局部遮光对鹅绒委陵菜基株形态塑性及生物量配置的影响
2016-04-27樊星蔡捡刘金平李莹张小晶曾晓琳
樊星,蔡捡,刘金平 ,李莹,张小晶,曾晓琳
(西华师范大学西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
局部遮光对鹅绒委陵菜基株形态塑性及生物量配置的影响
樊星,蔡捡,刘金平*,李莹,张小晶,曾晓琳
(西华师范大学西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009)
摘要:设置基株、母株、若干克隆片段被分别局部遮光处理,通过测定根、茎、叶的数量性状和生物量,分析局部遮光对鹅绒委陵菜母株、子株、基株形态塑性及生物量配置的影响。结果表明,1)局部遮光对母株株高、叶长、叶宽、叶面积及根系深度有显著影响(P<0.05),局部遮光均可提高母株的株高、叶片长度和总叶面积,不同遮光部位或遮光强度对母株叶宽、单叶面积和根长的影响不同;2)局部遮光对子株叶长、叶面积有显著影响,克隆片段遮光是影响子株叶片大小的主要因素,母株遮光次之;3)局部遮光对克隆片段子株数和新增子株数有显著影响(P<0.05),局部遮光可显著增加基株的克隆片段数(P<0.05),母株遮光对匍匐茎伸长和新克隆片段形成有重要的影响;4)局部遮光对基株根、茎、叶含水量及叶和根生物量累积有极显著影响(P<0.01),遮2个以上克隆片段或遮母株可显著增加基株根生物量累积,但基株完全遮光使根生物量显著低于CK;5)局部遮光对基株茎、叶、根生物量分配比均有极显著影响(P<0.01),遮光对叶分配比影响最大,对茎分配比次之,对根分配比较小;6)局部遮光对基株地上生物量比有极显著影响(P<0.01),遮光部位对基株茎根比和茎叶比有极显著影响(P<0.01),全遮光显著增加基株的茎根比和茎叶比。综上所述,局部遮光对克隆植物鹅绒委陵菜的母株、子株、克隆片段和基株形态塑性和生物量配置有直接的影响。
关键词:鹅绒委陵菜;克隆植物;遮光;形态可塑性;生物量配置
克隆植物最初母株(mother ramet)是由合子(种子)萌发形成的实生苗(seedling)。经母株克隆生长形成的后代分株(descendant ramet)或子代分株(简称子株,daughter ramet) 由匍匐茎或根状茎彼此相连,构成了克隆片段(clonal fragment)或分株系统(ramet system)。克隆植物具有分株、克隆片段和基株3个层次的等级结构系统特征[1]。诸多研究表明,克隆植物具有极强的表型塑性和生理整合潜力,从而提高对资源的利用效率[2]。不同克隆植物对水分、光合产物、生理活性物质的生理整合表现出明显的差异[2],致使不同植物的适应性、抗逆性及生存能力不同。地理分布狭窄、生存条件恶劣的草本克隆植物,不仅生活史远低于木本,且抵御胁迫的结构基础与物质基础较为简单,要求其能依据生境胁迫种类,快速、有效、系统地进行相应的生长策略调整,顺利完成开花结实,才能不被自然选择淘汰,使物种得以存活和延续。
鹅绒委陵菜(Potentillaanserina)属蔷薇科(Rosaceae)委陵菜属(Potentilla),又名曲尖委陵菜、人参果、延寿草、蕨麻[3],是典型的高寒地区多年生匍匐茎型莲座状草本植物[4-5],具有游击型(guerilla)克隆生长习性[6]。近年来,对其药用价值、食用价值[7]、饲用价值[8]及景观价值[9-10]进行了大量研究,对其克隆生长特性[11]、空间拓展性[12]、生物学特性[13]、无性系结构[4,14]、繁殖分配[15]、匍匐茎扩散模式[14]、克隆分株的风险分摊和存活调控[16]进行了较为深入研究。关于生境胁迫对其生长、生理及生殖过程的形态塑性及生理整合研究较少,只对不同光照下的形态塑性[17]、不同土壤养分和水分下的生长特征[18]进行了探讨。鹅绒委陵菜植株呈粗网状紧贴地表的匍匐生长习性,使其常处于植物群落的底层,分株、克隆片段和基株极易处于不同的遮光条件。遮光导致分株间光合产物的转移格局发生改变,从而改变原有的整合单位和格局[19]。目前,此类研究多在人工控制条件的实验室、温室进行,其结果难以准确反映该类植物在自然环境下生理整合的基本特性[20]。且关于局部遮光对鹅绒委陵菜形态塑性及生物量配置研究极少。
本研究采用野生群落人工干扰的方法,设置基株、母株、克隆片段3个等级结构被局部遮光,通过测定构件数量性状变化及生物量变化,分析鹅绒委陵菜形态塑性及生物量分配的调节能力,研究母株、分株、基株遮光下鹅绒委陵菜的生长策略差异。旨在通过异质性光照条件对其生物量格局及形态可塑性的影响,探讨适合其生长发育的局部光照条件,为鹅绒委陵菜人工栽培时光照设置及养护技术奠定基础,为野生植物资源的开发、利用及保护提供依据。
1材料与方法
1.1试验地概况
试验于阿坝州红原县邛溪镇二农场进行,该地为大陆性高原温带季风气候[21],东经102°32′,北纬32°46,海拔3497 m,年均温1.1℃,极端高温23.5℃,极端低温-33.8℃,年降水量738 mm,相对湿度71%,≥10℃年积温仅865℃,年总辐射量为20.93×106~29.30×106kJ/m2。土壤为草甸土,0~20 cm土壤的有效氮、有效磷、有效钾含量分别为276 mg/kg,10.2 mg/kg,131 mg/kg,有机质含量5.87%,pH值5.91。
1.2试验材料
于2014年7月,在老芒麦(Elymussibiricus)种子休闲地中,剔除共生植物,形成生境基本一致的鹅绒委陵菜种群。选株高(4.0±0.5) cm、具3根长(19.0±1.0) cm匍匐茎,且匍匐茎上子株大小(叶长2.0±0.2 cm)及数量(2个)相似的当年生野生委陵菜基株为材料。
1.3试验设计
随机3基株为1处理组,设A、B、C、D、E及CK共6组,每组设3次重复。于7月5日,记录每株形态参数后,用遮荫度80%的黑色网进行遮光处理,遮光高度为(5.0±0.5) cm,1条匍匐茎记为1个克隆片段,A组遮1片段,B组遮2片段,C组遮3片段,D组遮母株,E组遮基株(整株),CK组不遮光。遮光处理用1表示,未遮光用0表示。遮光后不进行相关养护,不考虑遮光对微环境的影响。处理40 d后(8月15日)测定如下参数。
1.4观测项目及测定方法
叶:遮光处理前,在母株(子株)上选大小相似叶片进行标定,测定处理后母株(子株)叶数、叶长、叶宽、单叶面积(系数法)及总叶面积;基株总叶面积为母株叶面积与子株叶面积的和。
茎:测定处理前后母株株高、克隆片段的匍匐茎长;测定处理后子株数,新增子株数为处理后子株数减处理前子株数;测定新增克隆片段数。
根:挖取、清洗、拭干后,测定母株的最大根长。
生物量:将每组3基株分为叶、茎、根3部分,称鲜重后,装入纸袋,在105℃下烘至恒重,称量干重为生物量。计算:含水量=(鲜重-干重)/鲜重×100%;分配比=某部分生物量/总生物量×100%;比叶面积=总叶面积(基株)/叶生物量(基株);比茎长=总茎长(基株)/茎生物量(基株);地上生物量比率=地上生物量/总生物量×100%;茎根比=地上生物量/地下生物量;茎叶比=茎生物量/叶生物量。
1.5数据处理
用SPSS 19.0进行多重比较等数据统计与分析。
2结果与分析
2.1局部遮光对母株形态特征的影响
局部遮光对母株株高有极显著影响(P<0.01) (表1),除E处理外,均显著提高了母株株高(P<0.05)。局部遮光对母株叶数无显著影响(P>0.05),但对叶长、叶宽、单叶面积有显著影响(P<0.05)。处理间母株叶宽差异显著,但均与CK无显著差异。B、D处理母株叶长、单叶面积显著高于CK。局部遮光对母株总叶面积有极显著影响(P<0.01),D处理的总叶面积显著高于其他处理,近为CK的2倍,遮2个克隆片段B处理的总叶面积显著高于遮1(A)、遮3(C)处理。局部遮光对母株根长有极显著的影响(P<0.01),克隆片段全部遮光C处理根长显著大于其他处理,而A、B、D下根长显著小于CK。可见,任何局部遮光均可提高母株的株高、叶片长度和总叶面积,不同遮光部位或遮光强度对母株叶宽、单叶面积和根长的影响不同。
2.2局部遮光对克隆片段形态特征的影响
2.2.1对子株的叶性状影响局部遮光对子株叶长、单叶面积、总叶面积有极显著影响(P<0.01),对子株叶宽无显著影响(P>0.05)(表2)。局部遮光均增大子株的叶长、叶宽和叶面积。A、B被遮克隆片段子株叶长、叶宽、叶面积均大于未遮片段。D处理的子株叶长、叶宽、叶面积显著高于CK(P<0.05);A0和C1处理,子株叶长、叶面积显著小于其他处理,但显著大于CK(P<0.05)。综上所述,局部遮光使子株单叶面积和总叶面积均显著大于CK,大小顺序为B1>A1>B0>D0>A0>E1>C1>CK,克隆片段遮光是影响叶片大小的主要因素,母株遮光次之。

表1 遮光对鹅绒委陵菜母株的形态特征的影响
注:同列不同小写字母间差异显著(P<0.05),下同。
Note: Values in a same column with different lowercase letters indicate significant differences (P<0.05). The same below.
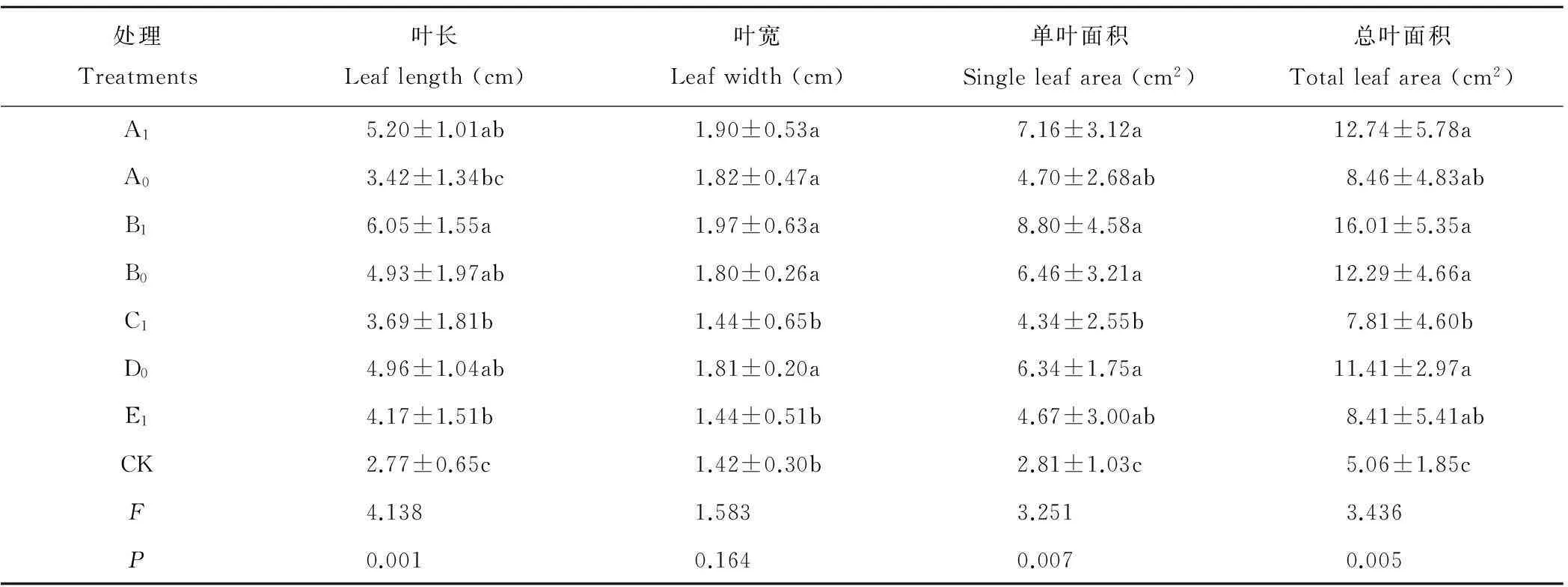
表2 遮光对鹅绒委陵菜子株叶形态特征的影响
2.2.2对子株数及匍匐茎影响局部遮光对克隆片段子株数和新增子株数有显著影响(P<0.05)(表3),A1的子株数最大,A0与CK无显著差异,B0子株数显著低于CK,其他遮光处理对子株数影响不显著。局部遮光对克隆片段匍匐茎长有极显著影响(P<0.01)。A1、B1匍匐茎长与CK无显著差异,A0、B0匍匐茎显著低于CK(P<0.05)。D处理显著促进了克隆片段匍匐茎伸长(P<0.05),而C、E处理对匍匐茎长度几无影响。局部遮光均可显著增加基株的克隆片段数(P<0.05), D处理新增克隆片段数显著大于其他处理,3个遮光克隆片段处理间差异较小。可见,母株遮光对匍匐茎伸长和新克隆片段形成有重要的影响,但基株全部遮光与子株遮光对形成新克隆片段的影响差异极小。
2.3局部遮光对基株形态特征的影响
局部遮光对基株总叶面积有极显著的影响(P<0.01)(表4),遮光处理基株总叶面积均大于CK,B、D处理基株总叶面积显著高于其他处理(P<0.05),尤其D处理总叶面积近为CK的2 倍,A、C、E处理与CK差异不显著。局部遮光对基株比叶面积有显著影响(P<0.05),基株遮光的比叶面积显著大于其他处理,而遮母株、遮子株虽对叶面积有显著影响,但比叶面积间差异较小。从A、B、C、D比叶面积与CK无差异,可见局部遮光不仅影响叶面积,也影响单位叶面积的质量。

表3 遮光对鹅绒委陵菜子株数及匍匐茎形态特征的影响
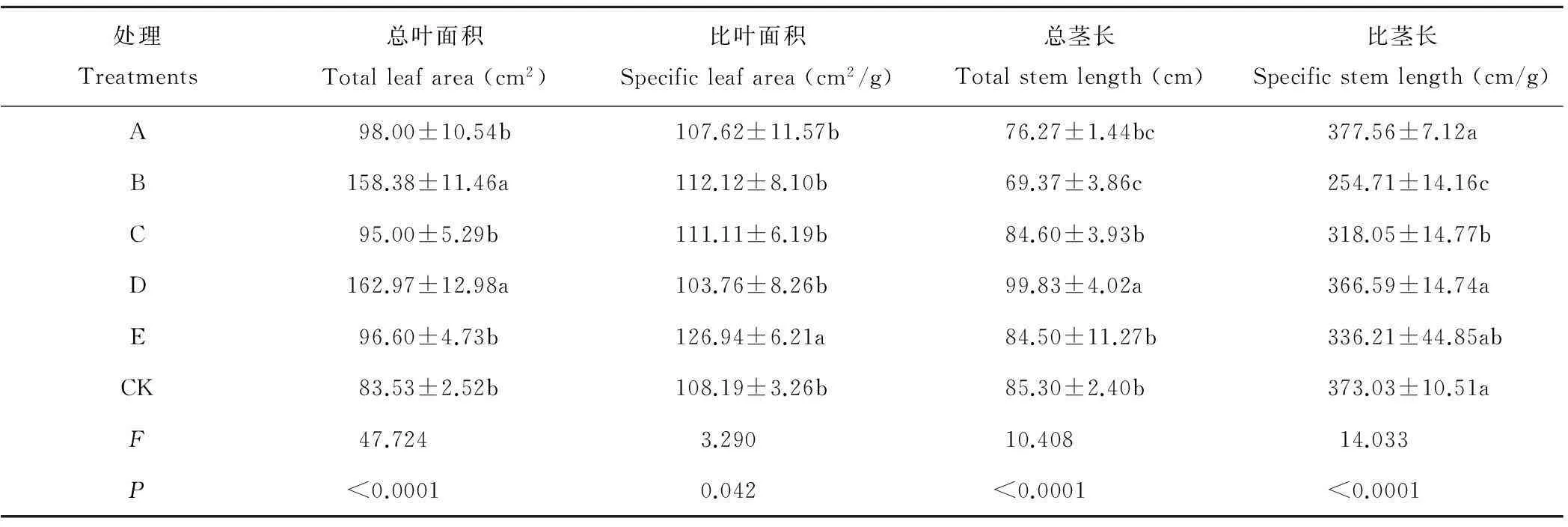
表4 遮光对鹅绒委陵菜基株形态特征的影响
局部遮光对基株总茎长和比茎长有极显著的影响(P<0.01)(表4)。D处理基株的总茎长显著高于其他处理(P<0.05),B处理总茎长显著低于C、D、E、CK处理(P<0.05),A、C、E处理与CK差异较小。B、C处理的比茎长显著低于其他处理,而A、D、E处理比茎长与CK无显著差异。可见,遮2个以上克隆片段显著影响基株匍匐茎的长度与粗细度;遮母株显著增加基株茎长,但对茎粗细无影响;遮基株对茎性状影响较小。
2.4局部遮光对基株含水量的影响
局部遮光对基株叶、茎、根含水量均有极显著影响(P<0.01)(表5),受影响顺序为根>茎>叶。遮光克隆片段越多,茎、叶含水量越高;D处理的叶含水量显著高于CK(P<0.05),而茎含水量与CK无显著差异;E处理茎叶含水量最高且显著高于CK(P<0.05)。被遮克隆片段数量显著影响根系含水量,但仅B处理显著高于CK;遮母株对根含水量影响较小;遮基株E处理的根含水量显著低于其他处理。可见遮光部位对根系吸水能力及活力有显著影响。
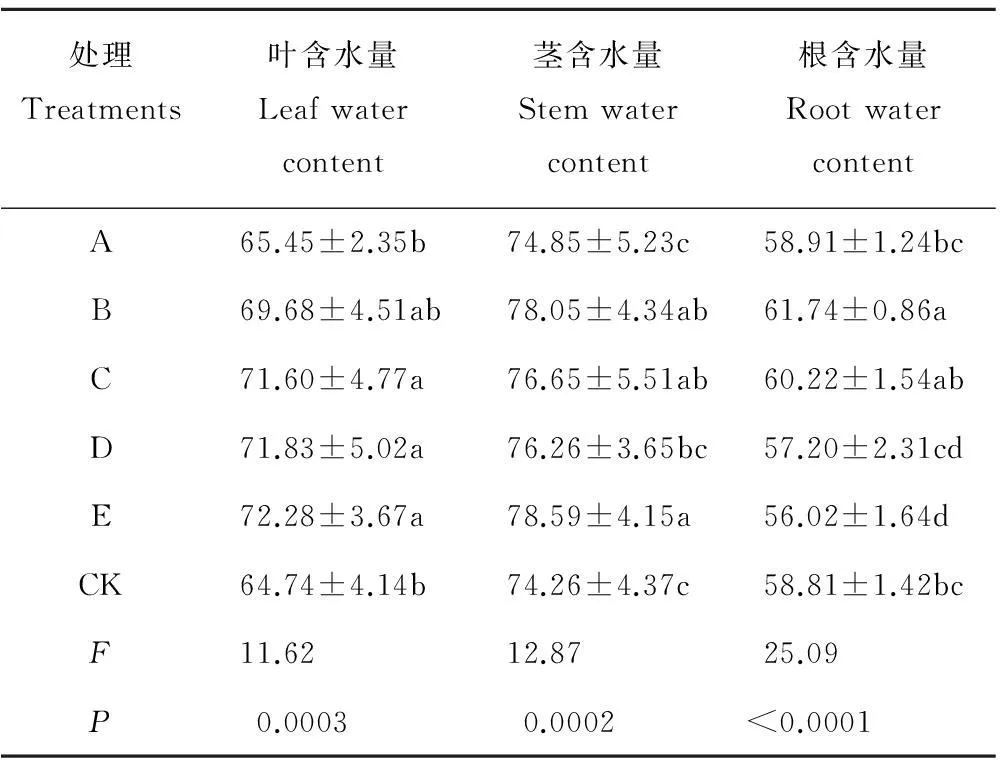
表5 遮光对鹅绒委陵菜各构件含水量的影响
2.5局部遮光对基株生物量的影响
2.5.1对基株生物量累积的影响局部遮光对基株叶、根生物量累积有极显著影响(P<0.01)(表6),对基株茎生物量累积无显著影响。各遮光处理茎生物量与CK无显著差异(P>0.05),但A处理茎生物量显著低于B、C、D处理(P<0.05)。B、D处理显著增加了基株叶生物量(P<0.05),而其他处理与CK无显著差异。遮2个以上克隆片段或遮母株可显著增加基株根生物量累积,但完全遮光E处理的根生物量显著低于CK。局部遮光对基株总生物量累积有极显著影响(P<0.01),B、D处理显著增加了基株生物量累积(P<0.05),而其他处理与CK均无显著差异。
2.5.2对基株生物量配置的影响局部遮光对基株茎、叶、根生物量分配比均有极显著影响(P<0.01)(表7),由F值可见,局部遮光对叶分配比影响最大,对茎分配比次之,对根分配比最少。多重比较说明,E处理显著增加了基株的茎分配比;B、D处理显著减少了茎分配比,而增加了叶分配比;C处理显著降低叶分配比而增加了根分配比。
局部遮光对基株地上生物量比有极显著影响(P<0.01)(表7),克隆片段全部被遮光的C处理地上生物量比显著低于其他处理,但其他遮光处理几乎没有改变基株地上、地下生物量的分配比率。进一步分析茎根比说明,遮光部位对茎根比有极显著影响(P<0.01),A、E处理显著增加、C处理显著减少了茎根比。局部遮光对基株茎叶比有显著影响,遮母株D处理显著降低了茎叶比,而全遮光的E处理茎叶比最大。

表6 遮光对鹅绒委陵菜各构件生物量的影响

表7 遮光对鹅绒委陵菜生物量配置的影响
3讨论与结论
光是植物一切生命活动的能量原初来源,光合作用所合成的有机物质是植物生长、发育的物质基础;同时光也为植物光形态建成提供信号,诱导、促进、调控植物的生长、发育和分化等过程。克隆植物具有分株、克隆片段和基株3个层次的等级结构系统特征,具有游击型(guerilla)生长习性的克隆植物对光照具有极强的表型塑性。诸多学者对异质光照下的克隆植物的可塑性进行了研究,但对不同植物研究结果不尽相同,如对蛇莓(Duchesneaindica)[22]和金戴戴(Halerpestesruthenica)[23]研究发现随光强减弱,间隔子长度、分枝强度、基株株长、茎节长度增大,但对喜旱莲子草(Alternantheraphiloxerxides)研究结果为随光强度下降而下降[24],或许是植物种间差异、光照梯度设置差异和处理时间差异共同作用造成研究结果不同。现有研究多通过不同光照强度对基株处理进行可塑性研究。本试验通过对分株、克隆片段和基株进行局部遮光处理表明,鹅绒委陵菜具有极强的形态可塑性,母株、子株、基株的根、茎、叶性状,新增子株及新增克隆片段数均能对局部遮光产生相应的形态反应,但对不同部位遮光间存在着一定的差异,遮母株对基株形态影响最大、遮克隆片段影响次之、遮基株影响最小。无论哪种局部遮光均增加了母株高度,且在叶片数保持相对稳定的基础上,均通过增加叶长或叶宽,增大了单叶面积,从而增加了基株的总叶面积,提高了整个植株的光合叶面积基础。母株被遮光时,基株叶面积和茎长的增加量显著高于其他处理。被遮光克隆片段越多,基株叶面积越小而匍匐茎长越大,克隆片段遮光对增加子株数和茎长有明显影响。鹅绒委陵菜对局部遮光的形态塑性,提高了基株对光胁迫的适应性。
常通过构件生物量累积量和分配比,分析植物对生境胁迫的响应能力,及为减少胁迫造成损害,植物在各功能间权衡关系。遮光降低了光照强度,植物对光照强度的形态塑性,导致构件生物量累积量和分配比的变化。喜旱莲子草的总生物量及根、茎、叶生物量均随光照强度下降显著减少[24],过路黄(Lysimachiachristinae)叶生物量、根生物量以及总生物量随高度遮光而严重下降[25],矮竹(Shibataeachinensis)的分株生物量在遮荫环境比开阔环境明显下降[26],结缕草(Zoysiajaponica)克隆植株的总分株生物量、分株叶和根生物量随光照强度减弱显著减小[27]。本试验中,遮光对鹅绒委陵菜基株的叶生物量、根生物量及总生物量有极显著影响,但无论对基株、母株或克隆片段遮光均没有降低基株茎、叶生物量和总生物量,反而B、D处理显著增加了基株的叶生物量、根生物量和总生物量,仅完全遮光(E)下基株根生物量显著下降。遮光对鹅绒委陵菜生物量累积的影响与上述几种克隆植物研究结果相反,或因鹅绒委陵菜具有极强的“补偿”生长特点,局部遮光加强了基株的光形态建成信号,使分株、母株、基株的叶面积均增大,光合同化能力加强,合成更多的物质,导致其生物量在遮光下反而增加。
一般认为在弱光下,基株常将生物量优先向地上部分分配,利于摄取更多的光照资源。诸多试验证实了这个假设,如结缕草在中度和深度遮荫下,叶生物量比均高于根生物量比和茎生物量比,随光照强度增大,叶生物量比逐渐降低,茎生物量比逐渐升高[28];又如马蹄金(Dichondrarepens)通过增加叶重比,降低分枝数,减少光合产物向根部分配等措施来维持其生长和适应遮光环境[29]。本试验中,局部遮光对鹅绒委陵菜叶分配比影响最大,对茎分配比次之,对根分配比最少。局部遮光对基株地上生物量比有极显著影响,但遮光不同部位产生的影响差异明显,E处理显著增加了基株的茎分配比,D处理显著减少了茎分配比而增加叶分配比,C处理显著降低叶分配比而增加了根分配比。可见,鹅绒委陵菜具有极完善的能量分配策略,当基株全部被遮光时,增加可帮助逃离荫蔽环境的匍匐茎生物量分配,当母株被遮光则增加叶分配比、增加光合能力来降低遮光造成的影响,当所有克隆片段被遮光限制空间拓展时,则降低地上生物量分配比,增加根分配比来保证根系健康,以增大基株的生存几率。
局部遮光对克隆植物鹅绒委陵菜的母株、子株、克隆片段和基株形态塑性和生物量配置有直接的影响,且遮光部位不同造成的影响差异很大。所以,开展光照对克隆植物可塑性、生物量配置及生殖策略等相关研究时,不仅要考虑光照强度,也要注意光照部位、光照时间对植物的影响。同时植株生长阶段、发育状况对研究结果也有影响,如鹅绒委陵菜的匍匐茎数目,对无性系中分株数、间隔子数、茎总长和匍匐茎比节间重,对用于繁殖的匍匐茎和分株干重比例均有影响[11]。鹅绒委陵菜基株、分株和分株的生物量配置对生境资源有高度适应性[12],对克隆片段遮光并未对其母株产生负影响,反而生长更好,表明鹅绒委陵菜具有顶向运输为主的克隆整合功能。基株中被遮光克隆片段的叶面积、茎长、子株数等指标均大于未遮光克隆片段,表明局部遮光改变了基株的整合格局,母株将更多的营养物质输送给被遮荫的克隆片段,与董鸣[1]提到的克隆整合对整个克隆片段的生长具有显著收益观点一致。生理整合使物质或资源在克隆分株之间进行运输与分享,使克隆植物具有良好的适应能力和对不良条件的抵抗力[2],甚至在异质营养条件下的生长状况好于同质营养条件[30]。整合作用使克隆分株对生境产生局部反应和非局部反应,两种反应共同影响着克隆植物的生长和繁殖,进而影响基株和分株的适合度[31]。但很多研究表明,生理整合对克隆植物的形态特征修饰效果有正效应,也有负效应[32]。关于鹅绒委陵菜生理整合能力,以及生理整合对形态可塑性、构件等级结构、种群调节能力及适应能力的影响,待于进一步深入研究。
References:
[1]Dong M. Cloning of Plant Ecology[M]. Beijing: Science Press, 2011.
[2]Zhang J, Zhang M R, Zhao Y K. Advances in researches of phenotypic plasticity of the clonal plant. Journal of Inner Mongolia Agricultural University, 2008, 29(4): 271-275.
[3]Zhang Y F. Study on biological characteristics ofPotentillaanserina. Acta Agriculturae Boreali-Occidengtalis Sinica, 2011, 20(12): 126-129.
[4]Eriksson O. Mobility and space capture in the stoloniferous plantPotentillaanserina. Oikos, 1986, 46: 82-87.
[5]Gou G X. A preliminary study on the growth characteristics ofPotentillaanserina. Prataculture & Animal Husbandry, 2012, 10: 14-15, 20.
[6]Zhou H K, Zhao X Q, Zhou L,etal. Alpine meadow degradation alter the clonal growing characteristics ofPotentillaanserina. Acta Ecologica Sinica, 2006, 26(2): 508-520.
[7]Cheng Y, Song J H, Liu S J. Effects of polysaccharides fromPotentillaanserinaon the levels of cytokines in tumor mice. Journal of Anhui Agricultural Sciences, 2012, 40(9): 5177-5178, 5180.
[8]Tie G C, Liu H X. The characteristics and application value ofPotentillaanserina. Journal of Qinghai Agriculture and Animal Husbandry, 2008, 1: 42-43.
[9]Zhang Q L, Chen X H, Zhang R F,etal. Study on landscape application ofPotentillaflagellaris. Shandong Forestry Science and Technology, 2006, 2: 25-26.
[10]Huang S P, Du G J, Ma F J,etal. Application ofPotentillaanserinain landscaping. Horticulture & Seed, 2012, 7: 47-48, 51.
[11]Zhou H K, Zhou X M, Zhou L,etal. The clonal growing characteristic in the stoloniferous herb,Potentillaanserina. Acta Botanica Boreali-Occidentalia Sinica, 2002, 22(1): 9-17.
[12]Shen Z X, Chen Z Z, Wang Y R,etal. Clonal growth of stoloniferous herbPotentillaanserinaon degraded and non-degraded alpine meadow soil. Chinese Journal of Applied Ecology, 2003, 14(8): 1332-1336.
[13]Li J Q, Wang L X. Elementary study on the biological characteristic ofPotentillaanserinaL. Journal of Northwest A & F University (Natural Science Edition), 2003, 31(3): 190-192.
[14]Rautiainen P, Koivula K, Hyvarinen M. The effect of within-genet and between-genet competition on sexual reproduction and vegetative spread inPotentillaanserinassp.egedii. Journal of Ecology, 2004, 92: 505-511.
[15]Saikkonen K, Koivnen S, Vuorisalo T,etal. Interactive effects of pollination and heavy metals on resource allocation inPotentillaanserinaL. Ecology, 1998, 79: 1620-1629.
[16]Eriksson O. Ramet behavior and population growth in the clonal herbPotentillaanserina. Journal of Ecology, 1988, 76: 522-536.
[17]Stuefer J F, Huber H. Differential effects of light quantity and spectral light quality on growth, morphology and development of two stoloniferousPotentillaspecies. Oecologia, 1998, 117: 1-8.
[18]Sheng H Y, Li J Q, Yang Y Z,etal. Response of phenotypic plasticity ofPotentillaanserinaL. to soil moisture. Agricultural Research in the Arid Areas, 2004, 22(3): 119-122.
[19]Stuefer J F, During H J, de Kroon H. High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments. Ecology, 1994, 82: 511-518.
[20]Wang C A, Li D Z, Zhu Z L,etal. Research methods for physiological integration of clonal plants and their application. Chinese Journal of Applied & Environmental Biology, 2006, 12(4): 581-588.
[21]Liu J P, You M H. Effect of growth inhibitors on biomass structure, energy distribution and lodging rates for populations ofElymussibiricus. Acta Prataculturae Sinica, 2012, 21(5): 195-203.
[22]Luo X G, Dong M. Architectural plasticity in response to light intensity in the stoloniferous herb,DuchesneaindicaFocke. Acta Phytoecologica Sinica, 2001, 25: 494-497.
[23]Yu F H, Dong M. Effect of light intensity and nutrient availability on clonal growth and clonal morphology of the stoloniferous herbHalerpestesruthenica. Acta Botanica Sinica, 2003, 45(4): 408-416.
[24]Xu K Y, Ye W H, Li G M,etal. Phenotypic plasticity in response to light intensity in the invasive speciesAlternantheraphiloxeroides. Journal of Wuhan Botanical Research, 2005, 23(6): 560-563.
[25]Wang Q, Liu X, Wang A L,etal. Clonal growth ofLysimachiachristinaein response to light intensity. Journal of China West Normal University (Natural Sciences), 2003, 24(4): 390-395.
[26]Saitoh T, Seiwa K, Nishiwaki A. Importance of physiological integration of dwarf bamboo to persistence in forest under storey: a field experiment. Journal of Ecology, 2002, 90: 78-85.
[27]Wang H, Li D Z, Cheng L L,etal. Effect of light intensity variation on the clonal growth characteristics ofZoysiajaponica. Ecological Science, 2014, 33(4): 664-671.
[28]Sheng L J. Phenotypic Plasticity in Response to Light and Patterns of Carbon Physiological Integration inZoysiajaponicaSteud., a Clonal Plant[D]. Shanghai: East China Normal University, 2007.
[29]Zhu X F, Zhou S B, Zheng H Q,etal. Effects of shading on biomass allocation and morphological characteristics ofDichondrarepens. Chinese Journal of Ecology, 2009, 28(7): 1419-1422.
[30]Luo D, Qian Y Q, Liu J X,etal. Phenotypic responses of a clonal plant (Buchloedactyloides) to nutrient heterogeneity. Acta Prataculturae Sinica, 2014, 23(3): 104-109.
[31]Evans J P. The effect of local resource availability and clonal integration on ramet functional morphology inHydrocotylebonariensis. Oecologia, 1992, 86: 268-275.
[32]Ye S Y, Pan S, Wang J B,etal. Research progress on physiological integration of clonal plant. Grassland and Turf, 2008, 5: 63-69.
参考文献:
[1]董鸣. 克隆植物生态学[M]. 北京: 科学出版社, 2011.
[2]张瑾, 张明如, 赵银宽. 克隆植物表型可塑性研究若干进展. 内蒙古农业大学学报, 2008, 29(4): 271-275.
[3]张彦芬. 鹅绒委陵菜的生物学特性. 西北农业学报, 2011, 20(12): 126-129.
[5]苟桂香. 鹅绒委陵菜生长特征初步研究. 草业与畜牧, 2012, 10: 14-15, 20.
[6]周华坤, 赵新全, 周立, 等. 高寒草甸退化对鹅绒委陵菜克隆生长特征的影响. 生态学报, 2006, 26(2): 508-520.
[7]成英, 宋九华, 刘素君. 鹅绒委陵菜多糖对荷瘤小鼠细胞因子的影响. 安徽农业科学, 2012, 40(9): 5177-5178, 5180.
[8]铁桂春, 刘红献. 鹅绒委陵菜的特性及应用价值. 青海农牧业, 2008, 1: 42-43.
[9]张庆良, 陈秀红, 张仁富, 等. 匍枝委陵菜的园林应用研究. 山东林业科技, 2006, 2: 25-26.
[10]黄淑萍, 杜桂娟, 马凤江, 等. 蛇莓委陵菜在园林绿化中的应用. 园艺与种苗, 2012, 7: 47-48, 51.
[11]周华坤, 周兴民, 周立, 等. 鹅绒委陵菜生长特征. 西北植物学报, 2002, 22(1): 9-17.
[12]沈振西, 陈佐忠, 王彦荣, 等. 高寒退化与未退化草甸土壤下匍匐茎鹅绒萎陵菜的克隆生长特征的比较. 应用生态学报, 2003, 14(8): 1332-1336.
[13]李军乔, 王立祥. 鹅绒委陵菜生物学特性的初步研究. 西北农林科技大学学报(自然科学版), 2003, 31(3): 190-192.
[18]盛海彦, 李军乔, 杨银柱, 等. 土壤水分对鹅绒委陵菜表型可塑性的影响. 干旱地区农业研究, 2004, 22(3): 119-122.
[20]王长爱, 李德志, 朱志玲, 等. 克隆植物生理整合作用的研究方法及其应用. 应用与环境生物学报, 2006, 12(4): 581-588.
[21]刘金平, 游明鸿. 生长抑制剂对老芒麦种群生物量结构、能量分配及倒伏率的影响. 草业学报, 2012, 21(5): 195-203.
[22]罗学刚, 董鸣. 蛇莓克隆构型对光照强度的可塑性反应. 植物生态学报, 2001, 25: 494-497.
[23]于飞海, 董鸣. 光照强度和基质养分对匍匐茎草本金戴戴克隆生长和克隆形态的影响. 植物学报, 2003, 45(4): 408-416.
[24]许凯扬, 叶万辉, 李国民, 等. 入侵种喜旱莲子草对光照强度的表型可塑性反应. 武汉植物研究, 2005, 23(6): 560-563.
[25]王琼, 刘霞, 王爱丽, 等. 过路黄克隆生长对光照强度的反应. 西华师范大学学报(自然科学版), 2003, 24(4): 390-395.
[27]王浩, 李德志, 程立丽, 等. 光资源强度变化对结缕草植株克隆生长特征的影响. 生态科学, 2014, 33(4): 664-671.
[28]盛丽娟. 克隆植物结缕草对光照的表型可塑性响应与碳素整合格局研究[D]. 上海: 华东师范大学, 2007.
[29]朱肖锋, 周守标, 郑和权, 等. 遮光对马蹄金生物量分配和形态特征的影响. 生态学杂志, 2009, 28(7): 1419-1422.
[30]罗栋, 钱永强, 刘俊祥, 等. 克隆植物野牛草对异质营养的表型可塑性响应. 草业学报, 2014, 23(3): 104-109.
[32]叶上游, 潘爽, 王景波, 等. 克隆植物生理整合作用研究进展. 草原与草坪, 2008, 5: 63-69.
Effect of partial shading on the morphological plasticity and biomass allocation ofPotentillaanserina
FAN Xing, CAI Jian, LIU Jin-Ping*, LI Ying, ZHANG Xiao-Jing, ZENG Xiao-Lin
KeyLaboratoryofSouthwestChinaWildlifeResourcesConservation,MinistryofEducation,ChinaWestNormalUniversity,Nanchong637009,China
Abstract:In this study, genet, mother ramet and some clonal fragments of Potentilla anserina were used to analyze the effect of partial shading. The quantitative traits and biomass allocation of root, stem and leaf were also measured. The purpose of the study was to analyze the effect of partial shading on the morphological plasticity and biomass allocation of the mother ramet, daughter ramet and genet of P. anserina. The results showed that: 1) For mother ramets, partial shading displayed a significant effect on plant height, leaf length, leaf width, leaf area and root depth (P<0.05). Partial shading increased plant height, leaf length and total leaf area, but different shading positions or shading intensities had different effects on leaf width, single leaf area and root length. 2) For daughter ramets, partial shading had a significant impact on leaf length and leaf area. Shading of the clonal fragment was the main factor affecting leaf size of daughter ramets, with shading of mother ramets second placed. 3) Partial shading significantly increased the number of cloned fragment daughter ramets and new daughter ramets, and also increased the cloned fragment number of genets (P<0.05). Shading mother ramets had an important influence on stolon elongation and the formation of new cloned fragments. 4) Partial shading displayed a highly significant effect on the root, stem and leaf water content of genets, as well as on their leaf and root biomass accumulation (P<0.01). With shading of more than 2 clonal fragments or mother ramets, the root biomass accumulation of genets significantly increased, but with full shading of genets the root biomass was clearly lower than the control. 5) Partial shading also had significant effects on the allocation ratio of stem, leaf and root biomass of genets (P<0.01). Shading had the greatest effect on leaf allocation ratio and a lower effect on stem allocation ratio, but the lowest effect was on the root allocation ratio. 6) Partial shading had a significant effect on the underground biomass ratio (P<0.01). Shading had an important effect on the root/stem and stem/leaf ratios (P<0.01), which increased remarkably with full shading. In summary, partial shading displayed a direct influence on the morphological plasticity and biomass allocation of various types of P. anserina plants, including mother ramets, daughter ramets, clonal fragments and genets.
Key words:Potentilla anserina; clonal plant; shading; morphological plasticity; biomass allocation
*通信作者
Corresponding author. E-mail:jpgg2000@163.com
作者简介:樊星(1992-),男,四川攀枝花人,在读硕士。E-mail: aqz348985051@qq.com
基金项目:四川省科技支撑计划(2011NZ0064)和校基金(11A035)项目资助。
收稿日期:2015-04-23;改回日期:2015-05-20
DOI:10.11686/cyxb2015208
http://cyxb.lzu.edu.cn
樊星,蔡捡,刘金平,李莹,张小晶,曾晓琳. 局部遮光对鹅绒委陵菜基株形态塑性及生物量配置的影响. 草业学报, 2016, 25(3): 172-180.
FAN Xing, CAI Jian, LIU Jin-Ping, LI Ying, ZHANG Xiao-Jing, ZENG Xiao-Lin. Effect of partial shading on the morphological plasticity and biomass allocation ofPotentillaanserina. Acta Prataculturae Sinica, 2016, 25(3): 172-180.
