RBAP96 Mediates Radiosensitivity of Breast CancerCellsviaInteractingwithRetinoblastoma Protein
2016-05-13JunlingZhangXiaoleiXueQinghuiMengLuLuMingCuiSaijunFan
Junling Zhang,Xiaolei Xue,Qinghui Meng,Lu Lu,Ming Cui,Saijun Fan
Tianjin Key Laboratory of Radiation Medicine and Molecular Nuclear Medicine,Institute of Radiation Medicine,Chinese Academy of Medical Sciences and Peking Union Medical College,Tianjin 300192, China(Zhang JL,Lu L,Xue XL,Cui M,Fan SJ);Department of Oncology,Lombardi Comprehensive Cancer Center,Georgetown University,Washington DC 20079,USA(Meng QH)
·论著·
RBAP96 Mediates Radiosensitivity of Breast CancerCellsviaInteractingwithRetinoblastoma Protein
Junling Zhang,Xiaolei Xue,Qinghui Meng,Lu Lu,Ming Cui,Saijun Fan
Tianjin Key Laboratory of Radiation Medicine and Molecular Nuclear Medicine,Institute of Radiation Medicine,Chinese Academy of Medical Sciences and Peking Union Medical College,Tianjin 300192, China(Zhang JL,Lu L,Xue XL,Cui M,Fan SJ);Department of Oncology,Lombardi Comprehensive Cancer Center,Georgetown University,Washington DC 20079,USA(Meng QH)
ObjectiveTo identify a novel retinoblastoma protein(RB)-associated protein(RBAP 96) and to explore the impact of RBAP96 on radiosensitivity of human breast cancer cells.MethodsAn in vivo and in vitro association of RBAP96 with RB was determined by immunoprecipitation-Western blotting and GST pull-down assay.Protein expression was measured by Western blot assay.Cellular survival was evaluated by using a colony formation assay.ResultsIn both in vitro and in vivo assays,we found that the RBAP96 and RB interaction required a513LXCXE517motif on the RBAP96 protein and an intact A/B binding pocket of RB.RBAP96 enhances RB-mediated transcriptional repression.Finally,enforced expression of RBAP96 caused an elevated radiosensitivity of human breast cancer cells bearing wtRB,but did not affect radiosensitivity of breast cancer cells bearing mutant RB.Expression of a full-length RBAP96 with an513LXCXE517inactivating mutation(LXCXE→RXRXH)failed to result in any radiosensitivity alteration. Conclusion This study for the first time characterizes a novel RB-interacting protein RBAP96 and demonstrates that enforced expression of RBAP96 causes an increase of RBAP96-mediated transcription activation and radiosensitivity via a RB-interacting dependent manner.
RBAP96;Retinoblasloma protein;Breast neoplasms;Transcription repression; Radiation tolerance
Fund programs:National Natural Science Foundation of China(81071906,81172127,81572969, 81402633);Technology and Development and Research Projects for Research Institutes,Ministry of Science and Technology(2014EG150134);Tianjin Science&Technology Pillar Program(14ZCZDSY00001);Natural Science Foundation of Tianjin(16JCQNJC13600);Peking Union Medical College Youth Innovation Fund(1581);IRM-CAMS Research Fund(1614)
The retinoblastoma susceptibility gene encodes a tumor suppressor protein(called"RB").Inactivation of which contributes to a broad range of tumors,including breast cancer[1].The RB protein plays major roles in regulation of cell proliferation,cell cycle progression, cell fate,telomerase activity,and radiosensitivity.Nevertheless,the majority of human breast cancers does not contain a defective(mutated)or otherwise inactivated RB gene[2].Thus,breast cancers have other mechanisms of overcoming the function of the RB protein, such as transcription repression and radiosensitivity, but these mechanisms are mostly unknown[3].
In the current study,using a yeast two-hybrid screen of a human breast cDNA library,we isolated a novel RB-interacting protein that we named RBAP96 (for RB-Associated Protein of molecular size 96 kilodaltons),and demonstrated that the513LXCXE517motif embedded inside the RBAP96 protein was necessaryfor binding of RBAP96 to RB,an intact A/B binding pocket of RB was also required for the binding.Furthermore,this motif was required for the functional consequences of the BRCA1:RB interaction,including RB-mediated transcriptional activation(E2F1 and Cyclin A)and RB-dependent radiosensitivity.
1 MATERIALS AND METHODS
1.1 Cell culture
Human breast cancer MCF-7 and BT549 cells, and human osteoblast-like SAOS-2 cells were obtained from the Georgetown University(Washington DC,USA),and maintained in Dulbecco′s modified Eagle′s medium(DMEM)(Gibco-BRL,USA)supplemented with 5%(MCF-7 and BT549)or 10%(SAOS-2) fetal bovine serum,non-essential amino acids(5 mmol/L), penicillin(100 U/mL),and streptomycin(100 μg/mL) in an incubator at 37℃with 5%CO2.
1.2 Expression vectors and transfection.
The construction of p3XFLAG-RBAP96 expression plasmid was performed as the protocol described in our previous studies[4].For transient transfection assays,subconfluent proliferating cells were incubated overnight with 10 μg of plasmid DNA per 100 mm dish,in the presence of Lipofectamine 2000,and then washed to remove the Lipofectamine 2000 and the excess plasmid DNA.
1.3 Irradiation
The transfected and untransfected cells were irradiated with various doses of 0,2,4,6,8,and 10 Gy rays using a137Cs irradiator at a dose rate of 1 Gy/min at room temperature.
1.4 Colony formation assay
Cells irradiated were plated in 60-mm culture dishes at a cell density of 2×103.After 3 weeks postirradiation,cells were fixed and stained with Giemsa solution.Colonies containing>50 cells were counted. Colony forming efficiency=number of colonies formed/ number of cells plated.
1.5 Western blot analysis
Western blot analysis was carried out as described before[5].Briefly,after cells were harvested and lysed,equal amounts of protein were separated using 10%SDS-PAGE gel electrophoresis,transferred onto nitrocellulosemembranesand blocked with 5%skimmed milk.The membranes were blotted with primary antibodies against the FLAG epitope(M2,mouse monoclonalantibody,Sigma,1:500 dilution),RB(C-20,rabbit polyclonal,Santa Cruz Biotechnology,1:200),and α-actin(I-19,goat polyclonal,Santa Cruz Biotechnology,USA,1:2000),and then incubated with HRP-conjugated anti-goat or anti-rabbit IgG antibodies(SantaCruz Biotechnology,Santa Cruz,CA,USA,1:2000). Protein bands were visualized using the enhanced chemiluminescence detection system(Amersham Life Sciences,USA).Equality of protein loading was confirmed by fast green staining of the membrane and by immunoblotting for α-actin.Colored markers(BioRad Corp.,USA)were used as molecular size standards.
1.6 Immunoprecipitation(IP)
Subconfluent proliferating cells were harvested, and nuclear extracts were prepared,as described earlier[4].Each IP was carried out using 6 μg of antibody and 1000 μg of nuclear extract protein.Precipitated proteins were collected using protein G beads, washed,eluted in boiling Laemmli sample buffer,and subjected to Western blot assay.The IP antibodies used were:RBAP96 IP(anti-FLAG M2 antibody,Sigma,USA)and RB IP (M-153,Santa Cruz,USA);and the western blot antibodies were:RBAP96[N20,a rabbit polyclonal IgG against recombinant GST-RBAP96 (aa 25-105)];and RB(C-15,Santa Cruz,USA).
1.7 Assay of transcriptional activity
Transcriptional activity assay was described in our previous studies[4].Briefly,sub-confluent proliferating cells in 24-well plastic dishes were incubated overnight with 0.5 μg of each vector in serum-free DMEM containing LipofectAMINE 2000(Life Technologies,Inc.).The total transfected DNA was kept constant by addition of an appropriate amount of the control vector.To control transfection efficiency,plasmid pRSV-β-gal was cotransfected to allow normalization of luciferase values to β-galactosidase activity in the same sample.The transfected cells were harvestedfor luciferase assays for 24 h.Luciferase activities were expressed relative to the un-transfected controls. Each experimental condition was tested in four replicate wells;and the experiments shown are representative of at least three independent experiments.
1.8 Glutathione-S-transferase(GST)capture assays
GST bead assays were performed essentially as described previously[5].[35S]methionine-labeled proteins were prepared by in vitro transcription(using the T7 promoter of pcDNA3)and translation.The GST fusion proteins were generated from cDNAs cloned into the p-GEX vector,expressed in Escherichia coli,and purified by affinity chromatography.Labeled proteins were incubated with GST(control)or GST fusion proteins for 24 h at 4℃,recovered using glutathione agarose beads,eluted in boiling sample buffer,and analyzed by SDS-PAGE autoradiography.GST fusion proteins were visualized by Western blot assay,using an anti-GST mouse monoclonal(B-14,1:10 000;Santa Cruz Biotechnology,USA).
1.9 Statistical methods
Where appropriate,statistical comparisons were made using the two-tailed Student′s t test with the statistical software SPSS 13.0(SPSS Inc.,Chicago,IL, USA).A P value of less than 0.05 was considered statistically significant.The data are shown as the mean± standard error of the mean(SEM).
2 RESULTS
2.1 Isolation of RBAP96
We conducted a yeast two-hybrid screen using the standard A/B pocket of RB [amino acid 395-876]as bait.After screening of a human breast cDNA library(Clontech,USA),we isolated and sequenced a single positive 2 564-bp clone that encodes a predicted protein of 854 amino acid,encoded by a gene of unknown function(Protein ID BAA91666)(Fig.1).In vitro translation(IVT)gave a product close to the expected size of 96 000,which we named RB Associated Protein of 96 000(RBAP96).Structural features of RBAP96 include an LXCXE site(513LDCGE517),an essential role for the LXCXE(leucine-X-cysteine-X-glutamic;X=any amino acid)motif.
2.2 RBAP96 binds to RB via LXCXE motif
We showed an in vivo association of RBAP96 with RB by IP-Western blot(Fig.2A).RB was found in the RBAP96 IP of MCF-7 breast cancer cells containing wild-type(wt)RB,but not in the RBAP96 IPs of BT549 bearing an Rb deletion).Neither RB nor RBAP96 was found in the control IP(normal IgG)of MCF-7 cells.We also showed direct binding of RBAP96 to RB by GST capture assays.IVT wtRBAP96 bound to beads coated with GST-RB but not to beads coated with GST alone.IVT-RBAP96/RXRXH, in which513LXCXE was converted to RXRXH,failed to bind to GST-RB(Fig.2B).IVT-wtRBAP96 failed to bind to GST-RB-N757F,which contains a mutation of the A/B pocket of RB,suggesting that an intact A/B pocket of RB is required to bind RBAP96.

Fig.1 RBAP96 protein sequence.A LXCXE motif(amino acid 513-517)is indicated as bold.
2.3 RBAP96 enhances RB-mediated transcriptional repression
RB regulation of cell cycle progression depends upon the repression of E2F1-regulated transcription[4]. We tested the effect of RBAP96 on the RB-regulatedactivity of an E2F-responsive reporter.Exogenous RB inhibited the E2F1-induced activation of the E2F reporter in Rb-null SAOS-2 cells,and this inhibition was enhanced by co-expression of wtRBAP96(t=1.987, P<0.001),but not by RBAP96/RXRXH or empty vector(Fig.3),wtRBAP96 did not inhibit E2F1 activity without RB.wtRBAP96 also enhanced RB-mediated repression of the cyclin A promoter(another target of RB)(t=1.489,P<0.01),suggesting that RBAP96 acts as a corepressor for RB transcriptional repression,at least E2F1 and cyclin A.

Fig.2 RBAP96 and RB interaction.A.In vivo association.MCF-7 or BT549 cells were transfected with a FLAG-wtRBAP96 expression vector; and IP-WB was performed,as described before[5].A normal mouse IgG IP from MCF-7 cells was used as negative control.The IP antibodies used were:RBAP96 IP(anti-FLAG M2 antibody,Sigma)and RB IP(M-153,Santa Cruz);and the westen blot antibodies were:RBAP96[N20, a rabbit polyclonal IgG against recombinant GST-RBAP96(aa 25-105)];and RB(C-15,Santa Cruz).The data are representative of three experiments.B.In vitro binding.IVT35S-labeled wtRBAP96 or RBAP96-RXRXH proteins were prepared using the T7 promoter of p3XFALG vector.GST-RB and GST-RB-N757 vectors were described before[5].The input lanes show 10%of the IVT product used for capture.
2.4 RBAP96 causes an increase of RB-dependent radiosensitivity
Finally,we determined any alteration of radiosensitivity in breast cancer cells with wild-type RB(MCF-7)vs.mutant(deleted)RB(BT549)by transfection of p3XFLAG-CMV,p3XFLAG-wtRBAP96 or p3XFLAGRBAP96/RXRXH.As illustrated in Fig.4,wtRBAP96 expression significantly increased MCF-7 cell susceptibility to γ-ray irradiation,but did not exhibit any effect in BT549 cells irradiated at the same condition.For example,IC90,a dose which inhibits 90%of cell survival,was 1.7 Gy(t=1.320,P<0.05)in wtRBAP96 transfected cells compared to p3XFLAG-CMV transfected control cells(3.7 Gy).On other hand,RBAP96/ RXRXH expression failed to cause any change of radiosensitivity.These results indicate that regulation of radiosensitivity by RBAP96 requires the interaction ofRBAP96 with RB.
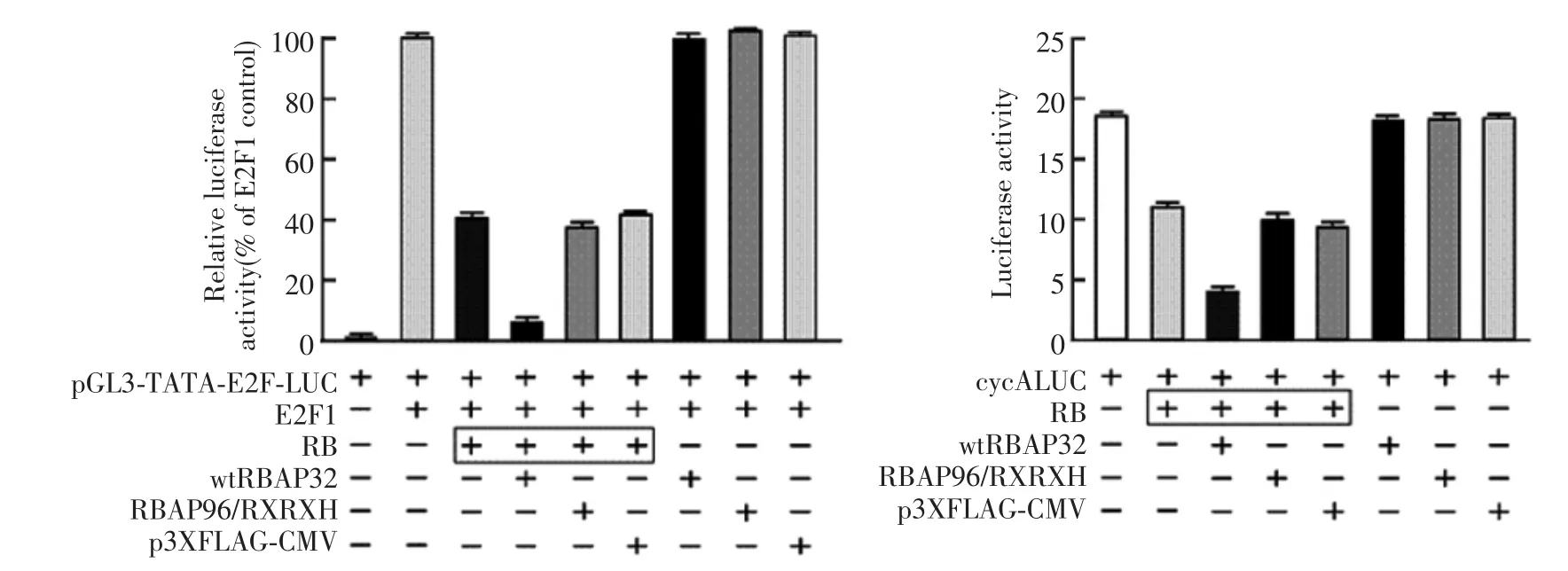
Fig.3 RBAP96 enhances RB-repression of E2F-responsive promoters.Exponentially growing SAOS-2 cells in 2 cm2well dishes were transfected overnight with 0.25 μg of each vector per well using Lipofectamine 2000 and post-incubated for 24 h.The total transfected DNA was kept constant by adding control vector.The pRSV-β-gal plasmid was used to monitor transfection efficiency.Luciferase values are means±SEMs from three independent experiments,each using 10 replicate wells per condition.The E2F reporter consists of an E2F binding site upstream of a TATA box and the luciferase gene;the cyclin A reporter contains nucleotides-608 to+97 of the human cyclin A promoter upstream of luciferase.
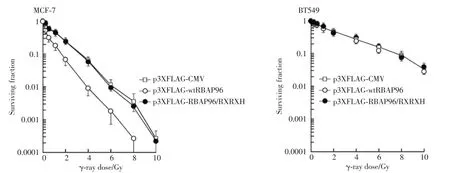
Fig.4 RBAP96 enhances radiosensitivity in MCF-7 cells,not in BT549 cells.The MCF-7 and BT549 cells were transfected with the p3XFLAG-wtRBAP96 or p3XFLAG-RBAP96/RXRXH expression plasmid as described in the“Materials and Methods”,irradiated at indicated doses 24 h following transfection,and subjected to clonogenic survival assay.
3 DISCUSSION
Previous studies suggest that cell cycle regulatory proteins(G1/S cyclins and CDKs)and several DNA tumor viruses express proteins(E1a from adenovirus,E7 from HPV,and T antigen from SV40)affect RB functions,including cell growth,cell cycle and radiosensitivity,by interacting with RB via a LXCXE binding site[1-3].Moreover,LXCXE sequence disruption in these proteins results in a loss of their effects on RB and their ability to transform cells,indicating the importance of the LXCXE binding site[6-10].In the current study,we isolated a novelRB binding protein RBAP96 by a yeast two-hybrid screen using the standard A/B pocket of RB as bait.As well-known,RB contains at least four distinct protein-binding domains, including the large A/B pocket(amino acid 395-876), corresponding to the binding site for E2F,the A/B pocket,the C pocket,and the N-terminal domain.Our results with IP-Western blotting and GST capture assay revealed that RBAP96 associated with RB,the LXCXE motif embedded inside the RBAP96 protein (amino acid 513-517)and the A/B pocket of RB were absolutely required for RBAP96 binding to RB,suggesting that RBAP96 is a LXCXE-dependent RB interacting protein.
RB encoding a tumor suppressor phosphoprotein is often lost or functionally inactivated in many types of tumors,and plays an important role in the regulation of cell proliferation,cell cycle progression,apoptosis,and telomerase activity by interacting with transcription factors and altering the activity of transcription factors[1-3,11].One target of RB is the E2F family of cell cycle transcription factors,the binding of which by RB blocks E2F-and cyclin A-mediated transcriptional activation[5,12-13].We found RBAP96 enhanced RB repression of E2F1 transcription activity.Not only E2F1,RB-mediated repression of cyclin A promoter transcription activity was also reinforced by RBAP96. As expected,mutation of LXCXE motif failed to produce any impact on RB transcriptional repression. These findings suggest that RBAP96 not only interacts with RB,but also affect RB regulation of transcription activity,and the LXCXE motif is also essential.
RB is an important target in cancer radiotherapy, including breast cancer[14-15].Our further studies demonstrated that RBAP96 significantly increased human breast cancer cell susceptibility to γ-ray irradiation via a RB-interacting dependent manner,since such increased radiosensitivity was only seen in MCF-7 cells containing intact RB function,but in BT-549 bearing mutant(deleted)RB.Moreover,RBAP96 with a mutant LXCXE motif failed to influence radiosensitivity in both MCF-7 and BT-549 cell lines.These results make clear that the LXCXE motif is not only essential for the RBAP96:RB interaction,but also necessary for RBAP96-mediated regulation of RBtranscription repression and radiosensitivity.A followup study is being carried out to determine whether RBAP96 affects DNA damage and repair pathway via interacting with other as-yet-undefined cellular regulators,except binding to RB.Additionally,our previous studies found that HMGB1-4,other new RB binding proteins by a LXCXE motif,affected radiosensitivity via a RB-dependent or independent manner[16-17]. Together,these findings that the interaction with RB is necessary in mediating radiosensitivty for some of RB associated proteins,such as RBAP96,but not criticalfor otherRB-associated proteins,including HMGB1[16-17].It will be very interesting to explore what mechanism(s)involved in such difference.
Conflict of interestThe authors declare no conflicts of interest.
Authors contribution statementCell culture,survival and Western blot assay were carried out by Junling Zhang,Xiaolei Xue,Lu Lu,Ming Cui.Construction of expression vectors and IP-Western blot were conducted by Qinghui Meng.Junling Zhang and Saijun Fan were in change of experiment design and writing.
REFERENCES
[1]Dyson NJ.RB1:a prototype tumor suppressor and an enigma[J]. Genes Dev,2016,30(13):1492-1502.DOI:10.1101/gad.282145.
[2]Witkiewicz AK,Knudsen ES.Retinoblastoma tumor suppressor pathway in breast cancer:prognosis,precision medicine,and therapeutic interventions[J].Breast Cancer Res,2014,16(3):207.DOI: 10.1186/bcr3652.
[3]Johnson J,Thijssen B,Mcdermott U,et al.Targeting the RB-E2F pathway in breast cancer[J].Oncogene,2016,35(37):4829-4835. DOI:10.1038/onc.2016.32.
[4]Fan SJ,Yuan RQ,Ma YX,et al.Disruption of BRCA1 LXCXE motif alters BRCA1 functional activity and regulation of RB family but not RB protein binding[J].Oncogene,2001,20(35):4827-4841. DOI:10.1038/sj.onc.1204666.
[5]Fan SJ,Ma YX,Wang CG,et al.p300 modulates the BRCA1 inhibition of estrogen receptor activity[J].Cancer Res,2002,62(1):141-151.
[6]Decaprio JA.How the Rb tumor suppressor structure and function was revealed by the study of Adenovirus and SV40[J].Virology, 2009,384(2):274-284.DOI:10.1016/j.virol.2008.12.010.
[7]Iwahori S,Hakki M,Chou S,et al.Molecular determinants for the inactivation of the retinoblastoma tumor suppressor by the viral cyclin-dependent kinase UL97[J].J Biol Chem,2015,290(32):19666-19680.DOI:10.1074/jbc.M115.660043.
[8]Wang Y,Zheng Z,Zhang J,et al.A novel retinoblastoma protein (RB)E3 ubiquitin ligase(NRBE3)promotes RB degradation and is transcriptionally regulated by E2F1 transcription factor[J].J Biol Chem,2015,290(47):28200-28213.DOI:10.1074/jbc.M115.655597.
[9]Nor Rashid N,Yong ZL,Yusof R,et al.HPV 16E7 and 48E7 proteins use different mechanisms to target p130 to overcome cell cycle block[J/OL].Virol J,2016,13:2[2016-08-17].http://virologyj.biomedcentral.com/articles/10.1186/s12985-015-0460-8.DOI: 10.1186/s12985-015-0460-8.
[10]Chan HM,Smith L,La Thangue NB.Role of LXCXE motif-dependent interactions in the activity of the retinoblastoma protein[J]. Oncogene,2001,20(43):6152-6163.
[11]Vormer TL,Wojciechowicz K,Dekker M,et al.RB family tumor suppressor activity may not relate to active silencing of E2F target genes[J].Cancer Res,2014,74(18):5266-5276.DOI:10.1158/ 0008-5472.CAN-13-3706.
[12]Kawatsuki A,Yasunaga JI,Mitobe Y,et al.HTLV-1 bZIP factor protein targets the Rb/E2F-1 pathway to promote proliferation and apoptosis of primary CD4(+)T cells[J].Oncogene,2016,35(34): 4509-4517.DOI:10.1038/onc.2015.510.
[13]Sharon E,Volchek L,Frenkel N.Human herpesvirus 6(HHV-6) alters E2F1/Rb pathways and utilizes the E2F1 transcription factor to Express viral genes[J].Proc Natl Acad Sci USA,2014,111(1): 451-456.DOI:10.1073/pnas.1308854110.
[14]Ma W,Yu J,Qi X,et al.Radiation-induced microRNA-622 causes radioresistance in colorectal cancer cells by down-regulating Rb[J]. Oncotarget,2015,6(18):15984-15994.DOI:10.18632/oncotarget. 3762.
[15]Mao A,Liu Y,Wang Y,et al.miR-449a enhances radiosensitivity through modulating pRb/E2F1 in prostate cancer cells[J].Tumour Biol,2016,37(4):4831-4840.DOI:10.1007/s13277-015-4336-8.
[16]Jiao Y,Wang HC,Fan SJ.Growth suppression and radiosensitivity increase by HMGB1 in breast cancer[J].Acta Pharmacol Sin,2007, 28(12):1957-1967.DOI:10.1111/j.1745-7254.2007.00669.x.
[17]Wang LL,Meng QH,Jiao Y,et al.High-mobility group boxes mediate cell proliferation and radiosensitivity via retinoblastoma-interaction-dependent and-independent mechanisms[J].Cancer Biother Radiopharm,2012,27(5):329-335.DOI:10.1089/cbr.2012.1199.
(Received by 2016-08-18)
Fan SJ,Email:fansaijun@irm-cams.ac.cn
10.3760/cma.j.issn.1673-4114.2016.05.001
