丹参酚酸B和山楂黄酮合用对游离脂肪酸诱导的大鼠肝细胞脂质沉积和凋亡的作用
2016-05-04薛冬英袭渤人张洁叶军
薛冬英 袭渤人 张洁 叶军
200062 上海中医药大学附属普陀医院感染科
·论著·
丹参酚酸B和山楂黄酮合用对游离脂肪酸诱导的大鼠肝细胞脂质沉积和凋亡的作用
薛冬英袭渤人张洁叶军
200062上海中医药大学附属普陀医院感染科
【摘要】目的研究丹参酚酸B和山楂黄酮对游离脂肪酸诱导大鼠肝细胞脂质沉积和凋亡的作用及其机制。方法采用肝脏原位胶原酶灌注法分离大鼠原代肝细胞,油酸:棕榈酸=2:1造模。原代肝细胞和HepG2均分为正常对照组、模型组、模型+丹参酚酸B组、模型+山楂黄酮组、模型+丹参酚酸B+山楂黄酮组、模型+SP600125组、丹参酚酸B组、山楂黄酮组、丹参酚酸B+山楂黄酮组、SP600125组、DMSO对照组。游离脂肪酸刺激原代肝细胞的浓度为250 μmol/L、刺激HepG2的浓度为500 μmol/L;丹参酚酸B的浓度为1 μmol/L;山楂黄酮的浓度为5 μg/mL;SP600125的浓度为10 μmol/L。观察①油红O染色:丹参酚酸B和山楂黄酮对游离脂肪酸刺激的肝细胞内脂滴变化的影响;②ELISA法:丹参酚酸B和山楂黄酮对游离脂肪酸诱导的肝细胞凋亡的影响;③高内涵筛选Hoechst33258染色:丹参酚酸B和山楂黄酮对游离脂肪酸诱导的肝细胞凋亡的影响;④Western blot:丹参酚酸B和山楂黄酮对游离脂肪酸刺激的肝细胞内p-JNK表达的影响。结果①油红O染色显示:与正常对照组比较,游离脂肪酸作用于原代肝细胞、HepG2细胞24 h后,细胞内脂滴含量显著增加(P均<0.01)。丹参酚酸和山楂黄酮可显著减少游离脂肪酸诱导的原代肝细胞、HepG2细胞内脂滴的含量(P<0.01,P<0.05)。②ELISA检测结果:与正常对照组相比游离脂肪酸组原代肝细胞及HepG2细胞吸光值[Absorbance(A405 nm-A490nm)]显著增高,细胞凋亡明显(P均<0.01);丹参酚酸B和山楂黄酮显著抑制游离脂肪酸诱导的原代肝细胞、HepG2细胞凋亡(P均<0.01)。③ 高内涵筛选Hoechst33258染色:与正常对照组比较,游离脂肪酸组细胞核平均荧光强度值及凋亡率明显增高(P均<0.01)。丹参酚酸B和山楂黄酮显著降低游离脂肪酸刺激的原代肝细胞细胞核平均荧光强度值及凋亡率(P均<0.01)。HepG2细胞检测结果与原代肝细胞趋势一致。④Western blot结果:与正常对照组比较,游离脂肪酸刺激原代肝细胞24 h时可显著促进p-JNK的表达(P均<0.01)。丹参酚酸B和山楂黄酮对游离脂肪酸刺激原代肝细胞24 h时p-JNK的表达有显著抑制作用(P均<0.01)。HepG2细胞p-JNK结果与原代肝细胞趋势一致。结论①丹参酚酸B和山楂黄酮合用能够抑制游离脂肪酸诱导的肝细胞内脂滴增加及肝细胞凋亡。②丹参酚酸B和山楂黄酮合用能够抑制游离脂肪酸诱导的肝细胞内JNK的活化。二者是丹参酚酸B和山楂黄酮合用防治非酒精性脂肪性肝炎的重要机制。
【关键词】肝细胞;丹参酚酸B;山楂黄酮;游离脂肪酸;肝细胞凋亡;c-jun氨基末端激酶
非酒精性脂肪性肝炎(nonalcoholic steatohepatitis,NASH)以肝细胞的损伤、脂肪贮积、凋亡和肝脏炎症、纤维化为特征,可进展至肝硬化、肝功能衰竭甚至肝细胞肝癌等[1],目前没有特效药治疗,本研究探讨丹参酚酸B与山楂黄酮合用对游离脂肪酸诱导的大鼠肝细胞内脂质沉积、肝细胞凋亡及其相关JNK信号蛋白的作用。
资料和方法
一、材料
(一) 实验动物Sprague Dawley雄性大鼠,体质量120~180 g,清洁级,购自上海西普尔-必凯实验动物有限公司,饲养于上海中医药大学实验动物中心。
(二) 实验细胞采用人肝癌细胞株(human hepatoblastoma cell line,HepG2),购自中国科学院上海生命科学研究院细胞库。
(三) 试剂丹参酚酸B(Salvianolic acid B,Sal B):Salvianolic acid B-Mg2+(Sal B-Mg2+)购自上海同田生物技术股份有限公司,纯度≥95%(by HPLC),分子式(C36H30O16Mg),分子量为742;山楂黄酮(Hawthorn flavone)购自南京泽郎医药科技有限公司,纯度≥95%(by HPLC)。油酸、棕榈酸、DMSO、胶原酶Ⅳ型、油红O购自美国Sigma公司;JNK Inhibitor II(SP600125);MEM培养基干粉、DMEM培养基干粉、RPMI1640培养基干粉;胎牛血清、胰蛋白酶购自美国Gibco公司。Phospho-SAPK/JNK(Thr183/Tyr185)(98F2) Rabbit mAb购自美国Cell Signaling Technology公司产品。PageRulerTMPrestained Protein Ladder Plus(蛋白预染marker)购自美国MBI Fermentas公司。GAPDH抗体、KC-5G4购自上海康成生物工程有限公司。IRDye 680 goat anti-rabbit IgG、IRDye 800CW donkey anti-mouse IgG、Odyssey Blocking Buffer购自美国LI-COR公司。PVDF膜(0.45 μm)购自美国Millipore公司。Western及IP细胞裂解液、Hoechst 33258染色液、BCA蛋白浓度测定试剂盒、台盼蓝染色细胞存活率检测试剂盒、SDS-PAGE蛋白上样缓冲液(5×)购自碧云天生物技术研究所。蛋白酶抑制剂混合物(complete mini)、磷酸酶抑制剂混合片(PhosSTOP)、Cell Death Detection ELISAPLUS均为德国Roche公司产品。HEPES、Tris、EDTA、TEMED、Glycine、SDS购自美国Amresco公司。
(四)设备高温灭菌CO2培养箱,371型,美国Thermo Fisher Scientific公司。生物安全柜,HR60-IIA2,青岛海尔特种电器有限公司。微孔板扫描分光光度计,MQX200R,美国Bio-Tek公司。多功能电子蠕动泵,DDB-320,宁波石浦海天电子仪器厂,留置针,18G,上海卫生技术设备公司。双色红外激光成像系统,Odyssey;电泳仪,PowerPac Basic;半干转膜仪,1703940型,均为美国BIO-RAD公司生产。冷冻离心机,Jouan MR23i型;高内涵筛选分析仪,ArrayScan○RVTI;酶标仪,Multiskan MK3,为美国Thermo Fisher Scientific公司生产。
二、方法
(一)肝细胞分离使用3%戊巴比妥钠(0.2 mL/100 g)腹腔注射麻醉大鼠,将大鼠固定在解剖板上。75%酒精消毒胸腹部皮毛,而后U型剖腹,18G留置针穿刺门静脉,结扎线固定穿刺针;待血液流出后,接通静脉输液管,开启恒流泵(流量为10 mL/min),37 ℃温育的前灌流液冲洗肝脏,立即剪断下腔静脉。接着以37 ℃预温育的Ⅳ型胶原酶液(5 mL/min)灌流,最后摘下肝脏,置于盛有4 ℃预冷的MEM培养液的灭菌100 mm培养皿中。分散后的细胞悬液经双层纱布过滤并分装入灭菌的50 mL聚丙烯离心管内,经4 ℃离心3次。
(二)肝细胞培养根据细胞计数,调整细胞浓度至5×105cells/mL接种。置于37 ℃、5% CO2~95%空气的CO2培养箱中培养,约4~6 h细胞贴壁后换液。实验前改为0.5%FBS-RPMI1640培养。
(三)HepG2细胞株培养以0.25%胰酶/0.02%EDTA消化传代,以含10%FBS-DMEM培养液,置于37 ℃、含5% CO2~95%空气的CO2培养箱中培养。待细胞融合度至70%~80%,换液,实验前改为0.5%FBS-DMEM培养。
(四)分组原代肝细胞和HepG2细胞株均分为:正常对照组、模型组、模型+丹参酚酸B组、模型+山楂黄酮组、模型+丹参酚酸B+山楂黄酮、模型+SP600125组、丹参酚酸B组、山楂黄酮组、丹参酚酸B+山楂黄酮组、SP600125组、DMSO对照组。根据参考文献[2,3],以250 μmol/L(油酸∶棕榈酸=2∶1)为原代肝细胞非酒精性脂肪性肝炎体外模型的刺激浓度,500 μmol/L(油酸∶棕榈酸=2∶1)为HepG2非酒精性脂肪性肝炎体外模型的刺激浓度。根据参考文献常用剂量[4-7]SP600125浓度为10 μmol/L,丹参酚酸B浓度为1 μmol/L,山楂黄酮5 μg/mL。
(五)脂滴油红O染色将原代肝细胞及HepG2以适当密度接种于96孔培养板,每组设5个复孔,100 μL培养液/孔,加药培养24 h。用PBS洗3次,100 μL 50%异丙醇/孔固定1 min。油红O新鲜染液100 μL/孔染色15 min,60%异丙醇100 μL/孔洗去多余染液,三蒸水100 μL/孔冲洗3次。每孔加100 μL DMSO溶解油红O,振荡混匀,酶标仪510 nm波长处测定光密度(OD)值。
(六)ELISA法检测细胞凋亡各组细胞加药培养24 h后,以PBS洗涤3次,每皿(60 mm)加入200 μL裂解液(Lysis B?ffer),将培养皿中的细胞刮下,收入1.5 mL离心管中,混匀,15~25 ℃作用30 min,然后200 g离心10 min,收集细胞裂解物按照分组将96孔微孔板中每孔加入20 μL样本,同时加入80 μL免疫反应试剂(抗组蛋白-生物素∶抗-DNA-POD∶温育缓冲液=1∶1∶18),封口,置摇床上,300 rpm,15~25 ℃,孵育2 h。弃上清,用250 μL温育缓冲液/孔(Incubation Buffer)洗涤3次,加入100 μL ABTS底物缓冲液/孔,置摇床上,250 rpm,15~25 ℃,孵育至颜色变化至适合程度。以DNA-组蛋白复合物作阳性对照,底物缓冲液作空白对照,15 min后作比色分析,检测波长405 nm,内参波长490 nm。以吸光值[Absorbance(A405nm-A490nm)]进行统计分析。
(七)高内涵筛选Hoechst33258染色检测细胞凋亡按分组将细胞以每孔100 μL接种于96孔培养板,每组设3个复孔,加药培养24 h后,以温育的PBS洗涤三遍,每孔加入Hoechst33258染色液100 μL,置含5% CO2-95%空气的37 ℃ CO2培养箱中培养30 min。弃染色液,用PBS 100 μL/孔洗涤2次,使用高内涵筛选分析仪进行检测。
(八)Western-blot检测细胞内p-JNK蛋白表达用预冷的PBS洗涤细胞两次,加入含蛋白酶磷酸酶抑制剂的细胞裂解液。收集细胞至EP管中,混匀,冰上静置30 min。4 ℃,12 000 rpm离心15 min,取上清。按照BCA蛋白浓度测定试剂盒进行蛋白定量,行SDS-PAGE凝胶电泳。电转移至PVDF膜,Western 印迹检测p-JNK(p-JNK工作液浓度为1∶1000),二抗为相应来源的以Odyssey封阻液稀释的红外染料标记二抗(1∶5 000稀释)。GAPDH的检测:一抗为GAPDH多克隆抗体,二抗为相应来源的以Odyssey封阻液稀释的红外染料标记二抗(1∶5 000稀释)。选择合适的扫描区域、分辨率、荧光强度,设置700通道(680 nm激发,720 nm检测)或800通道(780 nm激发,820 nm检测),进行扫描。
三、统计学处理
计量资料用均数±标准差表示。多组间比较采用SPSS 20.0统计分析软件进行单因素方差分析(One-Way ANOVA)。两两之间多重比较,方差齐者用LSD检验,方差不齐者用Dunnett's T3检验。
结果
一、油红O染色检测丹参酚酸B和山楂黄酮对游离脂肪酸刺激的细胞内脂滴变化的影响(见表1)
与正常对照组比较,游离脂肪酸作用于原代肝细胞、HepG2细胞24 h后,细胞内脂滴含量显著增加(P均<0.01)。丹参酚酸B、山楂黄酮、丹参酚酸B +山楂黄酮、SP600125对正常培养的原代肝细胞、HepG2细胞内脂滴含量无明显影响,但显著减少游离脂肪酸诱导的原代肝细胞、HepG2细胞内脂滴含量(P<0.01,P<0.05)。
二、 ELISA法检测丹参酚酸B和山楂黄酮对游离脂肪酸诱导的细胞凋亡的影响(见表2)
与正常对照组相比,游离脂肪酸组原代肝细胞、HepG2细胞吸光值[Absorbance(A405nm-A490nm)]显著增高,细胞凋亡明显(P均<0.01);丹参酚酸B、山楂黄酮、丹参酚酸B +山楂黄酮、SP600125对正常培养的原代肝细胞、HepG2吸光值无明显影响,但显著减少游离脂肪酸诱导的原代肝细胞、HepG2细胞吸光值(P<0.01,P<0.05)。
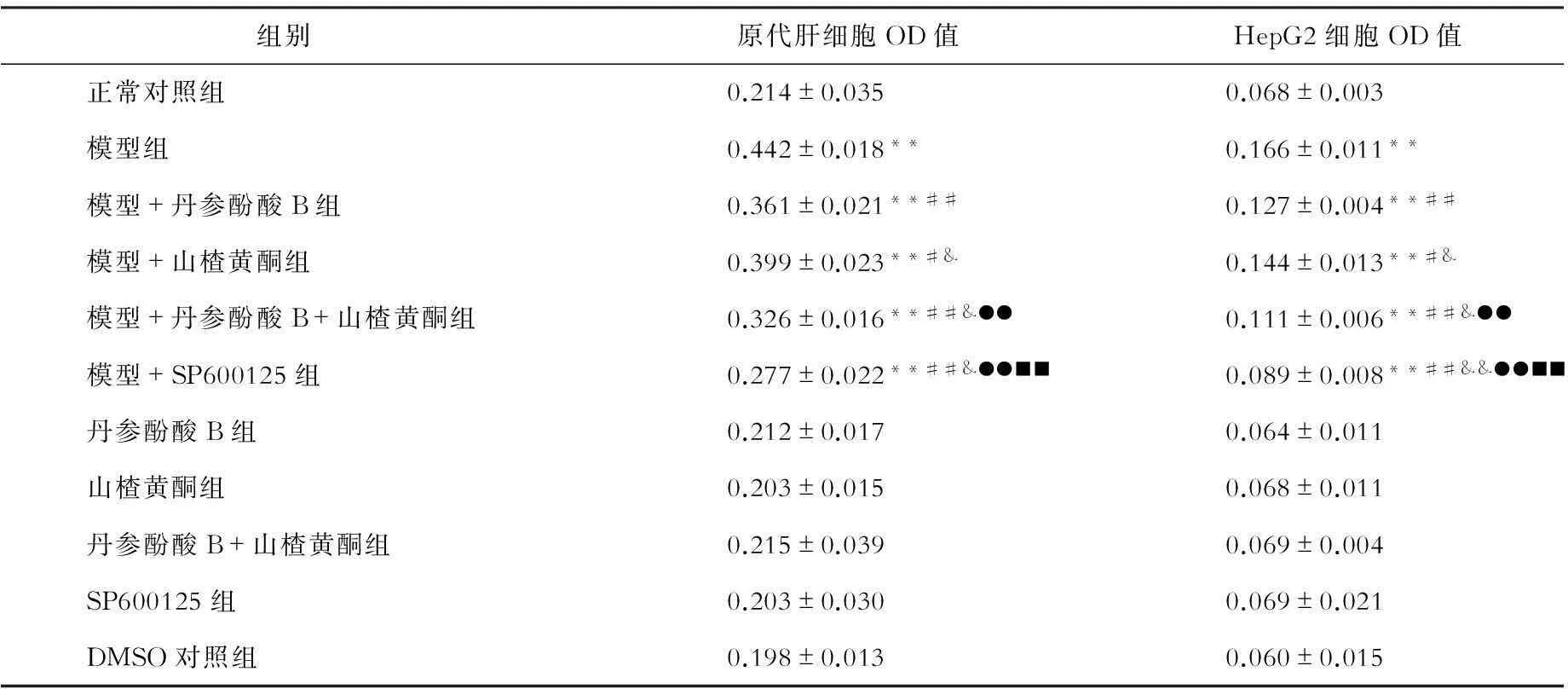
表1 各组原代肝细胞、HepG2细胞油红O染色光密度(OD)值比较
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
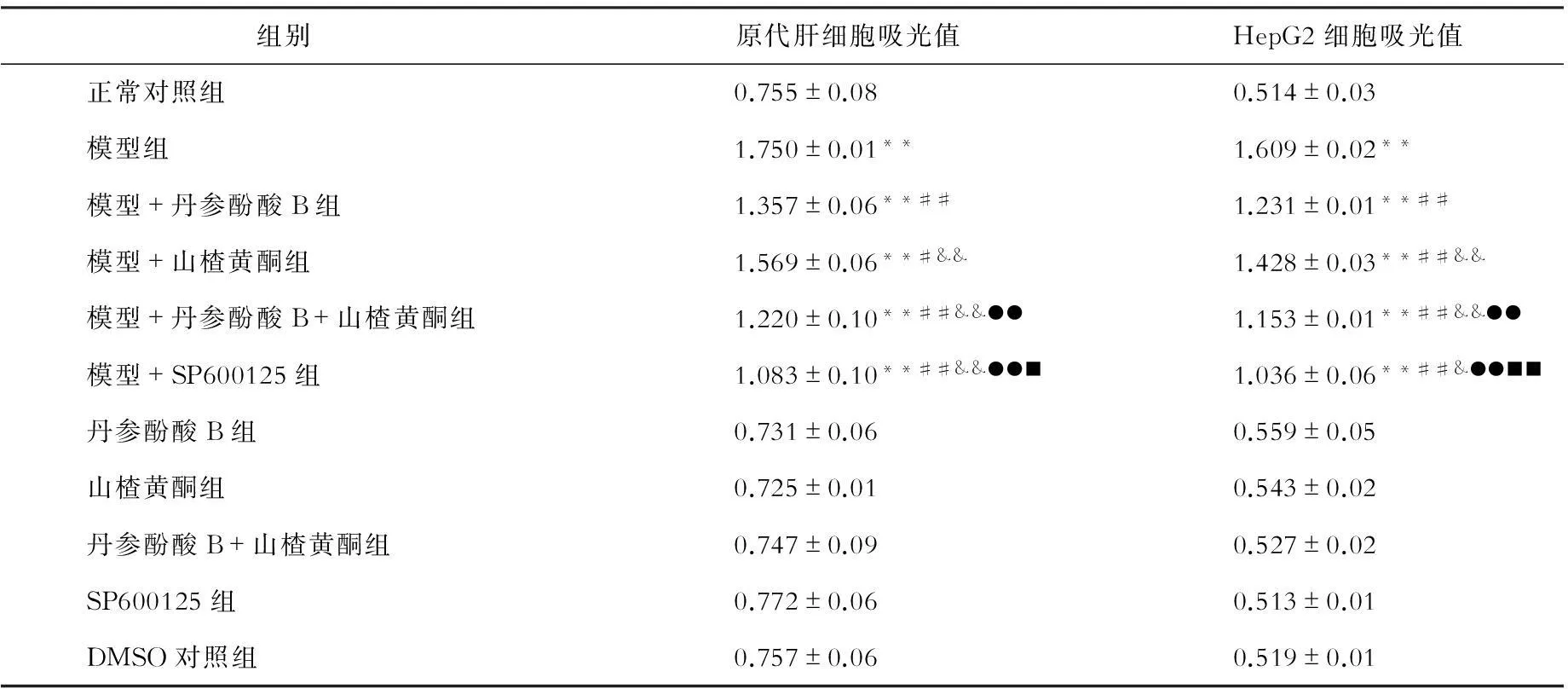
表2 各组原代肝细胞、HepG2细胞吸光值比较
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
三、高内涵筛选Hoechst33258染色分析丹参酚酸B和山楂黄酮对游离脂肪酸诱导的细胞凋亡的影响
与正常对照组比较,游离脂肪酸组原代肝细胞核平均荧光强度值及凋亡率明显增高(P均<0.01)。丹参酚酸B、山楂黄酮、丹参酚酸B和山楂黄酮、SP600125显著降低游离脂肪酸刺激的原代肝细胞细胞核平均荧光强度值及凋亡率(P<0.05,P<0.01)。见表3。
高内涵筛选Hoechst33258染色观察HepG2细胞凋亡(见表4):各组细胞核平均荧光强度值及凋亡率比较,趋势与原代肝细胞一致。
四、Western blot法观察丹参酚酸B和山楂黄酮对游离脂肪酸刺激的细胞内p-JNK表达的影响药物作用24 h,与正常对照组比较,游离脂肪酸可显著促进p-JNK的表达(P均<0.01)。丹参酚酸B、山楂黄酮、丹参酚酸B和山楂黄酮、SP600125对正常培养的24 h原代肝细胞p-JNK表达无明显影响,但对经游离脂肪酸刺激的p-JNK的表达均有显著抑制作用(P均<0.01)。见图1、图3。
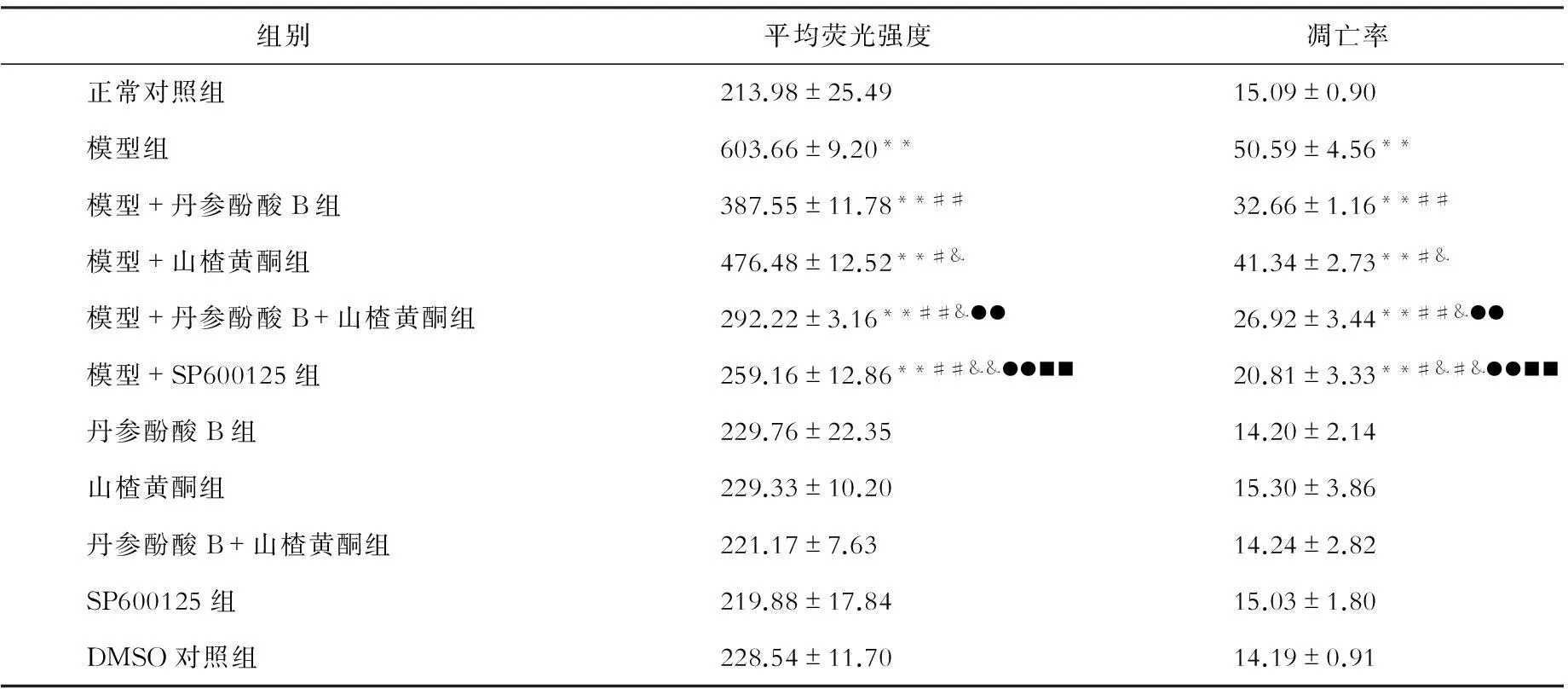
表3 各组原代肝细胞核平均荧光强度及凋亡率的比较
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
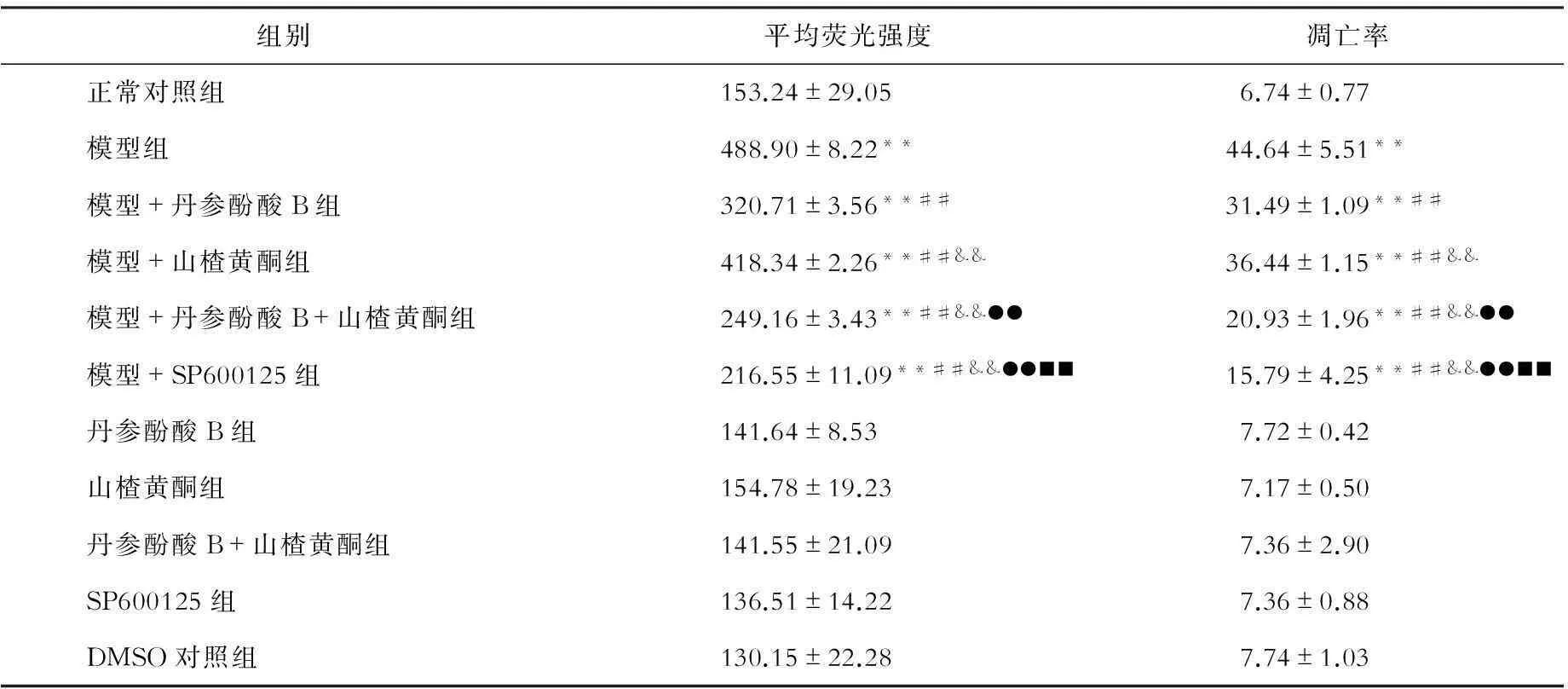
表4 各组HepG2细胞核平均荧光强度及凋亡率的比较
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
24 h HepG2细胞p-JNK表达结果见图2,图4,各组比较,趋势与原代肝细胞一致。
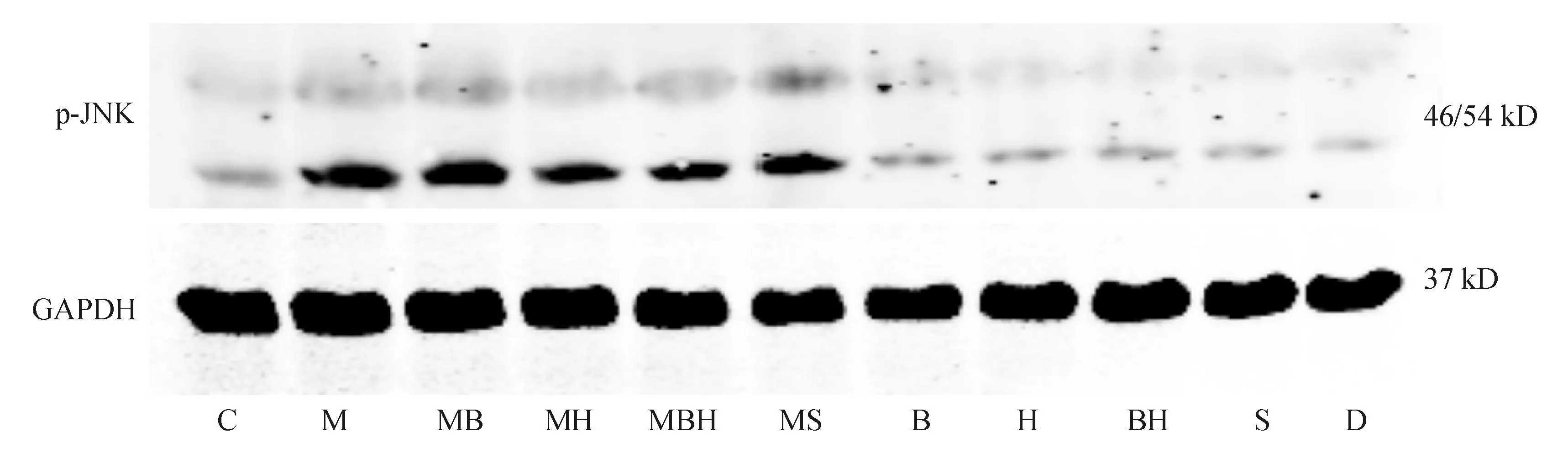
C:正常对照组;M:模型组;MB:模型+丹参酚酸B组;MBH:模型+丹参酚酸B+山楂黄酮组;MS:模型+SP600125组;B:丹参酚酸B组;H:山楂黄酮组;BH:丹参酚酸B+山楂黄酮组;S:SP600125组;D:DMSD对照组
图1Western blot法检测各组原代肝细胞p-JNK表达情况
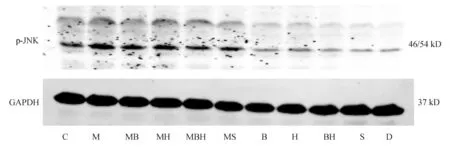
C:正常对照组;M:模型组;MB:模型+丹参酚酸B组;MBH:模型+丹参酚酸B+山楂黄酮组;MS:模型+SP600125组;B:丹参酚酸B组;H:山楂黄酮组;BH:丹参酚酸B+山楂黄酮组;S:SP600125组;D:DMSD对照组
图2Western blot法检测各组HepG2细胞p-JNK表达情况
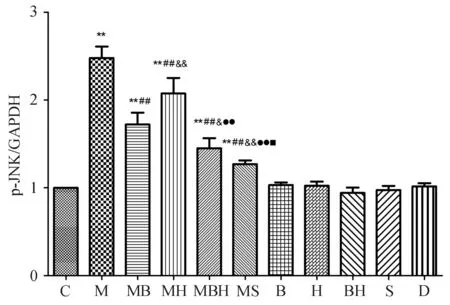
C:正常对照组;M:模型组;MB:模型+丹参酚酸B组;MBH:模型+丹参酚酸B+山楂黄酮组;MS:模型+SP600125组;B:丹参酚酸B组;H:山楂黄酮组;BH:丹参酚酸B+山楂黄酮组;S:SP600125组;D:DMSD对照组
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
图3原代肝细胞p-JNK的半定量分析图
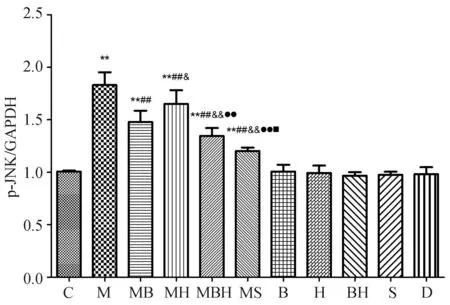
C:正常对照组;M:模型组;MB:模型+丹参酚酸B组;MBH:模型+丹参酚酸B+山楂黄酮组;MS:模型+SP600125组;B:丹参酚酸B组;H:山楂黄酮组;BH:丹参酚酸B+山楂黄酮组;S:SP600125组;D:DMSD对照组
注:与正常对照组相比,**P<0.01;与模型组相比,##P<0.01,#P<0.05;与模型+丹参酚酸相比,&&P<0.01,&P<0.05;与模型+山楂黄酮相比,●●P<0.01;与模型+丹参酚酸+山楂黄酮组相比,■■P<0.01
图4HepG2细胞p-JNK的半定量分析图
讨论
多项研究提示肝细胞凋亡与NASH发病相关。Feldstein等[2]应用TUNEL法检测NASH患者肝组织标本,发现肝细胞凋亡明显增加,且与NASH的严重性相关。南月敏等[8]采用高脂、胆碱-甲硫氨酸缺乏饮食方法建立小鼠NASH模型,TUNEL分析结果显示肝细胞凋亡为本模型的突出病理组织学特征。
而游离脂肪酸的增加对肝细胞的损伤作用在NASH的发病中起着重要作用。此外,游离脂肪酸还具有内毒素、促炎因子、肝星状细胞增殖因子、细胞凋亡诱导剂的作用[9,10]。游离脂肪酸对细胞的毒性作用与其种类相关,如油酸、亚油酸等多不饱和脂肪酸可通过激活肝内脂质过氧化反应促进酒精、四氯化碳引起的大鼠肝损伤[11,12];棕榈酸通过激活内质网应激反应,活化JNK信号通路,上调促凋亡蛋白的表达,诱导细胞脂性凋亡[13]。
本研究油红O染色结果显示,与正常对照组相比,游离脂肪酸(油酸+棕榈酸)分别作用于原代肝细胞、HepG2细胞24 h后,细胞内脂滴含量显著增加(P均<0.01)。以ELISA法检测原代肝细胞、HepG2细胞凋亡,结果显示与正常对照组相比,游离脂肪酸组原代肝细胞、HepG2细胞吸光值显著增高,细胞凋亡明显(P均<0.01),且高内涵筛选Hoechst33258染色观察原代肝细胞、HepG2细胞凋亡结果显示,与正常对照组相比,游离脂肪酸组细胞核平均荧光强度值及凋亡率明显增高(P均<0.01)。上述结果表明,游离脂肪酸能诱导原代肝细胞、HepG2细胞脂质代谢紊乱且促进其凋亡。
丹参酚酸B(Sal B)是丹参的主要水溶性有效成分之一,能减轻D-半乳糖胺大鼠肝细胞坏死,显著降低血清ALT和AST含量[6],能抑制体外传代培养的肝星状细胞的增殖,抑制肝星状细胞生成细胞外基质,减少胶原纤维的沉积[7]。
山植黄酮则可显著降低血清TC、TG、HDL -C水平[14];山楂叶总黄酮还能显著改善NASH大鼠的炎症程度,能提高大鼠的血清SOD水平,降低MDA和TNF-α的含量,降低肝组织NF-κB p65、IκBα基因的 mRNA和蛋白表达[15]。
本研究油红O染色结果显示,与游离脂肪酸组相比,游离脂肪酸+丹参酚酸B组+山楂黄酮组原代肝细胞、HepG2细胞内脂滴显著减少(P均<0.01)。以ELISA法检测原代肝细胞、HepG2细胞凋亡结果显示,与游离脂肪酸组比较,游离脂肪酸酸+丹参酚酸B+山楂黄酮组原代肝细胞、HepG2细胞吸光值显著降低,细胞凋亡程度明显减轻(P<0.05,P<0.01)。且高内涵筛选Hoechst33258染色观察原代肝细胞、HepG2细胞凋亡显示,与游离脂肪酸组比较,游离脂肪酸酸+丹参酚酸B+山楂黄酮组原代肝细胞、HepG2细胞细胞核平均荧光强度明显降低,细胞凋亡程度减轻(P<0.01)。上述结果表明丹参酚酸B+山楂黄酮可以减少肝细胞脂滴形成,改善肝细胞脂质代谢紊乱,同时抑制游离脂肪酸诱导的细胞凋亡。
目前研究认为多种刺激因素可引起从内质网到胞质和胞核的信号传导, 最终引起细胞凋亡或适应性存活,而JNK的激活通路是ERS(由各种原因引起的内质网中出现错误折叠与未折叠蛋白在腔内聚集以及Ca2+平衡紊乱的状态)诱发凋亡的途径之一[16]。
本研究油红O染色结果,与游离脂肪酸组相比,游离脂肪酸酸+SP600125组(JNK通路抑制剂) 原代肝细胞、HepG2细胞内脂滴显著减少(P均<0.01);SP600125组原代肝细胞、HepG2细胞与正常对照组相比,细胞内脂滴含量无差异统计学意义。
以ELISA法检测原代肝细胞、HepG2细胞凋亡显示,与游离脂肪酸组相比,游离脂肪酸+SP600125组原代肝细胞、HepG2细胞吸光值明显降低,凋亡明显减轻(P均<0.01);SP600125组原代肝细胞、HepG2细胞与正常对照组比较,细胞凋亡程度差异无统计学意义。且高内涵筛选Hoechst33258染色观察原代肝细胞、HepG2细胞凋亡显示,与游离脂肪酸组相比,游离脂肪酸+SP600125组细胞核平均荧光强度显著降低,凋亡明显减轻(P<0.01)。SP600125组与正常对照组相比,核平均荧光强度值差异无统计学意义。上述结果表明,SP600125亦可以改善游离脂肪酸诱导的细胞凋亡。提示JNK通路激活在游离脂肪酸诱导的细胞凋亡中起促凋亡作用。
Western blot检测JNK结果显示,药物作用24 h后,游离脂肪酸均可显著促进原代肝细胞及HepG2细胞p-JNK的表达(P均<0.01),即游离脂肪酸活化JNK的表达。结合油红O染色、ELISA及高内涵筛选Hoechst33258染色结果,说明游离脂肪酸诱导的JNK持续活化介导了细胞凋亡。JNK通路抑制剂SP600125对经游离脂肪酸刺激、24 h原代肝细胞及HepG2细胞内p-JNK的表达有显著抑制作用(P均<0.01),进一步证明JNK通路的激活参与了游离脂肪酸诱导的细胞凋亡。但油红O染色、ELISA及高内涵筛选Hoechst33258染色结果显示,SP600125并不能完全抑制游离脂肪酸诱导的细胞凋亡,推测可能还存在非JNK途径介导游离脂肪酸诱导的细胞凋亡途径。游离脂肪酸+丹参酚酸B+山楂黄酮组与游离脂肪酸组比较,丹参酚酸B+山楂黄酮可以抑制经游离脂肪酸刺激的24 h原代肝细胞及HepG2细胞内p-JNK蛋白表达(P均<0.01)。即丹参酚酸B和山楂黄酮抑制JNK的活化。提示丹参酚酸B和山楂黄酮对游离脂肪酸诱导的细胞凋亡的拮抗作用是通过抑制JNK信号途径实现的。丹参酚酸B和山楂黄酮抑制JNK活化是其抑制游离脂肪酸诱导的细胞凋亡的机制之一。
参考文献
1 de Alwis NM,Day CP.Non-alcoholic fatty liver disease:the mist gradually clears. J Hepatol,2008,48(Suppl 1):S104-S112.
2 Feldstein AE,Canbay A,Angulo P,et al.Hepatocyte apoptosis and fas expression are prominent features of human nonalcoholic steatohepatitis. Gastroenterology, 2003, 125: 437-443.
3 Kwan HY, Wang FF, Yang Z, et al. Inhibition of DNA-dependent protein kinase reduced palmitate and oleate-induced lipid accumulation in HepG2 cells. Eur J Nutr, 2013, 52: 1621-1630.
4 柳嘉, Popovich DG, 景浩. 山楂黄酮提取物的抗氧化活性和对癌细胞生长抑制作用. 食品科学,2010,03:220-223.
5 Pagliassotti MJ,Wei Y,Wang D.Insulin protects liver cells from saturated fatty acid-induced apoptosis via inhibition of c-Jun NH2 terminal kinase activity. Endocrinology,2007,148:3338-3345.
6 徐列明,刘成,刘平.丹参酚酸B对大鼠传代肝贮脂细胞增殖、形态和合成细胞外基质的影响.中华肝脏病杂志,1996,4:86-89.
7 薛冬英,洪嘉禾,徐列明.丹参酚酸B对大鼠肝星状细胞中丝裂原激活的蛋白激酶通路的抑制作用.中华肝脏病杂志,2004,12:471-474.
8 南月敏,乔梁,于君,等.Fas及其配体诱导非酒精性脂肪性肝炎肝细胞凋亡.中华消化杂志,2006,26:841-842.
9 徐正婕,范建高,王国良.游离脂肪酸在脂肪性肝炎发病中的作用.中华肝脏病杂志,2000,8:127-128.
10Galli A,Price D,Crabb D.High-level expression of rat class I alcohol dehydro- genase is sufficient for ethanol-induced fat accumulation in tranduced Hela cells. Hepatology,1999,29:1164-1170.
11Zeisel SH,da Costa KA,Aldright CD,et al.Choline and hepato-carinogenesis in the rat.Adv Exp Med Biol,1995,375:65-74.
12Akazawa Y,Cazanave S,Mott JL,et al.Palmitoleate attenuates palmitate-induced Bim and PUMA up-regulation and hepatocyte lipoapoptosis.J Hepatol,2010,52:586 -593.
13Liu P,Hu YY,Liu C,et al.Clinical observation of salvianolic acid B in treatment of liver fibrosis in chronic hepatitis B.World J Gastroenterol,2002,8:679-685.
14谢伟华,孙超,刘淑敏,等. 山楂黄酮对高脂血症模型小鼠血脂及生脂基因转录表达的影响.中国中药杂志,2009,2:224-229.
15严茂祥, 陈芝芸,何蓓晖. 山楂叶总黄酮对非酒精性脂肪性肝炎大鼠肝组织NF-κB及其抑制物表达的影响. 中华中医药杂志, 2009,2:139-143.
16Pagliassotti MJ,Wei Y,Wang D.Insulin protects liver cells from saturated fatty acid-induced apoptosis via inhibition of c-Jun NH2 terminal kinase activity. Endocrinology,2007,148:3338-3345.
(本文编辑:钱燕)
更正启事
《肝脏》2016年第21卷第2期第123~125页发表了湖北省鄂州市中心医院儿科高明、任章平撰写的论文《熊去氧胆酸联合护肝药物对婴儿肝炎综合征生化指标的影响》。按作者要求,添加通信作者:任章平,Email:chinaren1001@qq.com。特此更正。
《肝脏》编辑部2016年3月28日
Effects of salvianolic acid B and hawthorn flavone on lipid deposition and apoptosis of hepatocytes in rats induced by free fatty acids
XUEDong-ying,XIBo-ren,ZHANGJie,YEJun.
DepartmentofInfectiousDisease,PuTuoHospital,ShanghaiUniversityofTraditionalChineseMedicine,Shanghai200062,China
【Abstract】ObjectiveTo investigate the effects of salvianolic acid B and hawthorn flavone on free fatty acids (FFA)-induced lipid deposition and apoptosis of hepatocytes in rats, and its potential mechanism. MethodsHepG2 cells and primary cultured hepatocytes isolated from male SD rats by collagenase perfusion were subjected for this study. Oleic acid and palmitic acid (O∶P=2∶1) were used to induce nonalcoholic steatohepatitis (NASH) in primary cultured hepatocytes and HepG2 cells, respectively. The cell models were divided into 11 groups, including normal control group, FFA group, FFA + salvianolic acid B group, FFA+ hawthorn flavone group, FFA + salvianolic acid B + hawthorn flavone group, FFA+ SP600125 group, salvianolic acid B group, hawthorn flavone group, hawthorn flavone + salvianolic acid B group, SP600125 group and DMSO control group, respectively. Correspondingly, the cell models of NASH in primary cultured hepatocytes and HepG2 cells were stimulated with different doses of FFA (250 μmol/L and 500 μmol/L ), salvianolic acid B (10-6mol/L), hawthorn flavone (5 μg/mL) and SP600125 (10 μmol/L), respectively. In order to evaluate the therapy effects of salvianolic acid B and hawthorn flavone against NASH, the following tests were performed: Oil Red O staining for lipid deposition, and enzyme-linked immune-sorbent assay (ELISA), chromatin dye and Hoechst 33258 staining for apoptosis. In addition, it was speculated that the therapy effect was associated with c-Jun N-terminal kinase (c-JNK), which was verified by western blot to assess the expression of phosphorylated-JNK (p-JNK). ResultsAfter 24h of incubation with FFA alone, lipids in primary cultured hepatocytes and HepG2 cells were significantly increased (P<0.01). Moreover, salvianolic acid B, hawthorn flavoneor and SP600125 sharply reduced lipids deposition, respectively (P<0.01,P<0.05). The intracellular lipids in DMSO control group had no significant difference than that in normal control group. ELISA showed that FFA induced apoptosis in primary hepatocytes and HepG2 cells (P<0.01). Meanwhile, salvianolic acid B, hawthorn flavone or SP600125 could significantly protect liver from FFA-induced apoptosis (P<0.05, P<0.01). Through chromatin dye and Hoechst 33258 staining, FFA-induced apoptosis in primary cultured hepatocytes and HepG2 cells was also observed under high content screening (HSC) (P<0.01). Salvianolic acid B and hawthorn flavone could significantly reduce the apoptosis (P<0.05, P<0.01), respectively. Western blot analysis showed that FFA treatment led to a significant increase in c-JNK activity (P<0.01) in primary cultured hepatocytes and HepG2 cells, while total JNK levels remained unchanged. Meanwhile, salvianolic acid B and hawthorn flavone could significantly reduce the JNK activity. ConclusionSalvianolic acid B and hawthorn flavone could alleviate FFA-induced apoptosis and lipid metabolism disorders, respectively. Furthermore, they could also inhibit JNK activity, which reveals an important molecular mechanism for treatment of NASH.
【Key words】Nonalcoholic steatohepatitis;Salvia;Salvianolic acid B;Free fatty acid; Hepatocyte apoptosis;c-Jun N-terminal kinase
(收稿日期:2015-01-05)
Corresponding author:YE Jun, ZHANG Jie, Email: pzxgrk@163.com
通信作者:叶军,张洁,Email:pzxgrk@163.com
基金项目:上海市普陀区科委课题(2010PTKW004)
