三种增温情景对入侵植物紫茎泽兰种子出苗的影响
2016-05-17彭扬李景吉彭培好
彭扬, 李景吉, 彭培好
三种增温情景对入侵植物紫茎泽兰种子出苗的影响
彭扬, 李景吉, 彭培好*
成都理工大学生态资源与景观研究所, 成都 610059
全球气候变暖影响外来植物的入侵风险, 不同增温情景可影响入侵植物的生长和其他表型。种子出苗是种群生活史的重要阶段, 但是目前还不了解不同增温情景对入侵植物种子出苗的影响。设置三种增温情景(白天增温、晚上增温、全天增温), 通过同属近缘种比较, 研究不同增温对入侵植物紫茎泽兰(Eupatorium adenophorum)和本地植物异叶泽兰(Eupatorium heterophyllum)种子出苗的影响。与对照(不增温)相比, 三种增温方式都不同程度地降低异叶泽兰的出苗率, 白天增温时出苗率最低, 其次是全天增温; 对于紫茎泽兰而言, 晚上增温提高其出苗率, 白天增温降低其出苗率, 在全天增温条件下紫茎泽兰没有出苗。增温对紫茎泽兰的影响比对异叶泽兰的影响更明显。此外, 异叶泽兰与紫茎泽兰种子出苗率与积温基本呈线性关系, 且不同增温方式对各线性关系影响存在差异。这些结果初步表明, 如果未来的气候变暖发生在夜间, 则有利于提高紫茎泽兰的入侵风险; 如果发生在白天、尤其是全天, 则可能极大降低其入侵风险。
紫茎泽兰; 气候变暖; 增温情景; 出苗率; 入侵植物; 同属近缘种比较
1 前言
全球变暖已成为不争的事实, 这种变化正深刻影响着各种有机体的新陈代谢[1]。全球变暖主要由人类活动产生[2], 随着人口增长和人类活动的增加,全球变暖的过程仍在继续。据姜大膀等估计[3], 在未来50年内, 全球平均气温将增加2 ℃左右。早在20年前, Easterling[4]通过微分变元, 预测在全球变暖的大环境下, 昼夜温差将会缩小, 即全球气候变暖主要表现为夜间气温升高[5], 国内研究也得出类似结果[6]。也就是所谓的非对称性增温[7]。这种非对称性增温可能影响植物光合作用和呼吸作用, 并进一步影响植物的生长与碳积累[8]。尽管全球变暖的速度还存在诸多争议, 但全球变暖已经从不同的时空尺度对人类赖以生存的地球生态环境产生了深刻影响, 增温会为入侵种的扩增提供更为有利的条件,从而增加入侵的危害性。研究表明入侵植物往往具有较强的耐受性和适应能力, 微小的温度变化都可能给它们带来巨大的竞争优势[9]。全球气候变暖,势必将影响到植物的生理生态特征, 会引起一部分物种的灭绝或者地理迁移, 进而影响植物的种群、群落、生态系统甚至对整个生态系统也会产生巨大的影响。
气候因子是影响植物生长的重要因素之一, 对入侵植物的入侵机制、入侵速率以及分布区域有一定的影响; 其中温度是一个很重要的生态因子, 当大气温度发生变化时, 植物会对其变化做出适应性的调整, 即植物的生长、发育及繁殖对温度的变化有着极强的适应性和可塑性[10]。入侵杂草一般都有较宽的生理生态耐受性或可塑性, 其生长和形态属性对环境变化的响应明显[11]。国内外对非对称性增温早有大量报道, 但在不同情景增温对入侵植物的影响方面却研究很少[12–13]。我们早期研究过三种增温情景(白天增温、晚上增温、全天增温)对入侵植物空心莲子草克隆整合与形态可塑性的影响[12–13]。结果显示, 增温影响空心莲子草的克隆整合能力, 三种增温情景中全天增温影响最显著; 空心莲子草表现出对增温环境的可塑性变化, 这种形态的可塑性变化依赖增温情景, 夜间增温可能促进空心莲子草的入侵。
由于全球气候变暖, 一些适应性强的物种会入侵到其他领域, 排挤当地的物种, 从而改变当地物种的多样性和生态系统[14]。这意味着全球气候的变暖, 势必将影响到植物的生理生态特征, 会引起一部分物种的灭绝或者地理迁移, 进而影响植物的种群、群落、生态系统甚至对整个生态系统也会产生巨大的影响。在进行植物入侵过程研究时, 比较研究是常用的方法, 但以往对不同物种进行比较时往往忽略了亲缘关系对比较研究的影响[15]。入侵植物的近缘物种, 多为同属种, 在进化上与该入侵种处于同一地位, 往往表现出相近的生物学特征。将入侵种与其本地近缘种, 能够排除亲缘关系带来的影响, 更有利于揭示与入侵相关的性状[16–17]。
紫茎泽兰(Eupatorium adenophorum), 菊科(Compositae)泽兰属(Eupatorium), 多年生丛生型半灌木状草本植物。原产于中美洲, 19世纪曾作为观赏植物引种到欧洲, 后来引种到澳洲和亚洲, 上世纪40年代从中缅边境传入西双版纳后, 以平均每年20km的速度向东和向北快速蔓延扩散[18]。紫茎泽兰是我国外来入侵物种中危害最为严重的植物之一, 已被列入我国公布的第一批外来入侵物种名单之首[19]。种子出苗是植物生长周期中的重要阶段,它对种群个体的繁殖、扩展、更新和物种抵御不良环境有重要意义[20]。紫茎泽兰种子的产量惊人, 有性生殖在种群扩增方面有着极其重要的作用, 因此种子出苗率对紫茎泽兰入侵能力的影响是显而易见的。有实验发现, 与本地植物异叶泽兰相比, 紫茎泽兰种子萌发出苗对温度不敏感[21]。本实验以异叶泽兰与紫茎泽兰种子为材料, 采用红外线辐射加热器( Infrared radiators)加温, 比较不同方式(全天、白天、晚上)下增温2 ℃左右时异叶泽兰与紫茎泽兰种子出苗的不同, 旨在探讨昼夜增温不均匀对紫茎泽兰种子出苗过程及净率出苗的影响。
2 材料和方法
2.1实验地点
实验区地处四川省成都市, 地理坐标为30°40′41″N, 104°08′15″E, 海拔512 m, 年均温16.2 ℃, 年均降水量918.2 mm。该地地处亚热带湿润季风气候区,日照较少, 雨量充沛, 无霜期长, 四季分明。
2.2实验材料
实验所用异叶泽兰(E. heterophyllum)与紫茎泽兰(E. adenophorum)的种子均采自四川成都本地, 同种处理下每个物种的种子均采自同一株, 选取成熟饱满的种子, 可认为其自身出苗能力无差别[21]。
2.3实验设计
采集当地野生自由生长的异叶泽兰与紫茎泽兰种子, 选取成熟饱满的种子, 进行出苗实验。实验分四种处理, 即不增温、白天(7:00—19:00)增温、晚上(19:00—7:00)增温、全天增温, 每种处理五个对照,共2×4×5=40个实验盆, 每个实验盆均匀撒播50粒种子, 给予适宜的条件, 每天观察出苗情况。加热器采用HS-2408红外线辐射加热器(Kalglo Electronics, Bethlehem, PA, USA), 高度距离盆表面约1.5 m, 经测量调试后实验盆土壤表面增温约2 ℃[8]。实验处理相互之间相距约2 m, 以避免温度处理相互干扰。在对照组中设置一个铁皮制“虚拟加热器”来抵消加热器本身的荫蔽效应。
2.4观测指标及方法
实验于三月底开始, 持续两个周。实验期间每天11:00记录当日气温, 7:00和19:00记录出苗数, 并将每次出苗的种子全数剔除。每次出苗的种子数与每盆种子总数(50)之比, 可得出苗率; 将每天之前的种子出苗率相加, 可得该日累积出苗率; 将每日11:00测得的气温相加, 可得日积温; 三种增温处理相对于对照组(不增温)的出苗率变化值在对照组出苗率中所占比例, 定义为增温方式对出苗率的影响系数。
2.5统计分析方法
实验数据采用Excel、SPSS19.0进行统计、分析; 用单因素方差分析(One-way ANOVA)比较同一指标不同处理之间的差异显著性, 用双因素方差分析比较物种和不同增温处理对种子出苗率的影响;图形采用Sigma Plot 10.0软件绘制。
3 结果
3.1不同增温处理对异叶泽兰与紫茎泽兰种子出苗过程的影响
日累积出苗率与时间关系如图1所示。由图可知, 随着时间推移, 累积出苗率曲线由快速增长逐渐趋于稳定, 第8天后基本保持不变; 除全天增温下紫茎泽兰未见出苗外, 不同增温情景并未使其曲线形状发生实质性的改变。将日积温与异叶泽兰和紫茎泽兰出苗率百分比数值取对数, 其一一对应关系如图2所示。由图可知, 两对数基本呈线性关系(R²>0.799), 即说明, 出苗率与积温呈线性关系。令y=ln(100出苗率), x=ln(积温), 取其趋势线并分析可得表1。由图表可知, 对于异叶泽兰, 增温后的直线截距变小, 斜率变大; 而对于紫茎泽兰, 白天增温后直线截距变小, 斜率变大, 晚上增温后直线截距变大, 斜率变小; 总体上不同增温情景对直线斜率影响不显著(P>0.05), 且对紫茎泽兰影响较小。
3.2不同增温处理对异叶泽兰与紫茎泽兰种子净出苗率的影响

图1 四种气温处理下每日异叶泽兰与紫茎泽兰种子累积出苗率(均值±标准误)Fig. 1 The daily accumulative emergence rate of E. adenophorum and E. heterophyllum under four air temperature regimes (mean± SE)
三种增温处理相对于对照组(不增温)的出苗率变化值在对照组出苗率中所占比例, 定义为增温方式对出苗率的影响系数。不同增温处理对异叶泽兰与紫茎泽兰种子总出苗率的影响如图3所示。由图可知, 与对照(52.8%)相比, 三种增温方式都会不同程度地降低异叶泽兰的出苗率, 白天增温时出苗率最低(32.4%), 其次是全天增温(39.6%), 晚上增温时出苗率(48.8%)略微减小; 而对于紫茎泽兰, 与对照(9.2%)相比, 晚上增温能提高其出苗率(11.6%), 白天增温时大幅降低(1.6%), 而在全天增温条件下,紫茎泽兰没有出苗。总体上异叶泽兰出苗率(43.4%)明显高于紫茎泽兰(5.6%); 除晚上增温时的紫茎泽兰外, 增温抑制植物的出苗, 且对紫茎泽兰影响系数绝对值更大。双因素方差分析(表2)表明: 相对于不同的增温方式, 物种的变换对出苗率的影响更显著; 增温对净种子出苗的影响是边际显著的; 增温和物种之间存在强烈的交互作用。

图2 四种气温处理下积温与异叶泽兰和紫茎泽兰种子出苗率的关系Fig. 2 The relations between accumulative temperatures and accumulative emergence rates of E. adenophorum and E. heterophyllum under four air temperature regimes

表1 四种气温处理下积温与异叶泽兰和紫茎泽兰种子出苗率的关系方程Tab. 1 The relation equations of accumulative temperatures and emergence rates of E. adenophorum and E. heterophyllum under four air temperature regimes
4 讨论
4.1不同增温情景对种子出苗过程的影响

图3 不同气温处理对异叶泽兰与紫茎泽兰种子净出苗率及其影响系数比较Fig. 3 Comparisons of the net emergence rates and the influence coefficients on the emergence rates of E. adenophorum and E. heterophyllum under different air temperature regimes
种子出苗是植物成功定居的关键阶段[22]。然而,我们对入侵植物这个过程的了解依然很少。我们的实验室在野外条件下进行的, 所以更加接近自然状况。本次实验通过设置三种增温情景(白天、晚上、全天增温), 记录入侵种紫茎泽兰及本地种异叶泽兰的种子出苗率随时间变化, 分析不同增温对两物种种子出苗过程的影响。结果显示: 累积出苗率随时间推移趋于稳定, 除全天增温下紫茎泽兰未见出苗外, 不同增温情景只影响两植物种子出苗率, 而并未使其出苗过程发生实质性的改变; 出苗率与积温呈线性关系, 对于异叶泽兰, 增温后的直线截距变小, 斜率变大; 而对于紫茎泽兰, 白天增温后直线截距变小, 斜率变大, 晚上增温后直线截距变大,斜率变小; 总体上不同增温情景对直线斜率影响不显著, 且对紫茎泽兰影响较小。
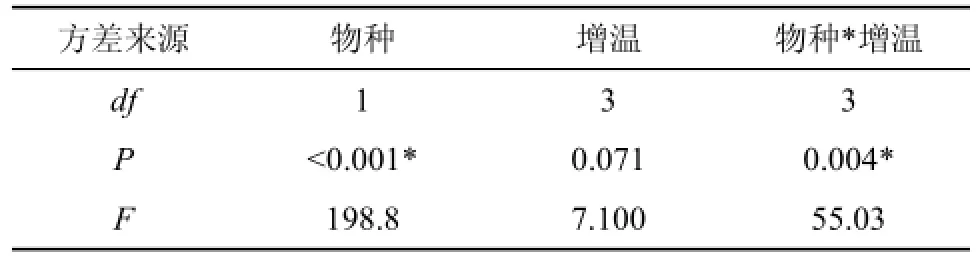
表2 物种、增温和二者交互效应对种子出苗率的影响Tab. 2 Effects of species, experimental warming and their interactions on the net emergence rate
种子的出苗包括两个过程, 即种子萌发和胚芽破土。在特定的环境下, 种子达到其最大萌发率并破土而出所需的时间是一定的[23]。温度对所有种子的影响是相同的, 种子在不同增温情境下出苗几率是不同的[24], 但总体上种子的出苗时间受增温情景影响不显著。以y=ln(100出苗率)、x=ln(积温)建立坐标系, 对于异叶泽兰, 增温后的直线截距变小,斜率变大, 意味着在较大积温时, 增温不再降低其出苗率, 而是使其升高。对于紫茎泽兰, 晚上增温后直线截距变大, 斜率变小, 说明在较小积温时, 晚上增温提高其出苗率, 即有利于提前其生长节律,从而在竞争中取得主动。总体上看, 不同增温情景只影响种子的出苗率, 而对种子出苗过程的影响不显著。
4.2不同增温情景对种子净出苗率的影响
不同增温情景对种子出苗过程的影响不显著,但对种子净出苗率的影响是显而易见的。实验结果显示: 与不增温对照相比, 全天增温情景下, 异叶泽兰出苗率明显降低, 紫茎泽兰则未见出苗; 白天增温情景下, 紫茎泽兰与异叶泽兰的出苗率都大幅降低; 晚上增温情景下, 异叶泽兰出苗率明显降低,紫茎泽兰出苗率升高; 与本地种异叶泽兰比较, 紫茎泽兰出苗率对增温响应更明显; 相对于不同的增温方式, 物种的变换对出苗率的影响更显著, 增温和物种之间无交互作用。
紫茎泽兰出苗率明显低于异叶泽兰, 可能是由于实验时种子播种过深、土壤水分过低、实验光照过弱、实验期与其生长节律冲突等原因造成的[25]。但考虑到实验变量控制的严格性, 增温对种子出苗率的影响数据是可信的。在全天增温与白天增温情境下, 紫茎泽兰出苗率大幅降低, 且降低幅度大于本地种异叶泽兰。白天气温相对夜间较高, 当白天对其进行增温时, 过高的温度可能会超出种子萌发的最适温度, 造成种子萌发环境干旱化, 降低种子出苗率[26]。紫茎泽兰种子比异叶泽兰较小, 受增温影响更为明显。而只有夜间增温时, 昼夜温差减小,增加了紫茎泽兰种子出苗所需的有效积温, 有利于提高其净出苗率。
4.3不同增温的生态学意义
种子出苗是植物生长周期中的重要阶段, 它对种群个体的繁殖、扩展、更新和物种抵御不良环境有重要意义[20]。对于入侵植物来说, 种子出苗率越高, 新的幼苗就越多, 种群数量就越大, 入侵风险就越高[22]。不同增温情景对植物的生长发育的影响是不同的[27]。白天增温能拉大昼夜温差, 夜间增温能缩小昼夜温差, 而全天增温可以不改变昼夜温差的情况下提高积温。实验发现, 夜间增温能为入侵植物紫茎泽兰提供更好的出苗环境, 从而提高其入侵风险; 白天增温与全天增温会不同程度的抑制其出苗, 进而降低其入侵风险。研究结果表明, 在未来一段时间内, 以夜间增温为主的全球变暖更有利于紫茎泽兰提高其出苗率, 增加入侵风险。随着人类活动的日益增多, 在今后的一段时间内, 气温增加是显而易见的。在全球变暖的大环境下, 外来种入侵的生理学及生态学过程还有待更进一步研究, 但防治入侵生物的重要性是毋庸置疑的。
[1] MICHAEL E D, GEORGE W, RAYMOND B H. Global metabolic impacts of recent climate warming[J]. Nature, 2010, 467(7): 704–707.
[2] 陈宜瑜, 陈泮勤, 葛全胜, 等. 全球变化研究进展与展望[J].地学前缘, 2012, 9(1): 11–18.
[3] 姜大膀, 富元海. 2°C全球变暖背景下中国未来气候变化预估[J]. 大气科学, 2012, 36(2): 234–236.
[4] EASTERLING D R, HORTON B, JONES P D, et al.Maximum and minimum temperature trends for the globe.[J]. Science, 1997, 277(18): 364–367.
[5] 房世波, 谭凯炎, 任三学. 夜间增温对冬小麦生长和产量影响的实验研究[J]. 中国农业科学, 2010, 43(15): 3251–3258.
[6] 翟盘茂, 任福民. 中国近四十年最高最低温度变化[J].气象学报, 1997, 55(4): 418–429.
[7] 谭凯炎, 房世波, 任三学, 等. 非对称性增温对农业生态系统影响研究进展[J]. 应用气象学报, 2009, 20(5): 634–641.
[8] WAN Shiqiang, XIA Jianyang, LIU Weixing, et al. Photosynthetic overcompensation under noctural warming enhances grassland carbon sequestration[J]. Ecology, 2009, 90(10): 2700–2710.
[9] ALPERT P, BONE E, HOLZAPFEL C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants[J]. Perspect Plant Ecology, 2000, 3(1): 52–66.
[10] WOOWARD F I. Climate and plant distribution[M]. Cambridge: Cambridge University Press, 1987.
[11] 刘勤, 赖星竹, 彭培好, 等. 模拟气候变暖对不同地区多斑矢车菊形态属性的影响[J]. 西北植物学报, 2011, 31(10): 2078–2084.
[12] LI Jingji, PENG Peihao, HE Weiming. Physical connection decreases benefits of clonal integration inAlternanthera philoxeroidesunder three warming scenarios[J]. Plant Biology, 2012, 14(2): 265–270.
[13] 褚延梅, 杨健, 李景吉, 等. 三种增温情景对入侵植物空心莲子草形态可塑性的影响[J]. 生态学报, 2014, 34(6): 1411–1417.
[14] 珊丹. 控制性增温和施氮对荒漠草原植物群落和土壤的影响[D]. 内蒙古: 内蒙古农业大学, 2008, 125.
[15] 王坤, 杨继, 陈家宽. 近缘种比较研究在植物入侵生态学中的应用[J]. 生物多样性, 2009, 17(4): 353–361.
[16] MACK R N. Predicting the identity and fate of plant invaders: emergent and emerging approaches[J]. Biological Conservation, 1996, 78(1): 107–121.
[17] VAN Kleunen M, DAWSON W, SCHLAEPFER D, et al. Are invaders different? A conceptual framework of comparative approaches for assessing determinants of invasiveness[J]. Ecology Letters, 2010, 13(8): 947–958.
[18] WANG Rui, WANG Yinzheng. Invasion dynamics and potential spread of the invasive alien plant speciesAgemtina adenophom(Asteraceae) in China[J]. Diversity and Distributions, 2006, 12(4): 397–408.
[19] 张修玉, 许振成, 宋巍巍, 等. 紫茎泽兰入侵地的生物多样性[J]. 生态环境学报, 2010, 19(7): 1525–1531.
[20] 韩利红, 刘潮, 郑玉龙. 紫茎泽兰与3种同属本地植物种子特性比较[J]. 种子, 2010, 2(2): 73–76.
[21] 张丽坤, 王朔, 冯玉龙. 紫茎泽兰种子形态特征和出苗特性与其入侵性的关系[J]. 生态学报, 2014, 34(13): 3584–3591.
[22] HARPER J L. Population biology of plants[M]. Blackburn Press, 2010.
[23] 王文琪, 王进军, 赵志模. 紫茎泽兰种子种群动态及出苗特性[J]. 应用生态学报, 2006, 17(6): 982–986.
[24] 倪文. 环境因子对杂草紫茎泽兰种子发芽的影响[J]. 生态学报, 1983, 3(4): 327–331.
[25] 董丽佳, 桑卫国. 模拟增温和降水变化对北京东灵山辽东栎种子出苗和幼苗生长的影响[J]. 植物生态学报, 2012, 36(8): 819–830.
[26] TURNBULL M H, MURTHY R, GRIFFIN K L. The Relative impacts of daytime and night-time warming on photosynthetic capacity inPopulus deltoides[J]. Plat, Cell and Environment, 2002, 25(12): 1729–1737.
[27] 马树庆, 王琪, 吕厚荃, 等. 水分和温度对春玉米出苗速度和出苗率的影响[J]. 生态学报, 2012, 32(11): 3378–3385.
Three warming scenarios differentially affect the seed emergence of an invasive forb Eupatorium adenophorum
PENG Yang, LI Jingji, PENG Peihao*
Chengdu University of Technology,Ecological Resources and Landscape Research Institute,Chengdu610059,China
Global climate warming commonly affects the invasion risk of exotic plants, and diverse different warming scenarios can affect their growth and other traits. Although seed emergence is among the key stages of life cycle of plant populations, little is known about how different climate warming affects the seed emergence of invasive plants. We conducted a simulated climate warming experiment at Chengdu, in which the invasive forbEupatorium adenophorumand its native congenerE. heterophyllumwere subjected to three warming treatments: day-warming, night-warming, daily warming. We recorded the seed emergence every day and then analyzed how different warming scenarios affected it of the two species. Compared with the controls, three warming ways reduced the emergence rate ofE. heterophyllum, and the emergence followed the order: day-warming < daily warming < night-warming. ForEupatorium adenophorum,night-warming increased its seed emergence, day-warming decreased it, and no emergence was found in the daily warming. Overall, simulated warming had greater effects onEupatorium adenophorumthan onE. heterophyllum. Additionally, there were linear relationships between the emergence rate of these two species and accumulated air temperatures, and the effects of different warming ways on these linear relationships were variable. Our findings suggest that the nighttime-based climate warming may be advantageous forE. adenophorumto enhance its invasion risk, and the opposite may be true in the day-warming, particularly in the daily warming.
climate warming; congeneric comparison;Eupatorium adenophorum; invasive plants; seed emergence; warming scenarios
10.14108/j.cnki.1008-8873.2016.05.008
Q948.1
A
1008-8873(2016)05-050-06
彭扬, 李景吉, 彭培好. 三种增温情景对入侵植物紫茎泽兰种子出苗的影响[J]. 生态科学, 2016, 35(5): 50-55.
PENG Yang, LI Jingji, PENG Peihao. Three warming scenarios differentially affect the seed emergence of an invasive forbEupatorium adenophorum[J]. Ecological Science, 2016, 35(5): 50-55.
2015-07-11;
2015-08-20
国家青年科学基金项目(41501060)
彭扬(1992—), 男, 山东人, 研究生在读, 研究方向:生态环境与景观修复, E-mail: 195693858@qq.com
*通信作者: 彭培好, 男, 教授, 研究方向: 生态环境评价及生物多样性, E-mail: peihaop@163.com
