖、蔗糖含量,以及蔗糖代谢相关酶活性水平进行测定。结果表明,果实发育后期,果皮质量增加的速度变缓,果皮厚度则表现为由厚到薄的变化模型。糖分水平显示,果实发育过程中果皮可溶性糖主要包括果糖、葡萄糖和蔗糖,同期果糖含量远远高于葡萄糖,蔗糖含量较低。蔗糖代谢酶活性表明,龙眼果皮中NI和SS(分解方向)活性较高,为调控‘石硖龙眼果皮糖代谢的主要酶类,SPS、SS(合成方向)活性则较低,为果糖和葡萄糖生成发挥非常重要的作用。
关键词 果皮;糖积累;相关酶活性
中图分类号 S667.2 文献标识码 A
Abstract In order to clear the variations both sugar accumulate and sugar metabolizing enzymes activities in longan pericarp in the process of fruit development stage,the weight and thickness of the pericarp, the concentration of fructose, glucose and sucrose, and the activities of the sucrose metabolizing enzymes were all measured in the pericarp of‘shixialongan during the stage that 45 d after flowering to fruit maturity. The results showed the growth rate of pericarp weight was from quickly to slowly and the thickness development model was thin~thick~thin. The sugar mainly included fructose, glucose and sucrose. The content of fructose was significant higher than glucose,however the content of sucrose was at low level during the whole growing period. The activities of NI and SS(the decomposition direction)maintained higher level, which were the main enzyme mediating the sugar accumulated in the pericarp, SPS and SS(the composition direction)with lower activity played very important role in accumulation of fructose and glucose that activities were lower.
Key words Longan pericarp; Sugar accumulate; Sugar metabolizing enzymes activities
doi 10.3969/j.issn.1000-2561.2016.06.002
龍眼(Dimocarpus longan Lour.)是我国南亚热带特产水果,其果实营养佳美,风味独特。一般认为龙眼果实的品质主要是果肉,所以大量的研究工作集中在果肉品质形成和采后保鲜等方面,而有关果皮的研究较少,少量报道仅限于保鲜技术、褐变机理、果皮结构与化学成分等方面[1-6]。林河通等[2]认为龙眼果皮与果实的耐藏性、抗病性和耐运性有关。有学者对龙眼果皮化学成分研究发现,龙眼果皮中有多糖、三萜、黄酮、香豆素、多酚、木栓酮、没食子酸、鞣花酸等次生物质[5-6]。此外未见龙眼果皮的相关报道。
糖是果实品质和风味的重要物质,也是其他次生物质合成的基础原料,在果树生长发育中有重要的生理功能,如为果实细胞膨大提供渗透推动力,通过复杂的信号转导调节植物中许多与生长发育相关基因的表达等。多数糖代谢方面的研究均集中在果肉及其他可食部分[7-10],在果皮的研究仅在柑橘[11]、苹果[12]、西瓜[13]等几种水果有少量报道,有关龙眼果皮糖代谢的研究还未见报道。据于安民[14]的观点,糖代谢是整个生物代谢的中心,沟通了蛋白质代谢、脂类代谢、核酸代谢及各类次生物质的代谢,因此本研究以‘石硖龙眼果皮为试材,对生长发育过程中糖分积累和蔗糖代谢相关酶活性变化进行研究,旨在为探索龙眼果实糖代谢的机理提供理论依据,也为进一步研究龙眼果皮物质的代谢提供参考。
1 材料与方法
1.1 材料
供试果实采自中国热带农业科学院南亚热带作物研究所(广东湛江)龙眼园内10年生的‘石硖龙眼。选择无病虫害生长健壮花期一致的植株5棵,挂牌标记,花后15 d开始采样,每10 d采样1次,直至果实成熟。每棵树每次从不同方向随机采集10个果实,混合所有果样。根据龙眼果实的发育特点,授粉受精后果皮和种皮开始发育,此期果皮和种皮难以分离,从花后45 d开始剥离果皮。剥取的果皮即时液氮冷冻,置于-80 ℃冰箱中保存备用。试验重复3次。
1.2 方法
1.2.1 果实发育动态研究 每次采样从混合果样中随机选取10个果实,用电子天平测定果皮质量,用游标卡尺测定果皮厚度。
1.2.2 糖的提取和测定 可溶性总糖的测定采用马志良[15]的蒽酮法。果糖、葡萄糖、蔗糖、麦芽糖、木糖、乳糖、甘露糖、山梨醇的提取參照赵智中[16]的方法,糖含量测定采用高效液相色谱(HPLC)法,色谱条件为:Series200氨基酸柱,柱温35 ℃,流动相:乙腈 ∶ 水=85 ∶ 15(V/V),流速1 mL/min,示差检测器及其相应的数据处理软件。所用的标准样均为分析纯样品。
1.2.3 酶的提取 蔗糖代谢相关酶的提取参照赵智中[16]的方法,测定参照张秀梅等[7]的方法。所有的操作均在低温进行。3次重复。
2 结果与分析
2.1 果皮发育动态
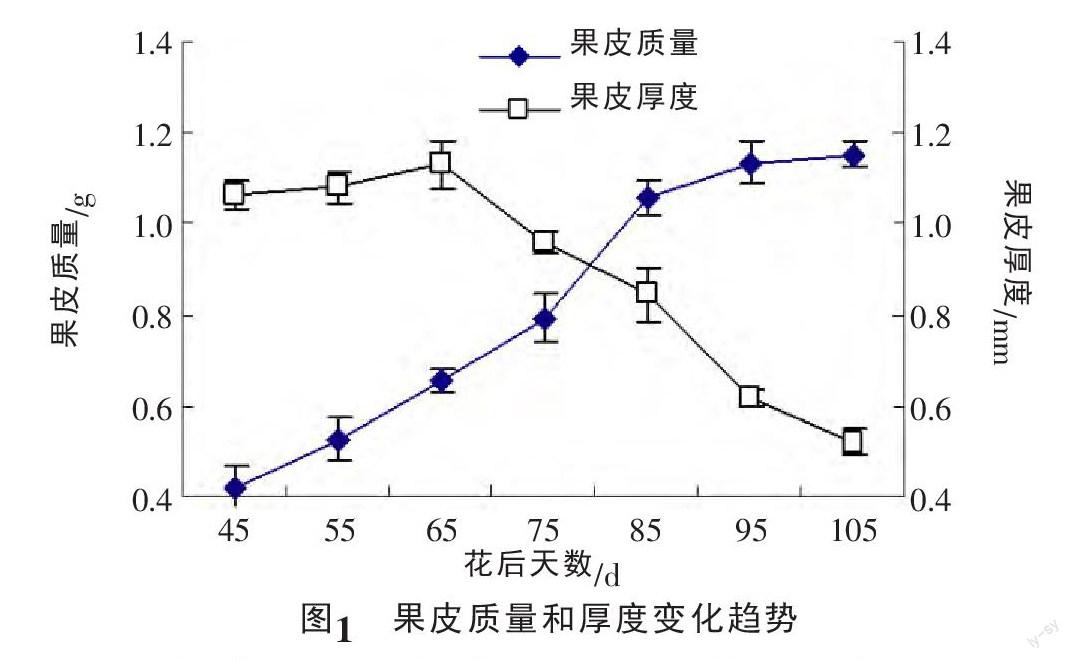
龙眼授粉受精后,果皮和种皮开始发育,此期果皮和种皮持续增大变厚,但难以将二者分离。果皮从花后45 d开始剥取,由图1可以看出,果皮质量从花后45 d开始急速增加,持续40 d左右,到85 d开始缓慢增加,呈现由快到慢的增长趋势。果皮厚度呈现由厚到薄的生长趋势,花后65 d达到最厚,约为1.131 mm,之后就迅速减小,到果实成熟期仅为0.520 mm,约为最大厚度的一半。果皮颜色在幼果期呈现绿色,之后叶绿素逐渐减少,红色素逐渐增加,到成熟时果皮呈现黄褐色。
2.2 果皮糖含量的变化
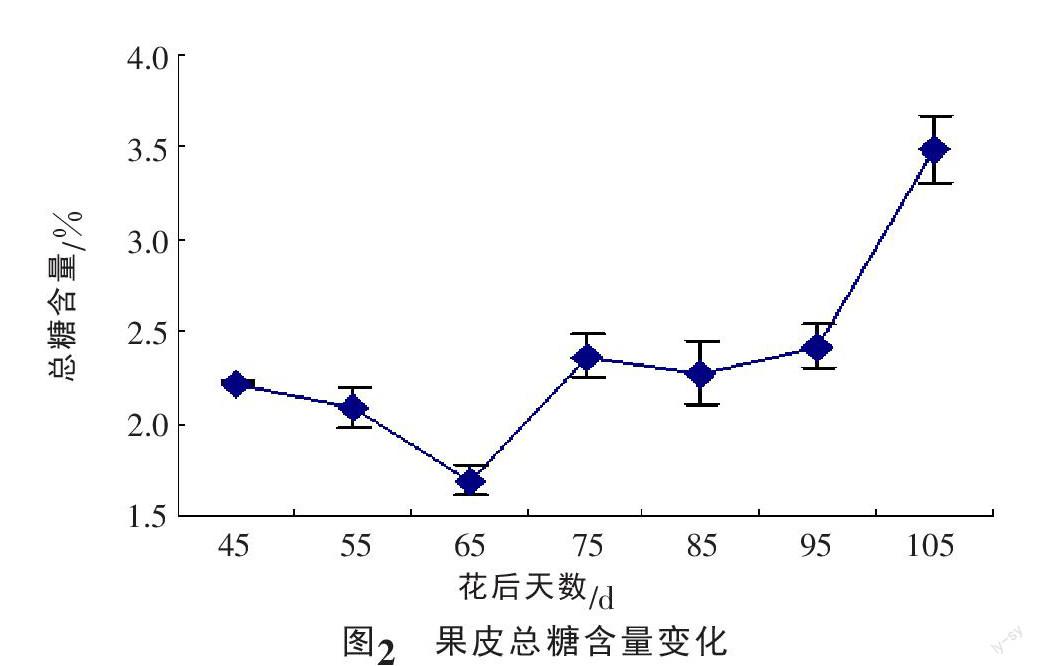
果皮可溶性总糖含量呈先降低后升高的变化趋势,前期降低速度缓慢,花后65 d达到最低峰值1.69%,之后开始急速上升,到成熟期达到生长发育期的最大值3.49%(图2)。
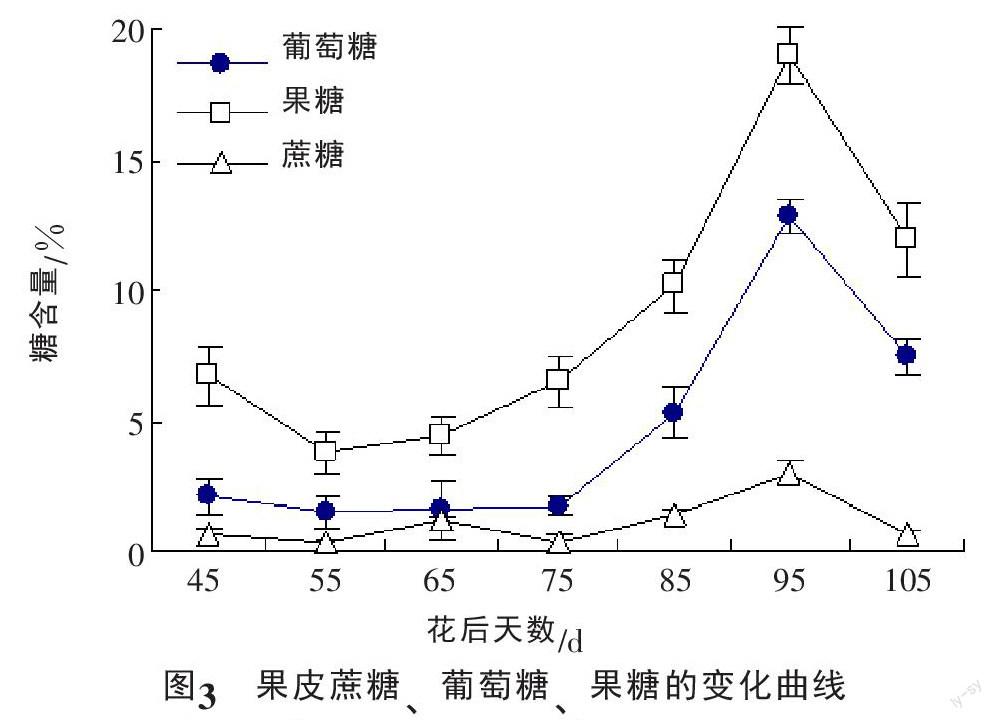
高效液相色谱检测结果显示,龙眼果皮中的可溶性糖主要包括果糖、葡萄糖和蔗糖,另外有微量麦芽糖,但由于麦芽糖峰面积较小,峰型和分离度差,没有进行统计分析。此外没有检测到其他糖的峰面积。由图3可以看出,果糖含量远远高于葡萄糖,从初期的6.71 mg/g急剧降低至生长发育期的最小值3.75 mg/g,而后逐渐增大,到花后95 d达到最高值19.01 mg/g,随着果实成熟,又急剧下降至11.98 mg/g。葡萄糖的变化呈“降低~升高~降低”的趋势,花后55 d前,葡萄糖含量有小幅度降低,之后趋于平缓,随着假种皮快速生长,果皮葡萄糖含量迅速上升,到花后95 d达到最大值12.88 mg/g,而后又急剧降低。蔗糖在整个生长发育期处于较低水平,呈波浪状曲线变化,生长发育初期和成熟期含量差异不显著。可见‘石硖龙眼果皮在生长发育过程中以积累单糖为主。
2.3 蔗糖代谢相关酶活性的变化
2.3.1 AI和NI活性的变化 ‘石硖 龙眼果皮发育过程中转化酶的活性见图4。NI活性从较高水平不断降低,花后75 d前,下降速度较平缓,之后维持在较稳定水平,10 d后开始大幅度下降,到果实成熟期,达到生长发育期的最小值8.64 μmol/(h·g FW)。AI活性处于较低水平,呈“降低~升高~降低”的变化趋势。花后45 d以后,活性缓慢降低,到花后65 d,活性降低至生长发育期的最小值4.27 μmol/(h·g FW),之后急速升高,到花后85 d AI活性达到生长极值14.88 μmol/(h·g FW),之后又开始降低,果实成熟期,与NI活性差异不显著。
2.3.2 SPS和SS活性的变化 由图5可见,SS分解蔗糖方向的活性变化趋势较为复杂,花后55 d前,活性从较低水平快速升高到最高峰值27.70 μmol/(h·g FW),为生长初期的3.8倍,之后迅速降低,到花后65 d又有小幅度升高,花后75 d后又持续降低,到果实成熟时达到15.76 μmol/(h·g FW)。合成蔗糖方向,SS活性一直处于较低水平,除65~75 d有小幅度降低以外,基本呈上升趋势,到果实成熟期达到最大,为6.80 μmol/(h·g FW)。龙眼果皮SPS活性呈先升高后降低的变化趋势,花后55 d之前,SPS活性基本保持不变,之后快速升高达到峰值13.24 μmol/(h·g FW),而后一直呈缓慢下降趋势,直至果实成熟时,SPS活性为6.41 μmol/(h·g FW)。
3 讨论
本研究发现,随着后期果实不断变大,果皮质量不断增加,而果皮却不断变薄。依据陈清西等[17]的观点,果皮生长与假种皮发育密切相关,假种皮生长对种皮有一定的挤撑作用,是导致种皮后期变薄的主要原因。果皮质量呈现不断增加的变化趋势,这与果糖、葡萄糖和蔗糖的积累变化趋势不一致,可见果皮中除了糖分积累以外,还有其他成分积累,有待进一步研究。
本研究中‘石硖龙眼果皮是以积累单糖(果糖和葡萄糖)为主,主要受NI和SS(分解方向)的调控和影响;而笔者对‘石硖龙眼果肉糖分积累的研究表明,果肉是以积累蔗糖为主,起主导作用的酶为AI和SS(分解方向),二者有较大差异。这说明,果实发育的不同阶段和不同部位,糖分积累的方式是不同的,这与前人在柑橘、番茄、葡萄等的报道相一致[11,16,18-19]。龙眼果肉和果皮的不同糖分积累和代谢,是内在的遗传特性和外在的因子等因素相互作用的结果。但是目前植物中蔗糖从源到库运输的全部生理途径并没有完全阐明,是研究的主要课题。
笔者发现,SS(分解方向)在龙眼果肉和果皮中均起主导作用,这与钱琼秋等[13]在西瓜的研究一致。此外,龙眼果皮起主导作用的酶NI和SS(分解方向)的活性高于果肉起主导作用的酶AI和SS(分解方向)的活性;而糖分积累量却远远低于果肉。笔者推测,果肉在生长发育前期,较高活性的AI促进蔗糖分解为己糖,主要用以维持胞间和液胞内外高的蔗糖浓度梯度,从而促进物质的韧皮部卸载和跨膜运输,来构建各种细胞器,而生长后期,细胞器已构建完成,AI主要促进糖分积累;而果皮呼吸代谢较强,较高活性的NI催化蔗糖分解为己糖,少部分糖分用于贮藏积累,大部分用于为呼吸代谢提供碳源和果皮其他物质合成提供能量。故果皮的酶活性高于果肉,而糖分积累量却低于果肉。蔗糖代谢相关酶在不同作物不同时期甚至不同部位的生理作用和活性大小均不相同,糖分积累和代谢是各种酶综合作用的结果,目前龙眼蔗糖代谢酶的生理功能和作用机制还没有完全阐明,有待于进一步证实。
4 结论
‘石硖龍眼果皮在生长发育过程中以积累单糖为主,可溶性糖主要包括果糖、葡萄糖和蔗糖,同期果糖含量远远高于葡萄糖,蔗糖在整个生长发育期处于较低水平。果皮生长发育过程中,蔗糖代谢相关酶NI和SS(分解方向)是调控‘石硖龙眼果皮糖代谢的主要酶类。高活性的NI和SS(分解方向)均有利于蔗糖分解成果糖和葡萄糖,这与果皮生长发育过程中以积累单糖(果糖和葡萄糖)为主相一致。而AI、SPS和SS(合成方向)活性较低,低活性的SPS和SS(合成方向)也有利于果糖和葡萄糖积累,AI活性较低则有利于蔗糖积累。
参考文献
[1] 林河通, 陈绍军, 席玙芳, 等. 龙眼果皮微细结构的扫描电镜观察及其与果实耐贮性的关系[J]. 农业工程学报, 2002, 18(3): 95-99.
[2] 林河通, 席玙芳, 陈绍军, 等. 龙眼果实采后失水果皮褐变与活性氧及酚类代谢的关系[J]. 植物生理与分子生物学学报, 2005, 31(3): 287-297.
[3] 刘熙东, 吴振先, 韩冬梅, 等. 龙眼采后果皮细胞壁代谢相关酶活性的变化[J]. 热带作物学报, 2006, 27(2): 24-28.
[4] 邓斯聪, 杨为海, 朱效传, 等. 龙眼果皮发育解剖学观察[J]. 果树学报, 2008, 25(2): 193-197.
[5] 郑公铭. 龙眼果实的化学成分及其生物活性的研究[D]. 广州: 中国科学院华南植物园, 2009.
[6] 陈丽华. 龙眼壳多糖的提取、 分离纯化与性质研究[D]. 重庆:西南大学, 2009.
[7] 张秀梅, 杜丽清, 孙光明, 等. 菠萝果实发育过程中糖积累与其代谢酶的关系[J]. 热带作物学报, 2008, 29(1): 10-13.
[8] 王惠聪, 黄辉白, 黄旭明, 等. 荔枝果实的糖积累与相关酶活性[J]. 园艺学报, 2003, 30(1): 1-5.
[9] 武红霞, 邢姗姗, 王松标, 等. ‘台农1号 芒果果实发育过程中的糖分积累与相关酶活性研究[J]. 西北植物学报, 2011, 31(9): 1 811-1 815.
[10] 张中霞, 刘 艳, 白立华, 等. 河套蜜瓜果实发育过程中糖积累与蔗糖代谢相关酶的关系[J]. 西北植物学报, 2011, 31(1): 123-129.
[11] 陈俊伟, 张上隆, 张良诚, 等. 柑橘果实遮光处理对发育中的果实光合产物分配、糖代谢与积累的影响[J]. 植物生理学报, 2001, 27(6): 499-504.
[12] 宋 哲, 李天忠, 徐贵轩, 等. ‘富士 苹果着色期果皮花青苷与果实糖份及相关酶活性变化的关系[J]. 中国农学通报, 2008, 24(4): 255-260.
[13] 钱琼秋, 刘慧英, 朱祝军, 等. 嫁接西瓜果实发育过程中蔗糖代谢及相关酶调控的研究[J]. 浙江大学学报(农业与生命科学版), 2004, 30(3): 285-289.
[14] 于安民, 张 敏, 杨锦芬, 等. 阳春砂果实发育过程中可溶性糖及蔗糖代谢酶活性变化规律的研究[J]. 世界科学技术-中医药现代化, 2014(7): 1 497-1 504.
[15] 张志良, 瞿伟菁. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2003.
[16] 赵智中, 张上隆, 徐昌杰, 等. 蔗糖代谢相关酶在温州蜜柑果实糖积累中的作用[J]. 园艺学报, 2001, 28(2): 112-118.
[17] 陈清西,廖镜思,胡又厘. 龙眼果实生长曲线和各组织的相关分析[J]. 福建农业大学学报, 1995, 24(1): 19-22.
[18] 齐红岩, 李天来, 刘海涛, 等. 番茄不同部位中糖含量和相关酶活性的研究[J]. 园艺学报, 2005, 32(2): 239-243.
[19] 孙凌俊, 赵文东, 郭修武, 等. 晚红葡萄着色期光合产物14C的分配规律[J]. 中国农学通报, 2010, 26(14): 186-189.
