赤霉素与多效唑协同影响超级稻甬优12产量形成的作用途径分析
2016-04-29杨雪芹陈婷婷赵霞张彩霞杨永杰符冠富陶龙兴
杨雪芹 陈婷婷 赵霞 张彩霞 杨永杰 符冠富 陶龙兴
(中国水稻研究所 水稻生物学国家重点实验室, 杭州 310006; #共同第一作者; *通讯联系人, E-mail:lxtao@mail.hz.zj.cn)
赤霉素与多效唑协同影响超级稻甬优12产量形成的作用途径分析
杨雪芹#陈婷婷#赵霞张彩霞杨永杰符冠富陶龙兴*
(中国水稻研究所 水稻生物学国家重点实验室, 杭州 310006;#共同第一作者;*通讯联系人, E-mail:lxtao@mail.hz.zj.cn)
杨雪芹, 陈婷婷, 赵霞, 等. 赤霉素与多效唑协同影响超级稻甬优12产量形成的作用途径分析. 中国水稻科学, 2016, 30(1): 53-61.
摘要:以超级稻甬优12为材料,于水稻抽穗后3 d喷施3.2%赤霉素·多效唑(俗名粒粒饱,LLB),分析3.2%赤霉素·多效唑对超级稻甬优12产量形成的影响,并探讨其作用途径。研究表明,3.2%赤霉素·多效唑可在一定程度上提高水稻产量,2012-2013年的实际产量比对照(CK)分别高4.41%和4.31%,理论产量比对照分别高9.21%和6.26%。结实率和千粒重的增加是3.2%赤霉素·多效唑提高超级稻甬优12产量的主要因素。3.2%赤霉素·多效唑处理的结实率分别比对照高9.94%(2012年)和4.46%(2013年),而柱头花粉粒萌发率的提高是结实率增加的主要原因,因为和对照相比,3.2%赤霉素·多效唑处理对水稻花粉粒育性及柱头花粉粒数的影响不大,处理间差异不显著,但其柱头花粉萌发率显著高于对照。与对照相比,3.2%赤霉素·多效唑处理的千粒重分别增加0.39 g(2012年)和0.46 g(2013年),尤其弱势粒,其粒重增幅明显大于强势粒。此外,研究还表明,3.2%赤霉素·多效唑处理的剑叶叶绿素含量、光合速率及叶片比叶重在籽粒灌浆后期高于对照,而其籽粒和叶片可溶性糖含量比值低于对照,表明3.2%赤霉素·多效唑不仅可延缓叶片衰老,还可以促进光合同化物(可溶性糖)向籽粒转运。
关键词:3.2%赤霉素·多效唑; 超级稻; 产量; 结实率; 千粒重
水稻是我国主要的粮食作物之一,种植面积约占我国粮食作物总面积的30%,总产量约占粮食总产的40%[1]。中国是水稻生产和消费大国,50%以上人口以稻米为主食。据预测,到2030年我国人口将达16亿,届时即使耕地面积不减少,粮食作物综合生产能力仍需提高55%才能基本满足吃饭之需。
在新增粮食产量中,水稻将占40%以上[2]。在人增地减的国情下,培育超高产水稻品种,提高水稻单产,是确保我国粮食安全最主要的措施之一。因而,培育超级杂交水稻品种不失为一个很好的选择。
然而,超级稻品种存在的一些问题,严重阻碍了其产量潜力的发 挥。最为突出的是结实率较低且不稳定,同一品种在不同年度和地区间结实率大起大落[3-4]。其次为强势粒及弱势粒充实整齐性差[5-6]。关于超级稻弱势粒充实差、粒重低的原因,国内外学者已经进行了大量的研究,认为主要有以下几个方面的原因:1)源-库不协调,库过大,源供应物不足,抽穗时茎鞘贮藏物较少及最终运转率偏低[7];2)弱势粒开花后灌浆启动“滞后”,启动后活力低,即灌浆初期弱势粒库强小或蔗糖转化为淀粉的生化效率低[8-9];3)籽粒中较高的抑制型植物激素ABA和乙烯与促进型植物激素(生长素、细胞分裂素及赤霉素)的比例抑制弱势粒充实[10];4)“粒间顶端优势”是导致弱势粒结实较差的原因[11];5)灌浆初期弱势粒细胞壁转化酶、液泡转化酶、腺苷二磷酸葡萄糖焦磷酸化酶(AGP)和淀粉合成酶(StS)的基因表达量低[12]。Yang等[9]认为施适量ABA可有效改善弱势粒充实度,从而达到增产的效果。但也有研究认为ABA显著增加强势粒粒重,对弱势粒的影响不大,而GA可显著增加强势粒和弱势粒粒重[13]。另外,陶龙兴等[14]认为适量GA3和PP333复配(3.2%赤霉素·多效唑,俗称粒粒饱)可有效解决水稻特别是亚种间杂交水稻籽粒充实度差的结实生理障碍,主要表现为千粒重和结实率的增加。
3.2%赤霉素·多效唑是一种二元植物生长调节剂,主要活性成分是1.6%赤霉素与1.6%多效唑,在水稻齐穗前施用可延缓叶片衰老,调节激素平衡,增进灌浆[15-16]。童继平等[17]认为3.2%赤霉素·多效唑可增加水稻生长后期的生理活性,提高水稻主茎干质量和主茎穗重。虽然有关3.2%赤霉素·多效唑的研究甚多,但其在现代大穗型超级稻(例如甬优12)中的增产效果依然值得关注,尤其对小穗育性的作用机理还有待研究。此外,以往外源调控水稻亚种间结实障碍多侧重于籽粒灌浆,而对小穗育性的研究偏少,对水稻花器官(花粉粒育性、柱头花粉粒数及柱头花粉粒萌发率)的影响大为缺乏。因而,本研究以超级稻甬优12为材料,从水稻花粉粒育性、柱头花粉粒数及萌发、籽粒灌浆、干物质量积累及籽粒和叶片可溶性糖含量比值方面分析3.2%赤霉素·多效唑影响超级稻甬优12产量形成的作用途径,可为减轻大穗型超级稻结实障碍提供理论参考。
1材料与方法
1.1供试材料与试验设计
试验在中国水稻研究所富阳试验基地进行。供试材料为超级稻甬优12(甬粳2A×F5032),系浙江省宁波市农业科学研究院与宁波市种子有限公司共同选育的三系籼粳杂交稻组合。于2012年5月28日(2013年5月28日)播种,2012年6月22日(2013年6月24日)移栽,单本插,株距为25 cm×20 cm。试验田块土壤为青紫泥, 有机质含量 36.9 g/kg, 全氮 2.7 g/kg, 全磷 0.6 g/kg, 全钾 20.1 g/kg, 碱解氮 232 mg/kg , 铵态氮 9.7 mg/kg, 速效磷 25.2 mg/kg , 速效钾 65.0 mg/kg, pH 值为 6.5。以尿素、氯化钾、钙镁磷肥作为肥源,用量(折合成纯N、P2O5和K2O)分别为7.0 g/667 m2、7.5 kg/667 m2和7.5 kg/667 m2。。其中氮肥的50%作基肥,30%作分蘖肥,20%作穗肥;磷肥全部作基肥施用;钾肥基、穗肥各占50%。水稻抽穗时间为2012年9月13日和2013年9月14日,水稻抽穗后3 d喷施3.2%赤霉素·多效唑,使用量为37.5 g/667 m2,兑水30 L (产品建议使用量为25 g/667 m2,因粳稻对3.2%赤霉素·多效唑敏感性较低,使用浓度有所增加),3次重复,小区面积为200 m2。
1.2方法
1.2.1产量及产量构成
水稻成熟期取样测产,实际测产小区面积为25 m2,9次重复。此外,每小区随机取36丛水稻进行考种,考查项目为有效穗数、每穗粒数、结实率和千粒重。
1.2.2花粉粒育性
于齐穗期(喷施3.2%赤霉素·多效唑后4 d),取同一部位(穗上部第4个枝梗)未开放的颖花100个,每个颖花取2个花药,20个为1重复,用镊子轻轻挤压花药让花粉粒散出,经1% I2-IK 溶液染色后于显微镜(DM4000B, Leica, Wetzlar, Germany)下观察。
1.2.3柱头花粉粒数
于齐穗期(喷施3.2%赤霉素·多效唑后4 d)上午10:00-11:00取同一部位(穗上部第4个枝梗)正在开放的颖花100个,用镊子取出柱头置于1%苯胺蓝溶液染色10~12 h,之后在荧光显微镜(DM4000B, Leica, Wetzlar, Germany)下观察。
1.2.4柱头花粉萌发率
于齐穗期(喷施3.2%赤霉素·多效唑后4 d),上午10:00-11:00取同一部位(穗上部第4个枝梗)正在开放的颖花100个,用镊子取出柱头并置于体积比为3∶1乙醇乙酸固定液中固定24 h。用蒸馏水洗净后,于56℃恒温箱中用10 mol/L NaOH溶液透明10 min。再次清洗干净后置于0.5%苯胺蓝溶液中染色12 h,随后用荧光显微镜100倍视野(DM4000B, Leica, Wetzlar, Germany)观察。
1.2.5籽粒灌浆特征
选择生长整齐一致、同日开花的稻穗200个,挂上纸牌并标记开花日期。自开花至成熟每隔6 d取标记穗15个用于考查强弱势粒的粒重,并用Richards 生长方程W=A/(1+Be-kt)1/N对籽粒灌浆过程进行拟合,计算灌浆速率[18-19]。
1.2.6光合作用
叶片喷施3.2%赤霉素·多效唑后7、21、35及49 d,于上午9:00-11:00用便携式光合仪(LI-6400,Li-Cor Inc., Lincoln, NE, USA)测定水稻剑叶光合速率,并取样用95%乙醇提取法测定剑叶叶绿素含量。
1.2.7可溶性糖含量
叶片喷施3.2%赤霉素·多效唑后10、20、30及40 d,用蒽酮比色法分别测剑叶和籽粒中可溶性糖含量。
1.2.8干物质量
喷施3.2%赤霉素·多效唑前10 d(抽穗前7 d),喷施后3、13、23、33、43及53 d每小区取样12丛,样品分为叶片、茎鞘及穗3部分,置于105℃烘箱中杀青60 min,随后置于80℃恒温箱烘48 h至恒重。此外,考查水稻叶片叶面积指数及叶片比重。
1.2.9数据处理
采用Microsoft Excel和SPASS ver 11.5软件进行统计分析。
2结果与分析
2.1产量及其构成因子
如表1所示,抽穗初期喷施3.2%赤霉素·多效唑(粒粒饱或LLB,以下简称LLB)可显著提高超级稻甬优12的产量,实际产量比对照分别增加4.41%(2012年)和4.31%(2013年),理论产量则分别增加9.21%和6.26%。LLB对有效穗数影响不大,处理间差异不显著,但LLB处理的结实率和千粒重均显著高于对照。2012年和2013年LLB处理的结实率分别比对照高9.94%和4.46%,千粒重上则分别增加0.39 g和0.49 g。LLB对每穗粒数的影响存在年份间的差异,2012年LLB处理每穗粒数显著低于对照,但2013年则显著高于对照。
2.2花粉粒育性、柱头花粉粒数及柱头花粉萌发率
由图1可知:1)LLB对水稻花粉粒育性的影响不大,处理间差异不显著,且两个处理的花粉粒育性都在90%以上(图1-A、D、H);2)虽然LLB处理的柱头花粉粒数与对照没有显著差异,且都在60粒以上,但LLB处理略高于对照(图1-B、E、I);3)LLB可显著提高水稻柱头花粉萌发率,增幅达80%以上(图1-C、F、J)。
2.3籽粒灌浆特征
表2显示的是由Richards方程拟合的籽粒灌浆特征参数,籽粒灌浆拟合度均在0.95以上,灌浆过程符合Richards模型。从表中可看出,LLB处理强势粒和弱势粒的最终粒重(A值)均高于对照,增重分别为0.40 g和0.91 g。由籽粒灌浆动态可知(图2-A),籽粒灌浆前期(0~24 d)处理间粒重的差异不明显,开花24 d后LLB处理的强势粒和弱势粒粒重均高于对照。此外,强势粒和弱势粒的籽粒灌浆速率均在花后12 d达到峰值,但随后强势粒出现大幅度下降,而弱势粒则在开花后18 d才出现大幅度下降(图2-B)。
表13.2%赤霉素·多效唑对超级稻甬优12产量及其构成的影响
Table 1. Effect of 3.2%GA3·PP333on the grain yield and its components in super rice Yongyou 12.

年份Years处理Treatments单位面积穗数Paniclenumber(×106·hm-2)每穗粒数Grainnumberperpanicle结实率Seed-settingrate/%千粒重1000-grainweight/g理论产量Theoreticalgrainyield/(t·hm-2)实际产量Actualgrainyield/(t·hm-2)2012对照CK1.56±0.134a456.36±29.88a71.75±1.68b21.06±0.35a10.75±0.56b11.19±0.04bLLB1.51±0.081a444.00±17.28b81.69±2.20a21.45±0.21a11.74±0.38a11.68±0.09a2013对照CK1.71±0.053a440.02±0.90b78.59±1.87b21.13±0.15b12.47±0.43b11.60±0.18bLLB1.55±0.092a476.89±23.77a83.23±1.15a21.62±0.08a13.25±0.03a12.10±0.11a
同一年份同一列中,数据(平均值±标准误)后跟相同小写字母者表示在P=0.05水平上差异不显著(最小显著差法,n=6)。下同。
In a column,data (Mean±SE) followed by the same lowercase letters are not significantly different at 0.05 level by LSD(n=6) in 2012 or 2013. The same as below.

柱上相同小写字母者表示在P=0.05水平上差异不显著(最小值显著差法,n=30)。
The same lowercase letters above the bars indicate significant difference at 0.05 level(by LSD,n=30).
图13.2%赤霉素·多效唑对水稻花粉粒育性(A、D、H)、柱头花粉粒数(B、E、I)及柱头花粉萌发率(C、F、J)的影响
Fig. 1. Effect of 3.2% GA3·PP333(LLB) on the pollen viability (A,D,H), pollen numbers on the stigma (B,E,I) and pollen germination rate on the stigma (C,F,J) of rice.
2.4剑叶叶绿素、光合速率及比叶重
图3显示的是LLB处理后超级稻甬优12剑叶叶绿素含量、光合速率和叶片比叶重的变化趋势。从中可看出,籽粒灌浆初期剑叶叶绿素含量(图3-A)、光合速率(图3-B)及叶片比叶重(图3-C)处理间差异不明显,然而随着灌浆进程,处理间差异逐渐增大,花后49 d处理间差异达到最大值。
表23.2%赤霉素·多效唑对超级稻甬优12籽粒灌浆参数的影响
Table 2. Effect of 3.2%GA3·PP333(LLB) on the grain filling parameters of rice.

粒位Grainposition处理TreatmentsAbkmR2强势粒Superiorgrain对照CK21.922.290.291.240.9900LLB22.320.940.240.840.9818弱势粒Inferiorgrain对照CK18.981.060.180.530.9584LLB19.892.490.201.060.9669
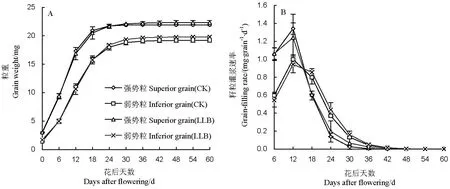
图23.2%赤霉素·多效唑对甬优12籽粒灌浆动态(A)及灌浆速率(B)的影响
Fig. 2. Effect of 3.2%GA3·PP333(LLB) on the grain-filling dynamics (A) and grain-filling rate (B) in rice.

图33.2%赤霉素·多效唑对水稻剑叶叶绿素含量(A)、光合速率(B)及叶片比叶重(C)的影响
Fig. 3. Effect of 3.2%GA3·PP333(LLB) on the chlorophyll content(A), net photosynthetic rate(B) and specific leaf weight(C) flag leaf in rice.

S/F表示强势粒与剑叶可溶性糖含量的比值; I/F表示弱势粒与剑叶可溶性糖含量的比值。
S/F, The ratio of soluble sugar concents in flag leaf to those in superier grains; I/F, The ratio of soluble sugar contents in flag leaf to those in inferier grains.
图43.2%赤霉素·多效唑对水稻剑叶和籽粒可溶性糖含量及其比值的影响
Fig. 4. Effect of 3.2%GA3·PP333(LLB) on soluble sugar contents in flag leaf and grains of rice.

图53.2%赤霉素·多效唑对水稻干物质积累及收获指数的影响
Fig. 5. Effect of 3.2%GA3·PP333(LLB) on the dry matter accumulation and harvest index of rice.
2.5籽粒和剑叶可溶性糖含量及其比值
如图4 所示,LLB处理的强势粒(图4-A)或弱势粒(图4-B)可溶性糖含量均低于对照,而其相对应的剑叶可溶性糖含量则高于对照。相应地,LLB处理的籽粒和剑叶可溶性糖含量比值均显著低于对照,尤其在花后20 d。
2.6干物质量及收获指数
如图5所示,LLB处理的水稻穗干质量在整个灌浆期均高于对照(喷施前10d除外)。此外,LLB处理的茎鞘及叶片干质量在籽粒灌浆前期和中期(LLB喷施后3~33 d)均高于对照,但灌浆后期(LLB喷施后33~53 d)则低于对照。与对照相比,LLB处理的收获指数均有所提高,尤其在喷LLB后43~53 d。
3讨论
水稻结实障碍是影响大穗型超级杂交稻产量潜力发挥的重要因素。前人在这方面进行了大量的研究,也取得很好的成果,3.2%赤霉素·多效唑是其中之一。3.2%赤霉素·多效唑是通过一定比例的GA3和PP333复配而成,可在一定程度减轻亚种间杂交稻结实障碍对产量的影响,主要有以下作用:1)促进抽穗,齐穗期提早2~3 d,并减轻“包颈”现象;2)促进开花,使花期集中,花期缩短约2 d;3)延缓剑叶和其他功能叶片的衰老,可谓“青秆黄熟”;4)调节“粒间顶端优势”,强势粒灌浆势进一步增强,弱势粒灌浆势提高,水稻产量增加[20]。粒粒饱(3.2%赤霉素·多效唑)已经在杂交水稻生产中大面积应用,表现出稳定的增产效果。
本研究结果表明,3.2%赤霉素·多效唑可显著提高超级稻甬优12产量,而结实率和千粒重增加是增产的主要原因(表1),这与前人的研究结果一致[14]。陶龙兴等[14]认为3.2%赤霉素·多效唑可促进水稻抽穗,缩短单株水稻开花时间,提高水稻结实,但未能进一步揭示其作用机制。花粉粒育性、柱头花粉粒数、柱头花粉萌发率及柱头花粉管伸长是直接影响水稻结实的因子,这些因子异常可导致小穗败育。本研究条件下3.2%赤霉素·多效唑提高水稻小穗育性的原因可能与水稻柱头花粉粒的萌发有关,因为LLB处理花粉粒育性及柱头花粉粒数与对照之间的差异不明显,且柱头花粉粒数均在60以上,高于水稻授粉成功所需要的最低花粉粒数(20粒)[21],但LLB处理的柱头花粉萌发率显著高于对照(图1-C、F、J)。水稻授粉后雌蕊组织中的激素含量尤其是IAA的变化,可能是3.2%赤霉素·多效唑提高水稻柱头花粉粒萌发的原因。因为水稻开花授粉过程中,生长素含量大幅提高,如果此时生长素代谢絮乱将会影响花粉粒的萌发及花粉管的伸长,从而导致小穗败育。虽然已有研究表明3.2%赤霉素·多效唑的使用可影响水稻植株中生长素含量的变化,但是否会影响水稻雌蕊组织生长素代谢还未见报道。此外,由于本研究未进行植物激素方面的测定,因而无法证明3.2%赤霉素·多效唑提高柱头花粉粒萌发与植物激素有关,但这也是笔者下一步将要研究的内容。
3.2%赤霉素·多效唑提高水稻小穗育性还可能与水稻开花期光合同化物的供应有关。Kobata等[22]观察到水稻种植密度调整可严重影响水稻结实,与对照相比,降低水稻种植密度可显著提高弱势粒的小穗育性。此外,剪去强势粒同样可以显著增加弱势粒柱头花粉粒数,增幅达1.3倍,表明水稻开花期同化物供应不足会影响水稻授粉过程而导致小穗败育。这与本研究结果比较相似。笔者的研究结果表明3.2%赤霉素·多效唑处理后的叶片、茎鞘及穗干物质质量均高于对照(图5)。3.2%赤霉素·多效唑的这种作用不仅在于有效促进水稻叶片光合的,即LLB处理的剑叶光合速率及叶片比叶重均高于对照(图3),还可能与其提高水稻光合同化物的运转有关,因为LLB处理籽粒及剑叶可溶性糖含量的比值(此值可反映“流”的质量)小于对照(图4)。这说明3.2%赤霉素·多效唑不仅能有效促进水稻光合同化产物的形成,还可提高“流”的质量,使得水稻光合同化产物向小穗运转,从而促进水稻柱头花粉粒的萌发。
大量的研究表明,杂交水稻单穗籽粒的强弱势粒存在“阶梯式灌浆”[23],且亚种间杂交稻比品种间杂交稻的“两段灌浆”现象更明显[24-26]。3.2%赤霉素·多效唑确实可在一定程度上改善亚种间杂交水稻籽粒的灌浆,不仅提高强势粒的粒重,弱势粒的粒重也显著增加,且增幅要明显大于强势粒(表2)。虽然GA3和PP333的作用行为完全不同,且PP333是GA3生物合成的抑制剂,但研究表明GA3和PP333均可调节大穗型杂交水稻的“粒间顶端优势”[27],且外源PP333对外源GA3并无拮抗作用。陶龙兴[15]等认为3.2%赤霉素·多效唑调节“粒间顶端优势”的作用途径包括两个方面:1)使强势粒的灌浆优势得到一定程度的抑制——灌浆峰值降低;2)弱势粒的灌浆得到改善——灌浆势启动值提高,峰期提前,灌浆势下降时间推迟。虽然笔者的研究结果也表明3.2%赤霉素·多效唑可促进水稻籽粒灌浆,其平均灌浆速率高于对照,但同时也观察到3.2%赤霉素·多效唑出现抑制强势粒及弱势粒最高灌浆速率的现象,类似于PPP333单独使用的作用效果。王熹等[27]的研究结果表明PPP333对单穗强弱势粒的灌浆势的“顶端优势”起抑制作用,虽然强势粒的最大灌浆速率仍然高于弱势粒,但它们之间的差距变小。这从另一方面也说明3.2%赤霉素·多效唑提高水稻籽粒灌浆速率主要在于延缓籽粒灌浆中后期籽粒灌浆速率的下降。对此,王熹等认为可能与3.2%赤霉素·多效唑可延缓水稻叶片和籽粒的衰老有关,其研究结果表明3.2%赤霉素·多效唑处理的剑叶叶绿素含量及净光合速率均显著高于对照,即所谓的“青秆黄熟”。这一点与本研究结果一致,笔者在研究中观察到3.2%赤霉素·多效唑处理的叶绿素含量、光合速率及叶片比叶重在整个灌浆期均高于对照(图3)。而在籽粒关键酶活性方面,杨建昌等[28]认为水稻弱势粒籽粒充实不足主要在于籽粒灌浆关键酶活性的限制,而不在于光合产物的不足,因而3.2%赤霉素·多效唑极有可能是通过促进籽粒灌浆关键酶活性延缓籽粒衰老的。虽然籽粒灌浆关键酶活性在本研究中没有测定,但从籽粒和叶片可溶性糖含量比值推断(图4),LLB处理的籽粒灌浆关键酶活性可能高于对照。
此外,笔者还观察到超级稻甬优12强势粒和弱势粒灌浆速率达峰值的时间比较一致,均在花后12 d左右,与前人的研究稍有不同[23-26]。但同时,笔者也注意到强势粒灌浆速率在花后12 d开始大幅度下降,而弱势粒则在花后18 d。这可能与水稻品种的灌浆特性有关:1)超级稻甬优12弱势粒灌浆速率在花后12 d和18 d差异并不大,分别为0.9999mg·grain-1day-1和0.7990 mg·grain-1d-1,而在强势粒上则分别为1.3424 mg·grain-1d-1和0.5964 mg·grain-1d-1;2)超级稻甬优12每穗粒数较多,平均达450粒(表1)。据笔者的观察,该组合灌浆中后期强势粒和弱势粒成熟度较高,反而很多中间类型籽粒还存在灌浆不充分的现象,由于类似现象还未见报道,因而是否与强势粒和弱势粒灌浆同步的现象有关还有待研究。
总之,抽穗期使用3.2%赤霉素·多效唑可显著提高超级稻甬优12的产量,增产的原因在于结实率和千粒重的增加。3.2%赤霉素·多效唑能够显著提高水稻结实率,主要在于此期喷施3.2%赤霉素·多效唑不仅提高水稻光合同化物的形成,还能促进水稻光合同化物向籽粒运转,充足的同化物供应有利于水稻柱头花粉粒的萌发。3.2%赤霉素·多效唑延缓水稻叶片衰老是千粒重增加的主要原因。
参考文献:
[1]谢华安. 华南型超级稻育种及其技术研究进展. 沈阳农业大学学报, 2007, 38(5): 714-718.
Xie H A. Advances on the breeding program and technology research in South China type super-rice.JShengyangAgricUniv, 2007, 38(5): 714-718. (in Chinese with English abstract)
[2]苏泽胜, 李泽福. 安徽省超级稻研究与应用现状及展望. 沈阳农业大学学报, 2007, 38(5): 739-743.
Su Z S,Li Z F.Present status and prospect of super-rice research and utilization in Anhui Province.JShengyangAgricUniv, 2007, 38(5): 739-743. (in Chinese with English abstract)
[3]杜士云, 王守海, 李成荃, 等. 超级稻育种进展及存在的问题. 中国农学通报, 2006, 22(8): 195-198.
Du S Y,Wang S H,Li C Q,et al. Introduction to the progress and problems on super rice breeding.ChinAgricSciBull, 2006,22(8):195-198. (in Chinese with English abstract)
[4]敖和军, 王淑红, 邹应斌, 等. 超级杂交稻干物质生产特点与产量稳定性研究. 中国农业科学, 2008, 41(7): 1927-1936.
Ao H J,Wang S H,Zhou Y B,et al.Study on yield stability and dry matter characteristics of super hybrid rice.SciAgricSin, 2008,41(7):1927-1936. (in Chinese with English abstract)
[5]Mohapatra P K, Patel R, Sahu S K. Time of flowering affects grain quality and spikelet partitioning within the rice panicle.AustrJPlantPhysiol, 1993, 20(2): 231-241.
[6]Yang J C, Peng S B, Visperas R M, et al. Grain filling pattern and cytokine in content in the grains and roots of rice plants.PlantGrowthReg, 2000, 30( 3): 261-270.
[7]吕川根, 谷福林, 陆曼丽, 等. 五个水稻籼粳杂种一代与其双亲生物学特性的比较研究. 中国水稻科学, 1993, 7(4):205-210.
Lv C G,Gu F L,Lu M L,etal.The comparative studies on biological characteristics between five Indica-Japonica hybrids and their parents.ChinJRiceSci, 1993,7(4):205-210. (in Chinese with English abstract)
[8]徐秋生, 李卓吾. 亚种间杂交稻谷粒灌浆特性与籽粒充实度的研究. 杂交水稻, 1994, 2: 26-29.
Xu Q S,Li Z W.Studies on the filling characteristics and the plumpness of grains in Inter-subspecific hybrid rice.HybridRice, 1994,(2):26-29. (in Chinese with English abstract)
[9]Yang J C, Zhang J H, Wang Z Q, et al. Hormones in the grains in relation to sink strength and post anthesis development of spikelets in rice.PlantGrowthReg, 2003, 41: 185-195.
[10]陶龙兴, 王熹, 黄效林, 等. 内源IAA对杂交稻强、弱势粒灌浆增重的影响. 中国水稻科学, 2003, 17(2): 149-155.
Tao L X,Wang X,Huang X L,etal.Effects of endogenous IAA on grain filling of hybrid rice.ChinJRiceSci, 2003,17(2):149-155. (in Chinese with English abstract)
[11]王熹, 田淑兰. IAA与亚种间杂交稻籽粒发育的关系及烯效唑的调节. 中国水稻科学, 1998, 12(2): 99-104.
Wang X,Tao L X,Xu R S,etal.Apical-grain superiority in hybrid rice.ActaAgronSin, 2001,27(6):980-985. (in Chinese with English abstract)
[12]Ishimaru T, Hirose T, Matsuda T, et al. Expression patterns of genes encoding carbohydrate metabolizing enzymes and their relationship to grain filling in rice (OryzasativaL): Comparison of caryopses located at different positions in a panicle.PlantCellPhysiol, 2005, 46: 620-628.
[13]杨卫兵, 王振林, 尹燕枰, 等. 外源ABA和GA对小麦籽粒内源激素含量及其灌浆进程的影响. 中国农业科学, 2011, 44(13): 2673-2682.
Yang W B,Wang Z L,Yin Y P,etal.Effects of spraying exogenous ABA or GA on the endogenous hormones concentration and filling of wheat grains.SciAgricSin, 2011,44(13):2673-2682. (in Chinese with English abstract)
[14]陶龙兴, 黄效林, 王熹. 粒粒饱对水稻的生物效应. 中国水稻科学, 2000, 14(2): 93-97.
Tao L X,Huang X L,Wang X.Biological effects of lilibao on rice plant.ChinJRiceSci, 2000,14(2):93-97. (in Chinese with English abstract)
[15]陶龙兴, 黄效林, 俞美玉, 等. “粒粒饱”的生理作用及水稻化控增产技术. 植物保护与植物营养研究进展, 1999: 375-378.
Tao L X,Huang X L,Yu M Y,etal.Biological Effects of Lilibao and technique of increased yield by chemical control.DOI: 10.16819/j.1001-7216.2016.5000AdvaPlantProPlantNutri. 1999:375-378.(in Chinese)
[16]张镇铭, 俞祥群, 邵达孚,等. 水稻不同生育阶段诱施用粒粒饱的生物学效应及增产效果: 中国植物生理学会植物生长发育与信息转导学术会议. 厦门:中国植物生理学会,1999:68.
Zhang Z M,Yu X Q,Shao D F,et al. Biological effects and yield-increasing effect of Lilibao on rice at different growth stage: Academic Conference on Plant Growth and Development and Information Transfer in the National Congress of Plant Physiology. Xiamen: CSPB, 1999: 68. (in Chinese)
[17]童继平, 吴跃进, 刘学军. 粒粒饱对水稻品种后期活性及抗折力的影响. 安徽农业科学, 2003, 3(6): 1077-1088.
Tong J P,Wu Y J,Liu X J. Effect of plant growth regulator on rice growth in later period.JAnhuiAgricSci, 2003, 3(6):1077-1088. (in Chinese with English abstract)
[18]朱庆森, 曹显祖, 骆亦奇. 水稻籽粒灌浆的生长分析. 作物学报, 1988, 14(3): 182-193.
Zhu Q S,Cao X Z,Luo Y Q.Analysis of rice grain filling.ActaAgronSin, 1988,14(3):182-193. (in Chinese with English abstract)
[19] Richards F J. A flexible growth function for empirical use.JExperBot, 1959, 10: 290-300.
[20]王熹, 陶龙兴, 俞美玉. GA3对杂交稻“粒间顶端优势”及灌浆期间籽粒内源IAA的影响. 植物生理学报, 2000, 26(3): 247-251.
Wang X,Tao L X,Yu M Y.Effect of exogenous GA3 on "Inter-grain apical dominance" and endogenous IAA in grain filling stage of hybrid rice.ActaPhytophysiolSin, 2000,26(3):247-251. (in Chinese with English abstract)
[21]Satake T, Yoshida S. High temperature-induced sterility in indica rice at flowering.JpnJCropSci, 1978, 47: 6-17.
[22]Kobata T, Yoshida H, Masiko U. Spikelet sterility is associated with a lack of assimilate in high-spikelet-number rice.CropEcolPhysiol, 2013, 105(6): 1821-1831.
[23]顾自奋, 朱庆森, 曹显祖. 水稻结实率的研究——稻穗上强弱势粒的干重积累过程与空秕粒的分布. 中国农业科学, 1981, 23(6): 38-42.
Gu Z F,Zhu Q S,Cao X Z.Studies on the grain ripenning rate of rice: The relationship between the accumulation of dry weight in vigorous and weak spikelets and the distribution of sterile,abortive kernsel.SciAgricSin, 1981,23(6):38-42. (in Chinese with English abstract)
[24]邓仲篪, 周鹏, 陈翠莲. 籼粳亚种杂交组合的结实率与光合产物供给水平及转运效率间的关系. 华中农业大学学报, 1993, 12(4): 333-338.
Deng Z C,Zhou P,Chen C L.Relationship between seed setting ratio and photosynthetic capacity and transport of substances in the Indica-Japonica intersubspecific hybrid rice.JHuazhongAgricUniv, 1993,12(4):333-338. (in Chinese with English abstract)
[25]朱庆森, 王志琴, 张祖建, 等. 水稻籽粒充实程度的指标研究. 江苏农业科学, 1979, 30(3): 52-59.
Zhu Q S,Wang Z Q,Zhang Z J,etal.Study on indicators of grain-filling of rice.JJiangsuAgricColl, 1979,30(3):52-59. (in Chinese with English abstract)
[26]周建林, 陈良碧, 周广洽. 亚种间杂交稻籽粒充实动态及生理研究. 杂交水稻, 1992, (5): 36-40.
Zhou J L,Chen L B,Zhou G Q.A Study on the dynamic state and physiology of grain filling of subspecific hybrid rice.HybridRice,1992,(5):36-40. (in Chinese with English abstract)
[27]王熹, 陶龙兴, 黄效林. 赤霉酸和多效唑对水稻粒间顶端优势的调节及对产量的影响. 中国水稻科学, 1999, 13(4): 217-222.
Wang X,Tao L X,Huang X L.Regulating the inter-grain apical dominance of rice by GA3 and PP333.ChinJRiceSci, 1999,13(4): 217-222. (in Chinese with English abstract)
[28]杨建昌. 水稻弱势粒灌浆机理与调控途径. 作物学报, 2010, 36(12): 2011-2019.
Yang J C.Mechanism and regulation in the filling of inferior spikelets of rice.ActaAgronSin, 2010, 36(12):2011-2019. (in Chinese with English abstract)
Mechanism Behind the Effects of GA3and PP333on Grain Yield Formation of Super Rice Yongyou 12
YANG Xue-qin#, CHEN Ting-ting#, ZHAO Xia, ZHANG Cai-xia, YANG Yong-jie, FU Guan-fu,TAO Long-xing*
(StateKeyLaboratoryofRiceBiology,ChinaNationalRiceResearchInstitute,Hangzhou310006,China;#These authors contributed equally to this work;*Corresponding auther, E-mail: lxtao@mail.hz.zj.cn)
YANG Xueqin, CHEN Tingting, ZHAO Xia, et al. Mechanism behind the effects of GA3and PP333on grain yield formation of super rice Yongyou 12. Chin J Rice Sci, 2016, 30(1): 53-61.
Abstract:In order to improve spikelet fertility and grain weight of inferior spikelets in super rice, 3.2%GA3·PP(333) (Lilibao,LLB) were sprayed on super rice Yongyou 12 three days after heading. We investigated the effect of LLB on grain yield and the underlying mechanisms. The results indicated that LLB increased the grain yield by 4.41% and 4.31% in 2012 and 2013, respectively compared to corresponding controls, while the theoretical yield showed 9.21% and 6.26% increment. This may be ascribed to increased seed-setting rate and grain weight under LLB treatment compared with the control. It had been observed that there was no significant difference in pollen viability and pollen numbers on stigma between the LLB treatment and control, while the LLB treatment significantly increased the pollen germination on stigma, which was responsible for the improvement in seed-setting rate. Additionally, the chlorophyll content, net photosynthetic rate and specific leaf weight of flag leaf in the LLB treatment were higher, while the ratios of soluble sugar content in leaf to grain were lower than those of the control in the late grain filling period, indicating that LLB could not only delay the leaf senescence but also promote transportation of assimilates to grains.
Key words:3.2%GA3·PP(333); super rice; grain yield; spikelet fertility; 1000-grain weight
文章编号:1001-7216(2016)01-0053-09
中图分类号:S482.8; S511.01
文献标识码:A
基金项目:国家自然科学基金资助项目(31501264;31101116);浙江省自然科学基金资助项目(LQ15C130003);公益性行业(农业)科研专项(201203029);国家水稻产业体系项目(CARS-01-27);比尔和梅琳达盖茨基金资助项目;中央级公益性科研院所专项资金资助项目(2014RG004-4)。
收稿日期:2014-11-26; 修改稿收到日期: 2015-05-18。
