朝营小流域石漠化治理区土壤动物群落结构
2016-05-03陈航陈浒王鹏举周政
陈航+陈浒+王鹏举+周政


摘要: 为了解贵州典型喀斯特石漠化治理区土壤动物群落结构,于2013年4月和8月对撒拉溪示范区朝营小流域不同等级石漠化样地的土壤动物进行了研究。共获得土壤动物976头,分属于3门10纲23目。结果显示,各样地以蜱螨目、弹尾目和双翅目为优势类群。水平结构上,不同季节不同等级石漠化土壤动物优势类群基本一致,而在个体数量表现常见类群和稀有类群的数量变化较大。垂直结构方面,2个季节土壤动物群落在0~10 cm土层均表现出明显的表聚性,个体数的表聚性强于类群数的,且在不同等级石漠化样地表现出较大差异。除个别样地外,土壤动物群落多样性指数、均匀性指数和丰富度指数均表现为春季大于夏季,各样地间土壤动物群落相似性指数在春季和夏季差异不大。
关键词: 土壤动物;群落结构;多样性;朝营小流域;石漠化治理
中图分类号: S154.5 文献标志码: A 文章编号:1002-1302(2016)03-0312-06
土壤动物是陆地生态系统中重要的生物组成成分,对环境变化敏感,其生物类群或群落组成等能够反映环境的细微变化[1-3]。在喀斯特生态环境中土壤动物是喀斯特生态系统主要的分解者,它们与微生物共同担负着喀斯特生态系统物质循环和能量流动的重要使命[4-5]。近年来,人们对森林生态系统、农业生态系统、不同土地利用方式和城市生态系统土壤动物进行了大量的研究[6-9]。对喀斯特生态系统的土壤动物的研究相对较少,陈浒等对贵州典型喀斯特地区土壤动物的研究表明,土壤动物种类数、个体数量随着纬度的增加而增加,土壤动物群落在水平分布上的差异与人类干预生境的程度和石漠化治理的生态效应差异有关[10]。熊康宁等对喀斯特石漠化治理区土壤动物生态功能研究表明,土壤动物以杂食性和植食性功能类群为主,捕食性和腐蚀性功能类群分布较少,随着石漠化程度的加深,物种数和个体数有减少趋势[11]。王仙攀等对喀斯特石漠化地区土壤动物培育发现,不同等级石漠化对土壤动物功能类群分布有影响,土壤动物种类和数量分布可以揭示石漠化生态恢复现状[12]。对国家石漠化重点治理区典型喀斯特高原山地地貌的撒拉溪示范区朝营小流域春夏两季土壤动物群落进行研究,以了解该区的土壤动物群落结构,为喀斯特地区土壤动物的保护和生态系统的科学恢复提供数据支撑。
1 材料与方法
1.1 研究区概况
撒拉溪示范区朝营小流域是国家喀斯特石漠化综合治理的重点区域,代表喀斯特高原山地轻-中度石漠化类型,平均海拔1 600 m,位于贵州省毕节市,处于乌江上游六冲河流域支流区,地理坐标为105°01′11.79″~105°08′38.34″E、27°11′09.09″~27°17′27.88″N,总面积5 633.30 hm2,石漠化面积 2 542.01 hm2。其中,轻度、中度和强度石漠化面积分别占流域石漠化面积的60.93%、28.72%、10.35%。
该区属亚热带季风湿润气候,年平均气温约为12.8 ℃,无霜期245 d,年平均日照时数为1 360 h,多年平均降雨量在984.4 mm,流域内阴雨天气较多,日照少,热量不足。岩石主要以碳酸盐岩为主,土壤以黄壤为主。植被为亚热带常绿落叶或针阔混交林,以次生林为主,并大多分布于山体上部。
1.2 研究方法
1.2.1 样地设置 参照熊康宁等的石漠化等级划分方法[13]划分石漠化类型。在对研究区详细调查的基础上,选择地势较为平坦,综合考虑小气候、土地利用类型、植被等因素的潜在石漠化、轻度石漠化、中度石漠化和强度石漠化典型样地进行土壤动物监测,共设立4个研究样方,每个样方的面积为10 m ×10 m,样地环境情况见表1。
1.2.2 样品采集与土壤动物分离 2013年4月和8月分别在研究区4个不同等级石漠化样地对土壤动物进行调查。样品采集参照《土壤动物研究方法手册》[14],采取多点取样。在每个石漠化样地设置3个取样点,拣去表面上的新鲜凋落物,每个样点按10 cm×10 cm的面积取定量土样,每层深度为 5 cm,按照0~5、5~10、10~15 cm分3层取样,装入布袋并进行样品编号。大型土壤动物于野外用镊子直接拣取,中小型土壤动物则将土壤样品带回室内后采用Tullgren法分离。所采动物均用75%的乙醇固定保存,根据《中国土壤动物检索图鉴》[15]和《昆虫形态分类学》[16]在OLYMPUS SZ61双目体视显微镜和OLYMPUS CX31生物显微镜下对土壤动物进行分类鉴定和数量统计。
1.2.3 数据处理 土壤动物群落多样性参数采用计算公式见式(1)至式(4)。
Shannon-wiener 多样性指数[17]:
2 结果与分析
2.1 土壤动物群落组成
4个样地2季采样获土壤样本72个,经分离后获得土壤动物976头,分属于3门10纲23目(表2)。其中,优势类群为蜱螨目、弹尾目和双翅目幼虫3类,个体数分别占总捕获量的35.66%、14.75%、27.97%;常见类群为膜翅目、鞘翅目幼虫、鞘翅目成虫、鳞翅目幼虫、线虫和蜘蛛目6类,个体数占总捕获量的16.39%;其余为稀有类群,共14类,其类群数达总类群数的60.87%,但个体数仅占总捕获量的5.53%。优势类群和常见类群个体数量多、分布广,构成了不同等级石漠化样地土壤动物群落的主体。
各样地中,(1)潜在石漠化样地捕获土壤动物数量和种类均最多,2次共捕获土壤动物18类,307头。优势类群为蜱螨目、弹尾目、双翅目幼虫和膜翅目4类,个体数占该样地捕获总量的85.34%,常见类群为鞘翅目幼虫、鞘翅目成虫、蜘蛛目、寡毛纲和等翅目等5类,个体数占捕获总量的 9.11%,其余为稀有类群,含鳞翅目幼虫、双翅目、半翅目等9类,个体数占捕获总量的占5.55%。(2)轻度石漠化样地共捕获土壤动物208头,16类,优势类群为蜱螨目、弹尾目和双翅目幼虫3类,个体数占该样地捕获总量的83.66%,鞘翅目幼虫、鞘翅目成虫和蜘蛛目等3类为常见类群,个体数占捕获总量的 9.61%,稀有类群为膜翅目、鳞翅目幼虫和唇足纲等10类,个体数占捕获总量的6.72%。(3)中度石漠化样地捕获的土壤动物数量最少,2次共捕获土壤动物197头,土壤动物优势类群和轻度石漠化样地相似,蜱螨目与其他样地相比,占样地捕获总量的比例最高,为54.31%;常见类群为膜翅目、鞘翅目幼虫、鞘翅目、线虫纲、双尾目、同翅目和寡毛纲7类,个体数占捕获总量的15.24%,稀有类群为蜘蛛目、鳞翅目幼虫和半翅目等6类,个体数占捕获总量的3.06%。(4)强度石漠化样地捕获的土壤动物类群数最少,2次共捕获土壤动物15类,优势类群为蜱螨目和双翅目幼虫2类,个体数占捕获总量的74.63%,常见类群为弹尾目、膜翅目、鞘翅目幼虫、鞘翅目和鳞翅目幼虫5类,个体数占捕获总量的21.59%,稀有类群为蜘蛛目、双尾目、同翅目和等翅目等8类,个体数占捕获总量的3.79%,该样地双翅目幼虫个体数量最多,所占捕获量比例也是各样地中最大的。
2.2 土壤动物群落多样性
对4类样地土壤动物多样性指数(H′)、均匀性指数(E)、优势度指数(C)、和丰富度指数(D)进行了测度,结果(表3)显示,不同等级石漠化样地的土壤动物群落多样性指数存在明显的差异,表现在多样性指数和均匀性指数:潜在石漠化>轻度石漠化>中度石漠化>强度石漠化。而均匀性指数在0.55~0.66之间,总体表现较高,说明各样地土壤动物群落的均匀性较相近;优势度指数表现为强度石漠化>中度石漠化>轻度石漠化>潜在石漠化,与多样性指数和均匀性指数的变化趋势相反。丰富度指数表现为中度石漠化>轻度石漠化>强度石漠化>潜在石漠化,表现出与多样性指数不完全一致的规律。
从不同样地多样性的季节变化(表4)看,除强度石漠化样地外,Shannon-Wiener多样性指数H′在4月份均大于8月份,这与类群数和个体数的变化趋势几乎相反,夏季虽然土壤动物类群数最多,个体数最大,但由于蜱螨目、弹尾目等少数类群数量激增,致使土壤动物群落分布不均衡,因此H′指数小;而春季虽然土壤动物类群数和个体数均较少,但由于个体数分布较夏季均匀,因此H′指数大。Pielou均匀性指数与Shannon-Wiener多样性指数变化趋势基本一致,除强度石漠化样地外,均表现为4月份均大于8月份。Simpson优势度指数的变化趋势与前2个指数大致相反,除轻度石漠化和强度石漠化样地外,其他2个样地均表现为8月份大于4月份。Menhinick丰富度指数上,除强度石漠化样地外,其他样地的丰富度指数均表现为4月份大于8月份。可见,丰富度指数与多样性指数在季节上存在明显的正相关关系。
2.3 土壤动物群落空间分布
2.3.1 土壤动物水平分布 4种石漠化样地土壤动物类群数和个体数的水平分布不同(图1),个体数量上,春季(4月)表现为:强度石漠化>潜在石漠化>中度石漠化>轻度石漠化;夏季(8月)表现为:潜在石漠化>轻度石漠化>中度石漠化>强度石漠化。类群数上,在春季(4月)表现为:潜在石漠化>中度石漠化=强度石漠化>轻度石漠化;夏季(8月)为:潜在石漠化>轻度石漠化>中度石漠化>强度石漠化。各样地8月份土壤动物个体数量和类群数表现出相同变化趋势,4月则略有不同。
4种石漠化样地土壤动物种类和个体数量在季节变化上表现为:除强度石漠化样地外,其他3个样地土壤动物的个体数均表现为夏季(8月)>春季(4月)。土壤动物类群数方面,潜在石漠化样地和轻度石漠化样地表现为夏季(8月)>春季(4月),而中度石漠化和强度石漠化样地土壤动物类群数表现出相反的变化趋势。
分析4种不同样地土壤动物主要类群季节变化规律(表5),潜在石漠化样地4月双翅目幼虫个体数最多,占捕获量的40.21%;8月份蜱螨目个体数量最多,占捕获量39.52%,蜱螨目、弹尾目和双翅目幼虫在4、8月均为优势类群;8月份为优势类群的膜翅目,在4月份仅为常见类群;在4月份为常见类群的蜘蛛目、半翅目和寡毛纲,在8月份仅为稀有类群;4月份双尾目、线虫纲和蛭纲3类为常见类群,但在8月份未捕获到。轻度石漠化样地4、8月均以蜱螨目个体数最多,各占捕获量的43.40%、41.49%;蜱螨目和弹尾目在4、8月均为优势类群;8月份为优势类群的双翅目幼虫,在4月份为常见类群;在4月份为常见类群的双翅目、鳞翅目幼虫、线虫纲和等足目,在8月份未捕获。中度石漠化样地4、8月均以蜱螨目和弹尾目为优势类群;4月份为优势类群的双翅目幼虫,在8月份为常见类群;在4月份为常见类群的鞘翅目、鳞翅目幼虫、线虫纲、半翅目、寡毛纲和盲蛛目,在8月份未捕获到。强度石漠化样地4月双翅目幼虫个体数最多,占捕获量的65.60%,8月份以蜱螨目最多,占45.65%;4月份优势类群为蜱螨目和双翅目幼虫,8月份优势类群为蜱螨目、弹尾目和膜翅目。分析表明,2个季节土壤动物的优势类群基本一致,而常见类群和稀有类群数的变化导致了类群数的季节差异。
2.3.2 土壤动物垂直分布 4种不同等级石漠化样地4月和8月土壤动物个体数量和类群数的垂直分布(图2、图3)表明,2个季节土壤动物群落在0~10 cm土层整体上均随着土壤深度的增加而减少,表现出明显的表聚性,但在不同等级石漠化样地表现出较大差异。
4月份土壤动物个体数总体上在各土层均较少,其中潜在石漠化、轻度石漠化和中度石漠化样地在0~5 cm土层上个体数少,但在强度石漠化样地中却有所增加,这主要是由于双翅目幼虫数量较大所致。8月份,轻度石漠化和强度石漠化样地土壤动物个体数在10~15 cm多于5~10 cm,表现出逆分布现象。8月份潜在石漠化和轻度石漠化样地的个体数在0~5 cm、5~10 cm、10~15 cm 3个土层均比4月份多。8月份中度石漠化样地在5~10 cm和10~15 cm土层的个体数均比4月份少,强度石漠化样地8月份土壤动物个体数在0~5 cm和5~10 cm土层明显少于4月份。
土壤动物类群数方面,2个季节均表现为明显的表聚性,4月份潜在石漠化样地在0~5 cm土层土壤动物的类群数最多,轻度石漠化的类群数最少,轻度石漠化和中度石漠化样地土壤动物类群数在5~10 cm和10~15 cm土层相同。8月份潜在石漠化样地的类群数也最多,强度石漠化样地的类群数最少,轻度石漠化土壤动物的类群数在10~15 cm土层比 5~10 cm 多。除潜在石漠化样地外,其他3个样地土壤动物类群数在0~5 cm土层4月份均大于8月份,4个样地的土壤动物类群数在5~10 cm土层均表现为8月份大于4月份。上述结果表明,在不同等级石漠化样地土壤动物表聚性的程度有所差别,个体数量的表聚性要强于类群数的表聚性。
2.4 土壤动物群落相似性
Jacard相似性系数的判读标准为:计算值在0.75~1.0为极相似,0.50~0.74为中等相似,0.25~0.49为中等不相似,0~0.24为极不相似[22]。各样地土壤动物群落的相似性进行分析显示(表6),相似性系数均在0.60~0.73之间,说明4种不同等级石漠化样地间的土壤动物群落均属中等相似,其中相似性最大的为中度石漠化和强度石漠化样地,其相似性指数为0.722 2,轻度石漠化和中度石漠化相似性最小,相似性指数为0.6。
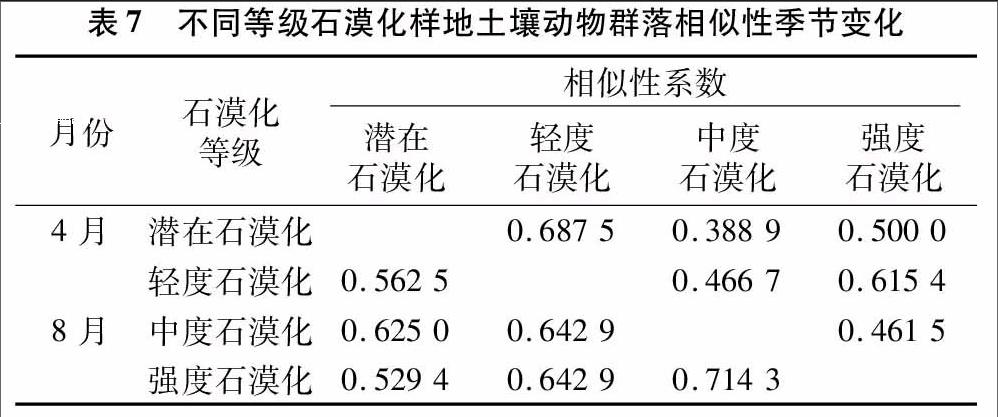
4种不同样地相似性指数的季节变化来看(表7),在春季(4月),4种不同等级石漠化样地间土壤动物群落Jaccard相似性系数均在0.52~0.72之间,表现为中等相似。夏季(8月),潜在石漠化和轻度石漠化样地的q值最大,属中等相似,其次是轻度石漠化和强度石漠化样地,潜在石漠化和强度石漠化样地,各对均属于中等相似;而中度石漠化样地与潜在石漠化、轻度石漠化和强度石漠化样地,均属中等不相似。上述表明,春季和夏季不同等级石漠化样地间的土壤动物群落的差异不大。
3 结论与讨论
朝营小流域不同等级石漠化样地土壤动物优势类群为蜱螨目、弹尾目和双翅目幼虫,这与赵基等的研究结果[25]相似。这3类物种在石漠化样地的生存、繁衍和对凋落物的分解能力强,已成为石漠化生态恢复过程中改良土壤的主要物种。常见类群为膜翅目、鞘翅目幼虫、鞘翅目成虫、鳞翅目幼虫、线虫和蜘蛛目6类,稀有类群14类,常见类群和稀有类群的变化较大,对环境变化反映敏感,这可能与各样地的微地域自然环境差异及各样地不同生态治理模式有关,可把常见类群和稀有类群的变化作为石漠化地区局部环境变化的指示性动物。
由于各研究区的自然环境不同,土壤动物水平分布也有差别。各样地春季(4月)土壤动物个体数和类群数的变化趋势有所不同,在个体数上强度石漠化最多,在类群数上潜在石漠化最多,轻度石漠化样地个体数和类群数均最少,这与各样地单类土壤动物个体数的激增和人类活动的干扰有关。夏季(8月),随石漠化程度的加深,土壤动物个体数和类群数均有减少的趋势,表明土壤动物的恢复响应速度变慢。
季节变化上,潜在石漠化样地和轻度石漠化样地土壤动物个体数和类群数均表现为夏季(8月)>春季(4月),强度石漠化样地表现出相反的趋势。2个季节土壤动物优势类群基本一致,而常见类群和稀有类群数的变化导致了类群数的季节差异。综合样地的环境差异分析认为,潜在石漠化和轻度石漠化样地的植被覆盖度较高,人类耕作的影响,使得土壤的肥力增强,加上该区降水主要集中在夏季,且雨热同期,有利于凋落物的分解,系统内养分流转速度快,因此土壤动物在夏季比较活跃;中度石漠化和强度石漠化样地由于岩石裸露率高,且坡度大,土层较薄,土壤贫瘠,夏季温度高,降水比较集中,雨水对地面的冲刷严重,使表面土壤水分饱和或过饱和,造成生活在此期间的中小型土壤动物逃逸或死亡,加上人为放牧的影响,使得该区土壤动物类群和个体数减少。
垂直结构方面,2个季节土壤动物群落在0~10 cm土层均表现出明显的表聚性,在不同等级石漠化样地其表聚性的程度有所差别,个体数量的表聚性要强于类群数的表聚性,这可能与石漠化治理后土壤表层的营养含量丰富、土壤水热条件以及理化特征的垂直差异有密切关系。夏季(8月),轻度石漠化和强度石漠化样地土壤动物个体数表现出逆分布现象,这可能与取样点微环境差异有关。季节变化上,夏季(8月)潜在石漠化和轻度石漠化样地的个体数和类群数在垂直分布上均比4月份多,表明夏季降水较多,植物生长旺盛,比较适合土壤动物生长。8月份中度石漠化样地在5~10 cm和10~15 cm土层的个体数均比4月份少,强度石漠化样地8月份土壤动物个体数在0~5 cm和5~10 cm土层明显少于4月份,可能由于中度石漠化和强度石漠化样地岩石裸露率高,植被稀疏,土壤贫瘠,气温相对较高和土壤干燥等引起。
从不同等级石漠化样地土壤动物群落多样性方面看,总体上,多样性指数与均匀性指数变化趋势基本一致,而与优势度指数则基本相反,与丰富度指数表现出不完全一致的规律。季节变化上,除个别样地外,多样性指数、均匀性指数和丰富度指数均表现为春季大于夏季。夏季虽然土壤动物类群数最多,个体数最大,但由于蜱螨目、弹尾目等少数优势类群数量占优势,致使土壤动物群落在类群数和个体数上分布不均衡。优势度指数的变化趋势与其他指数大致相反,可能与人类对不同等级石漠化土地的扰动程度差异和不同等级石漠化经不同模式生态治理后环境效应差异有关。相似性指数上,除个别样地外,春季和夏季不同等级石漠化样地间的差异不大,这可能与样地微地域环境的差异和人类活动的干扰等有关。
不同石漠化生境在土壤理化性质、生物因子等方面具有较大差异,石漠化及其生态恢复进程中不同的植被修复模式与管理等对土壤环境的影响较大,在土壤动物群落结构及多样性上均表现出不同程度的响应。石漠化地区土壤的恢复是一个长期、复杂的过程,土壤动物对地上植被恢复过程中的变化响应最为敏感,因此在某种意义上可以通过土壤动物在种群组成、个体数量、空间分布及优势类群的变化来监测石漠化生态恢复的状态。
参考文献:
[1]尹文英.土壤动物学研究的回顾与展望[J].生物学通报,2001,36(8):1-3.
[2]朱永恒,赵春雨,王宗英,等. 我国土壤动物群落生态学研究综述[J].生态学杂志,2005,24(12):1477-1481.
[3]李 涛,刘苑秋,郭圣茂,等. 瑞昌石灰岩红壤区退耕还林土壤动物群落特征[J].应用生态学报,2012,23(4):910-916.
[4]黄丽荣,张雪萍. 大兴安岭北部森林生态系统土壤动物的功能类群及其生态分布[J].土壤通报,2008,39(5):1017-1022.
[5]李金霞,王继仁,程 莉.稀土尾矿坝毗邻区大型土壤动物与土壤因子关系研究[J].干旱区资源与环境,2010,24(10):171-177.
[6]廖崇惠,李健雄,黄海涛.南亚热带森林土壤动物群落多样性研究[J].生态学报,1997,17(5):99-105.
[7]殷秀琴,王海霞,周道玮. 松嫩草原区不同农业生态系统土壤动物群落特征[J].生态学报,2003,23(6):1071-1078.
[8]王广力,王 勇,韩立亮,等. 洞庭湖区不同土地利用方式下的土壤动物群落结构[J].生态学报,2005,25(10):2629-2636.
[9]秦 钟,章家恩,李庆芳.城市化地区不同生境下中小型土壤动物群落结构特征[J].应用生态学报,2009,20(12):3049-3056.
[10]陈 浒,樊云龙,赵志成,等. 贵州典型喀斯特地区土壤动物生态地理研究[J].中国农学通报,2011,27(20):208-215.
[11]熊康宁,陈 浒,王仙攀,等. 喀斯特石漠化治理区土壤动物的时空格局与生态功能研究[J].中国农学通报,2012,28(23):259-265.
[12]王仙攀,陈 浒,熊康宁,等. 喀斯特石漠化地区土壤动物功能类群及培育研究[J].干旱区资源与环境,2012,26(12):191-195.
[13]熊康宁,黎 平,周忠发,等. 喀斯特石漠化的遥感-GIS典型研究——以贵州省为例[M].北京:地质出版社,2002:23-28.
[14]尹文英.土壤动物研究方法手册[M].北京:科学出版社,1998.
[15]尹文英,胡圣豪,沈韫芬,等. 中国土壤动物检索图鉴[M].北京:科学出版社,1998:51-387.
[16]忻介六,杨庆爽,胡成业. 昆虫形态分类学[M].上海:复旦大学出版社,1985:99-366.
[17]Peet R K. The measurement of species of diversity[J]. Annual Review of Ecology and Systematics,1974,5:285-307.
[18]Pielou E C. The measurement of diversity in different types of biological collections[J]. Journal of Theoretical Biology,1966,13:131-144.
[19]Simpson E H. Measurement of diversity[J]. Nature,1949,163:688.
[20]马克平. 生物群落多样性的测度方法Ⅰ. α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[21]马克平,刘玉明. 生物群落多样性的测度方法Ⅰ. α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.
[22]傅荣恕,尹文英.伏牛山地区土壤动物群落的初步研究[J].动物学研究,1999,20(5):396-397.
[23]王广力,王 勇,韩立亮,等. 洞庭湖区不同土地利用方式下的土壤动物群落结构[J].生态学报,2005,25(10):2629-2636.
[24]刘长海. 陕北枣林土壤动物群落的结构及其季节动态[D].北京:北京林业大学,2008.
[25]赵 基,陈 浒,徐 玲,等. 朝营小流域不同等级石漠化土壤动物群落结构[J].贵州农业科学,2013,41(6):127-130.
