粮食作物对高温干旱胁迫的响应及其阈值研究进展与展望
2016-04-19王润元王鹤龄赵福年
赵 鸿,王润元,尚 艳,王鹤龄,张 凯,赵福年,齐 月,陈 斐
(1. 中国气象局兰州干旱气象研究所,甘肃省干旱气候变化与减灾重点实验室,中国气象局干旱气候变化与减灾重点实验室,甘肃 兰州 730020; 2. 甘肃农业大学生命科学技术学院,甘肃 兰州 730070)
粮食作物对高温干旱胁迫的响应及其阈值研究进展与展望
赵鸿1,王润元1,尚艳2,王鹤龄1,张凯1,赵福年1,齐月1,陈斐1
(1. 中国气象局兰州干旱气象研究所,甘肃省干旱气候变化与减灾重点实验室,中国气象局干旱气候变化与减灾重点实验室,甘肃兰州730020; 2. 甘肃农业大学生命科学技术学院,甘肃兰州730070)
摘要:以大气温度升高和降水波动为主要标志的气候变暖对农业生产产生了重要影响,农作物生长发育、形态建成、生理生化过程等对气温、水分变化的响应特征、机理与后果等的研究,对揭示气候变化对农作物的影响及其机制具有重要作用,是制定适应对策的重要前提之一。本文分别回顾了国内外水稻、小麦、玉米等主要粮食作物生长、发育、生理生态因子、产量、水分利用效率等对高温、水分亏缺的反应以及对二者的协同响应,评述了高温和干旱缺水影响过程中作物的阈值反应及其临界值,讨论了当前高温干旱对作物影响研究中存在的问题。在此基础上,提出了今后应着重加强研究的关键科学问题:(1)干旱/湿润条件下的温度、水分阈值,以及多因子协同胁迫下作物的忍耐极限;(2)胁迫程度、时期、历时与作物自身生理生化过程的关系,以及细胞和分子水平上的响应机制;(3)作物对适度干旱的补偿效应在高温下是被削减还是增加,需要进一步研究和探索。
关键词:粮食作物;高温;水分亏缺;生长发育;生理生态;产量;阈值
引言
最新的观测事实证明,全球变暖毋庸置疑。近130 a (1880~2012年)来,全球平均地表温度升髙0.85 ℃;20世纪中叶以来,全球平均地表温度的升温速率 (0.12 ℃ /10 a) 几乎是1880年以来的2倍;过去30 a (1983~2012年)是1850年以来最暖的3个10 a,气候变暖趋势在全球各地几乎都可以观测到[1]。气温、降水的变化对农业生产的不利影响更加显著[2-6],其中小麦、玉米相对水稻和大豆受到的影响更大[2,7],气候变化导致小麦和玉米单产平均每10 a减少约1.9%和1.2%[7]。因此,研究农作物与气温、水分变化的关系,特别是明确动态变化过程中一些临界值(阈值)的形成及其机制,对于深入开展气候变化对农作物的影响研究、探索气候变化对农业生产的阈值、及时发现可能存在的问题、尽早采取适应对策具有非常重要的战略意义。
农业生态系统中的阈值(threshold),又叫临界值,是指某一(些)生态参数达到某一临界点的值,当低于或高于这个值时将引起某些性状或过程的突变[8-9]。根据生态学限制因子法则,不同的生物过程、生态因子等都存在阈值,如作物生长过程中植株叶片/组织扩展、生理过程的土壤有效水分阈值,以及温度、水分、水势等环境因子阈值等[8-9]。本文就国内外近年来主要粮食作物生长发育、生理特征、形态塑性、产量形成等对气温增加、水分亏缺等的响应及其阈值问题进行了回顾,以期为进一步深入开展相关研究奠定基础。
1主要粮食作物对温度升高的响应
温度是作物生长发育的关键气象要素,大量研究表明温度变化将对作物生产造成深远影响,尤其对水稻和小麦两大粮食作物[10-12]。对于春小麦和大麦,生长发育的最适宜温度是20~25 ℃[11]。温度升高农作物生长季提前[13-14],成熟期也提早出现[15-16],这使得作物生育期发生改变,其中以生育期缩短为主要表现形式[17-19]。温度的变化通过影响叶片的气孔导度和土壤蒸发速率来影响作物蒸散发过程[20-22],一般认为,在一定的临界值以下,随着温度升高,叶片气孔导度增加,且净光合速率增加的幅度大于蒸腾速率增加的幅度,导致叶片水分利用效率提高;当超过该临界值时,温度升高,叶片蒸腾增加,又导致叶片水分利用效率降低[23]。但也有不同的认识,温度升高,春小麦叶片气孔导度减小,从而导致叶片水分利用效率降低[20,24]。轻度高温胁迫对作物光合抑制主要由气孔限制引起,而严重胁迫时则不然[25]。由于高温下较高的新陈代谢、生长发育和蒸散发[21],造成作物对水分的需求增加[26]。当生长资源被热胁迫限制时,植物器官如叶片、分蘖、小穗的大小都相应减小[25]。高温通过加速物候期和减少生物量影响籽粒产量[27-28],高温增加了籽粒灌浆速率,但减少了籽粒灌浆持续期,使得千粒重等产量构成要素降低[29-30]。 作物新陈代谢过程对温度的增加非常敏感,高温胁迫影响植物体内核糖核酸RNA与蛋白质的结构,破坏细胞膜完整性,干扰细胞骨架的动态重塑,改变细胞内酶促反应速率,从而导致植物体内代谢紊乱[31-32]。气候变暖在加速农作物生长的同时,也使农作物的呼吸作用增强,加上高温下生育期缩短,导致热环境下的生物量和产量降低[14,33],如水稻结实期温度增加1~2 ℃时,产量将下降10%~20%,这种负影响随着纬度的升高而加重[34]。温度每增加1 ℃,玉米产量将减少3%。小麦也会由于气温升高、降水减少而不同程度减产[34]。这些变化继而引起物种地理分布的改变[35]。不同作物、不同品种的响应存在显著差异[13,33],不同海拔高度也差异显著[16],相同增温条件下水稻减产幅度高于小麦[36]。当冠层温度增加时,冬小麦籽粒蛋白质含量下降6.9%~9.1%,而籽粒中直/支淀粉的比例均显著提高[37]。增温导致高温热害的潜在风险提高、农田水分供应降低[38]。除了温度变化对农作物的影响外,昼夜温差的变化也会影响作物生长,如冬小麦生育期内昼夜温差能提高适应较小温差的小麦品种的净光合速率,但是适应较大温差的冬小麦品种的光合速率并没有增加,其适应性主要表现为叶面积的增加、叶片寿命延长以及叶片夜间呼吸强度的降低等[39]。从分子水平上来看,苗期和成熟期小麦叶片转录组对高温胁迫的响应模式基本一致,表现为苗期和成熟期的小麦叶片通过激活相同的基因和代谢途径来抵御高温胁迫[40]。
2主要粮食作物对水分亏缺的响应
干旱,作为降水减少、水分亏缺或者温度升高的结果之一,是全世界作物生产中最主要的限制因子之一,在干旱、半干旱地区表现尤为明显[41-43]。作物常常遭遇不同持续时间和强度的干旱[44-45],全世界每年由于水分亏缺导致的作物减产超过其他因素造成减产的总和[44]。作物对干旱胁迫的响应表现在分子水平、细胞水平和生理水平上[46-47],该响应除了取决于受旱的轻重程度和缺水时间的长短[48-49]、作物生长发育时期和阶段[50-51]外,也取决于不同作物的物种和基因型[52-53]、器官和细胞类型[54-55]、亚细胞区室[56]等。
作物通过土壤中的水分供应进行光合作用和干物质积累,其积累量的大小表现在株高、茎粗、叶面积和产量形成的动态变化上[41,53]。当土壤干旱或供水不足时,根系吸收的水分有限,向地上部分输送的水分减少,对地上部分的影响比对地下部分的影响增大,根冠比增加[55]。适度而缓慢的水分亏缺可增加绝对根重,抑制地上部分的生长,减少地上部分的干物质积累[50-51]。随着胁迫程度的加强,植株茎杆节间变短、叶面积减少、叶片数量增加缓慢,分生组织细胞分裂减慢或停止、生长速率大幅降低,细胞伸长受到抑制、出现植株个体低矮、光合叶面积明显减小[50-51],同时叶片组织内膨压下降,叶肉细胞扩张受阻,致使叶片出现萎蔫甚至干枯死亡[57-59]。土壤含水量降低引起叶片相对含水量和叶片水势降低、保卫细胞压力势降低而失水,引起气孔减小或关闭[60],阻止CO2进入叶片,光合速率下降,但这一过程持续的时间较短[55-56]。缺水时作物叶片生长减缓,光合面积显著减小,由于淀粉的水解作用增强,糖类积累增加,既影响光合产物的输出,又促进呼吸作用,使净光合速率降低,呼吸消耗增加[61-62]。禾谷类作物在拔节和抽穗期间遇到水分胁迫时,植株节间细胞的扩张生长受到抑制,植株矮小,还有可能抽不出穗子[48,51]。如果水分胁迫发生在灌浆期,会引起灌浆时间缩短、灌浆速率下降、植株老化提前,最终导致籽粒产量下降[61-64]。
当土壤干旱时作物叶片、花、籽粒发育过程中植物内源激素ABA浓度明显增加,且浓度随着干旱程度的变化而变化[62,65]。干旱胁迫时,植物体内清除活性氧的机制遭到破坏,酶保护系统超氧化物歧化酶(SOD)活性和含量下降,使避免或减轻植物受到氧化伤害的能力下降[62,65]。轻度胁迫可引起叶片过氧化氢酶(CAT)活性上升,而重度胁迫下CAT活性则下降。随着干旱胁迫时间的延长,叶片的过氧化物酶(POD)活性逐渐上升,丙二醛(MDA)含量不断增加,膜脂过氧化加剧,明显地抑制了植株的生长发育[62,65]。作物植株必须避免或忍耐细胞脱水来渡过干旱[66-67],避旱涉及植株快速生理生长、叶片卷曲、叶片遮阴、叶面积减小、增加气孔和表皮阻力等[66-68]。在水分亏缺加重的情况下,耐旱通过保持充足的细胞膨胀来维持新陈代谢[67]。在某些情况下,渗透调节能够使水分吸收持续。许多物种胁迫增加时,渗透调节与干旱胁迫下生长和稳定产量密切相关[68-69]。
干旱缺水并不总是造成作物产量降低,一定时期适当的水分亏缺可能对提高作物产量和水分利用效率反而有利[70],即作物对干旱缺水有一个从“适应”到“伤害”的过程,不超过适宜范围的缺水,一般在复水后,产生生理上、生长上以及水分利用上的补偿或超补偿效应,这对作物增产更加有利[70]。
3主要粮食作物对高温干旱的协同响应
高温和干旱是限制农作物生产的重要因子,自然界中这2种非生物胁迫常常同时发生[71-73],对大田作物造成很大的甚至致命的影响[74-75],尤其在雨养农业区[75-76]。虽然干旱是直接影响植物水分状况的环境胁迫因子,但是其危害的严重度很大程度上取决于温度[77]。高温能够增加水分胁迫的程度,作物对一种胁迫的响应可以被其他的胁迫及其程度来改变[40,78]。大气温度和土壤温度的升高会提高作物蒸腾耗水量和农田蒸散量,增加作物总耗水量,造成干旱缺水或进一步加剧干旱胁迫的危害[79-80]。多年多点资料分析表明,气温升高会加剧作物蒸发过程,当增温1.5 ℃时,华北冬麦区小麦全生育期内农田最大蒸散将增加50~70 mm,比当前气候下高8%~12%,实际蒸散可能增加1%~2%,其结果将导致小麦水分亏缺程度加剧[79]。模型模拟结果表明, 温度每升高1 ℃,农田潜在蒸散平均增加2.6%,最高可达3.7%[80]。当土壤水分充足时作物几乎保持稳定的水分关系,如土壤含水量、叶片相对含水量、叶水势、叶渗透势、叶细胞膨胀势和渗透调节等,温度升高对其影响不大,但当土壤水分亏缺时升温对水分关系会造成很大影响[67]。
Out-of-code structural design of Jing Feng Center building in Nanjing
植物对高温和干旱2种不同胁迫的分子和代谢响应是特有的[81],并且高温和干旱胁迫同时发生时其胁迫效应并不是2个单独胁迫的简单相加,而是会相互作用而激活不同于单独胁迫的响应基因和代谢途径[40,82]。二者结合起来到底如何影响植物?目前也有一些研究和探索[40,46]。有研究报道了高温和干旱胁迫对玉米、大麦、小麦等作物的生长和生产力的协同影响,如Vikender等[83]认为小麦开花后的高温干旱缩短了其成熟期、籽粒灌浆期、降低了籽粒产量、平均籽粒重量、籽粒数和千粒重;Hossain等[75]通过鉴定俄罗斯东南部各种基因型不同的大麦和小麦的适宜播种时间,研究了4个大麦和2个小麦耐热耐旱性基因型,结果表明在晚播时遇到高温和干旱协同胁迫,会缩短播种到现芒、抽穗和成熟收获的时间,最终对生长发育产生负面影响,导致植株群体密度降低,每株植株的分蘖数、高度和干物质减小。低温早播增加了发芽出苗的天数,减少了幼苗成苗率和分蘖能力,最终也影响了作物的生长发育;Pradhan等[73]认为春小麦从出苗到开花期遭遇干旱、高温及其协同胁迫可降低籽粒产量69%、81%、92%,而从出苗到花后21 d遭遇干旱、高温及其协同胁迫可降低籽粒产量26%、37%、 50%。增温1、2和3 ℃,降水不变条件下,春小麦产量分别下降8.4%、15.1%、21.8%;降水减少20%条件下,春小麦产量分别下降12.1%、24.7%、42.7%;降水增加20%条件下,春小麦产量分别下降9.0%、15.5%、22.2%。春小麦籽粒淀粉含量随温度的增加而下降,籽粒蛋白质含量随温度的增加而上升。可见2种胁迫的累加负效应大于单独胁迫的负效应。刘振山[40]对小麦抗旱耐热品种TAM107的幼苗进行了高温、干旱和二者协同胁迫,并用高通量RNA-Seq对比分析了3种胁迫条件下的转录表达谱。结果表明当高温和干旱胁迫同时发生时其胁迫效应并非简单相加,而是会相互作用激活不同于单独胁迫的响应基因和代谢途径。另外,他们基于高通量RNA-Seq和IWGSC释放的基因组信息,开发了小麦部分同源基因表达定量新算法,并对高温和干旱胁迫下小麦部分同源基因的表达分化进行了分析。结果发现,正常条件下约37%的基因发生了部分同源基因表达分化,干旱和高温胁迫后部分同源基因表达分化的比例更高(约39%~55%),并且胁迫程度越高部分同源基因表达分化的比例越高,对胁迫响应程度越高的基因其部分同源基因表达分化比例也越高。还发现部分基因对胁迫响应的互补效应,即在不同的胁迫类型以及胁迫后不同的时间点表现出响应。由此他们推测部分同源基因的表达分化可能有利于小麦对干旱和高温等非生物逆境的适应。
在热应激条件下,小麦、玉米、水稻等能诱导合成热激蛋白(heat shock proteins,HSPs)来适应高温逆境,它对减轻逆境胁迫引起的伤害有很大的作用。不同发育阶段的热激蛋白存在差异,不同抗热作物品种中热激蛋白也存在差异[84]。玉米的最适诱导温度为40~42 ℃,水稻在37~40 ℃开始合成热激蛋白,在43~46 ℃之间合成量最大。Ristic等[85]发现抗干旱和高温的一个玉米品系可以大量合成一种45 kD的热激蛋白,而对干旱和高温敏感的品系则不能。
在水分胁迫时,某些植物体内可主动积累各种有机或无机物质来提高细胞液浓度,降低渗透势,提高细胞吸水或保水能力,从而适应水分胁迫的环境,这一过程为渗透调节[86-87]。干旱环境下,渗透调节在许多C4植物的抗旱性中起到非常重要的作用,并且能使高粱在叶水势非常低的时候仍然能够保持生长[71]。C3作物小麦遇到水分胁迫时,也会发生渗透调节现象,但是往往不如在高粱中明显,并且不同基因型间存在一定的差异[87]。高粱在高温下也表现出明显的渗透调节,该高温对小麦常常是致命的[67]。增加小麦耐热性可能会提高其适应高温干旱的潜力[88]。
在作物生长最敏感时段如生殖生长和灌浆期避开高温干旱胁迫,作物遭受的负面影响就可以降低[75,89],一般可通过调整播种期或者选用早熟品种来应对[90-91]。在约旦,适合地中海气候的品种有较长的预起身期,其后发育期缩短,灌浆速率较高来避免后期的干旱和高达25~31 ℃的高温胁迫[91]。就耐性来说,C4谷类作物高粱一般比C3作物小麦更加耐热耐旱[78,92]。不同品种对胁迫的抗性表达存在很大差异,甚至有时是相反的,如Aprile等[81]研究认为对于有抗旱性的小麦品种Ofanto来说,高温干旱协同胁迫导致表达量发生显著改变的基因数量增加,超过单独胁迫时的累加影响。但是,对于品种Cappelli,相同的胁迫处理却仅仅引起比单独热胁迫处理还要少的基因表达量发生显著改变。干旱和高温胁迫同时发生时会相互作用加强对植物的胁迫效应,除了单独胁迫响应基因之外,植物也会激活一些特殊的基因和代谢途径来抵御这种更强的胁迫,从而形成多重胁迫下植物更复杂的基因表达调控机制[40]。
4主要粮食作物对温度的阈值反应
温度是农作物生命活动的生存因子,它直接影响作物的生长发育和最终产量,尤其是生长发育速率。作物生长、繁殖以及整个生活史都需要在一定的温度范围内才能进行,通常维持作物生命的温度范围大致在-10~50 ℃之间,而适宜农作物生长的温度在5~40 ℃之间,农作物发育要求的温度通常为20~30 ℃。作物的各种生理活动都要求有三基点温度,即最低温度(Tmin)、最适温度(Topt)、最高温度(Tmax)。在最适温度下,作物生长发育迅速而且良好。在最高和最低温度下,作物停止生长发育,但仍然能够维持生命。三基点温度是最基本的温度指标,它在确定温度的有效性、作物种植季节与分布区域及计算作物生长发育速度、光合潜力与产量潜力等方面,都有非常重要的作用[93]。三基点温度受作物种类、生育时期、生理状况、不同器官等因素的影响而存在一定的差异(表1和表2)。最低、最适、最高温度指标都不是一个具体的数值,而是具有一定的范围,不仅与强度有关,还与作用的持续时间有关。无论是生存、生长还是发育,最适温度基本上是在同一个变幅范围,差异很小。各种作物最低温度的最低点差异很大,耐寒作物可以忍受-10~-20 ℃以下的低温,而喜温作物甚至不能安全度过0 ℃左右的温度,且最低温度与最适温度差值较大。与最低温度比较,各种作物最高温度指标值差异较小,且各种作物的最高温度与最适温度值也比较接近。在作物的生命过程中,最低温度远较最高温度出现的几率大[93]。
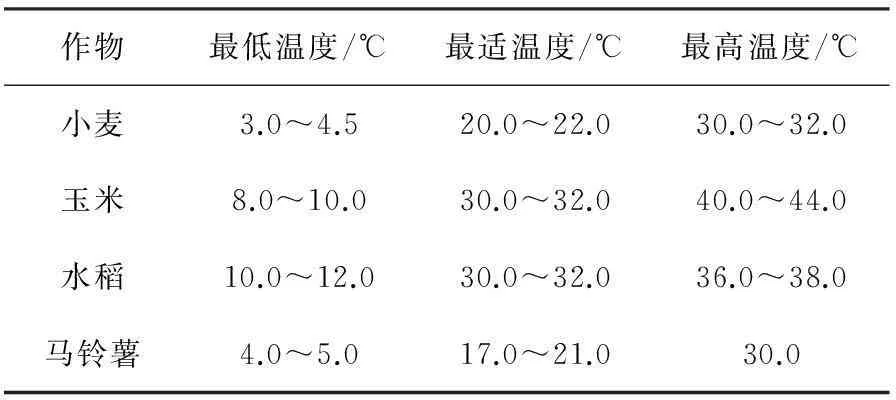
表1 几种主要作物的三基点温度[93]
在最低、最高温度的范围内,如果温度继续升高或降低,就会对作物产生不同程度的危害,直至死亡,所以在三基点温度之外,还可以确定使植物受害或致死的最高与最低温度指标,即致死最高温度(LTmax)、致死最低温度(LTmin)[94],与三基点温度合并起来称为5个基本点温度[93],各类植物能忍受的最高温度界限是不一样的,图1描述了春小麦的5个基本点温度[94-95]。

表2 主要作物不同发育阶段对温度和水分的要求及其临界值[96-97]
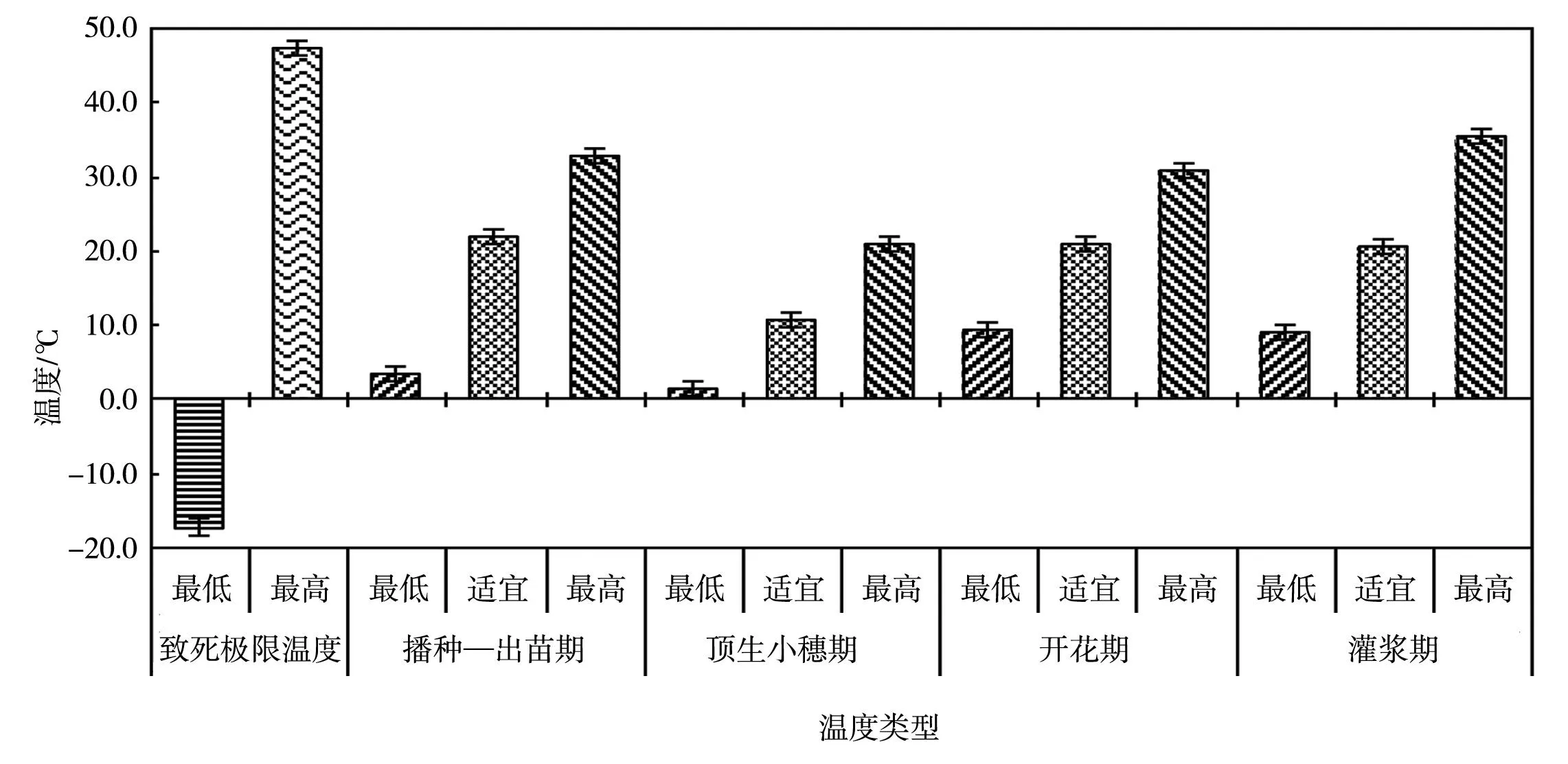
图1 小麦不同生育期的最低、适宜、最高温度以及全生育期内的致死极限温度[94-95]
温度对作物的生长发育固然重要,但温度变化尤其超过一定的临界值,会对作物生长带来很大的甚至致命的影响。如Wheeler等[98]研究发现作物在开花前后将温度突然增加2 ℃比平均增加2 ℃对作物产量的影响更大;Dupuis等[99]认为温度>35 ℃对玉米花粉活力是致命的;Prasad 等[100]通过高温对高粱的影响研究,发现高粱籽粒产量、收获指数、花粉活力的最适温度是22~32 ℃,生物量、光合速率和籽粒大小的最适温度分别是30~40 ℃、34~44 ℃和26~36 ℃。有些农作物的生长发育对温度变化较敏感,尤其是玉米、水稻等喜温作物。周曙东等[101]根据谢云[102]提出的喜温作物温度影响函数绘制了喜温作物对温度的敏感性曲线图,发现喜温作物的5个敏感临界点温度,分别是6、21、28、32和44 ℃。当温度<6 ℃时,喜温作物停止生长,继续低至0 ℃以下时,则可能会产生冻害;温度在6~21 ℃时,作物开始生长发育,但生长速度缓慢;温度在21~28 ℃之间时作物生长速度明显加快;在28~32 ℃之间温度最适合喜温作物的生长;当>32 ℃时,随着温度上升,作物生长明显受到影响;当温度>44 ℃时,喜温作物停止生长。0 ℃与44 ℃温度敏感临界点表明喜温作物所能承受的温度最低阈值与最高阈值,当温度一旦超过这个阈值时,作物就会停止生长甚至死亡。
5主要粮食作物对水分亏缺的阈值反应
根据生态学的限制因子法则,植物生理过程对土壤水分的需求存在不同水平的临界值(阈值),水分过多或过少都会影响植物的光合生理过程[104]。土壤水分阈值受土壤、植物和气象条件等因素的影响。不同土壤类型之间差异显著,在黄土区土壤水分阈值大小顺序是塿土 (田间持水量的88%) 、黄绵土(田间持水量的78%)、黑垆土 (田间持水量的70%)[105]。就一些植物叶片扩展的平均阈值来说,在不同的土壤质地和土壤类型条件下差异显著,粗粒土和细粒土的土壤水分阈值分别为0.72和0.43[8]。不同作物的阈值有明显差异,双子叶植物(0.66)高于单子叶植物0.50[8,106]。当土壤湿度在60%这一临界值以上时,土壤含水量对冬小麦的气孔阻力、叶水势、光合是等效的,只有低于此临界值时,随着土壤湿度的降低,气孔导度、叶片水势和光合才明显降低,作物本身的水分运输受到影响,表现出明显的阈值反应,而夏玉米的土壤湿度阈值高于60%[107]。玉米连续干旱25 d,土壤干旱程度达到0.55后,玉米产量降低5%,将此作为玉米拔节—抽穗期土壤的干旱阈值,该值既可以表示土壤干旱状况又能表示作物受旱程度[108]。植物的品种或基因型不同,土壤水分阈值也存在较大差异[109]。
除了土壤、植物等因素外,土壤水分阈值还会受到不同气象因子的影响。有研究认为玉米日蒸腾速率随土壤含水量的动态变化过程受到大气蒸发力的影响,土壤水分阈值随着大气蒸发力的升高而升高[110],而水汽压亏缺对土壤水分阈值不产生影响[111]。但也有研究认为,当参考蒸散量< 3 mm·d-1时,土壤水分阈值随大气蒸发力的增大没有明显升高,相反,当参考蒸散量> 3 mm·d-1时,土壤水分阈值随着大气蒸发力的增大而明显升高[112]。由于印度地处热带地区,大气蒸发力强,雨养农业区连续2周内20 mm的降水量即为水稻的干旱阈值[113]。
植物可利用的土壤水分(plant available soil water,PAW)是广泛用于衡量植物对水分亏缺响应的敏感参数,Sadras等[8]回顾对比了一些植物种类(包括单子叶、双子叶植物)及其生长过程(包括叶片扩张、气体交换等变量)中的PAW,指出了PAW值变化的整个可能范围(0~1),评述了一些变量的平均阈值,如组织扩张、气体交换、叶水势、气孔导度等过程中的平均阈值分别为0.56、0.4、0.61、0.37,非常接近于组织扩张和气体交换的平均阈值,这些值对水分亏缺有较大的依赖性 (图2)。在快速生长的组织中细胞扩张、细胞壁合成、蛋白质合成过程是对水分亏缺最敏感的生理过程。在植物水平上,叶片扩张是对水分亏缺最敏感的过程之一[8]。相对于组织扩张来说,气孔导度对水分亏缺的响应较少。就气孔导度的受限、通过水分胁迫的非气孔影响将会导致光合速率减少[8]。由于不同的生理参数对土壤水分亏缺的敏感性不同,已有报道的研究结果差异较大,如玉米、小麦等作物叶片或组织扩展的土壤水分阈值高于气体交换指标(包括蒸腾和光合速率的土壤水分阈值)[8]。
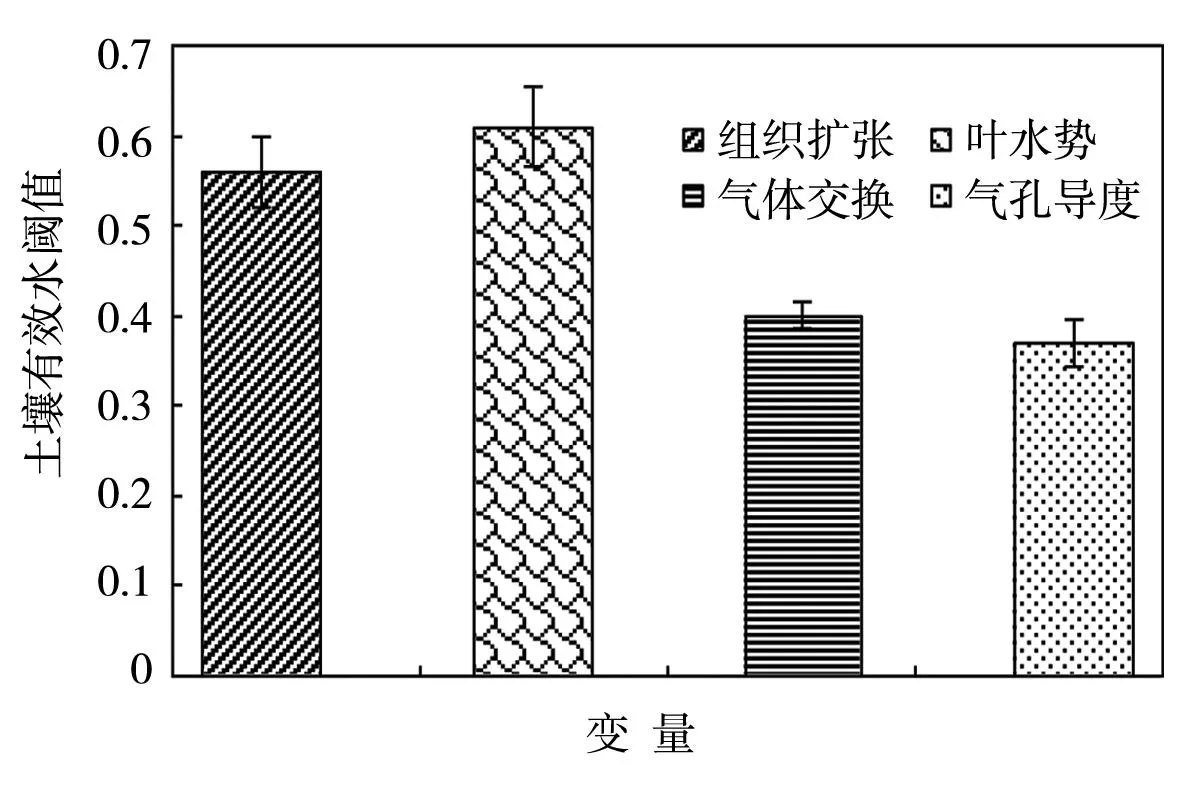
图2 植株组织扩张、叶水势、气体交换和
6存在的问题
通过以上的归纳分析,可以看出气温升高与降水减少加剧了水分的不足,对农业生产带来了一些不可逆转的负面影响,造成主要粮食作物生长发育受阻、产量下降、水分利用效率低下等。虽然国内外已有大量研究揭示了这些影响,但因作物生长环境复杂,涉及的因素较多,因而也存在一些问题。
(1)气象因素中温度增加、水分减少、CO2浓度升高、光照等的任何变化都会影响到作物的生长发育和产量形成。不同环境条件(温度、光照、水分)下植物的生长发育和生理活动,随着作物品种类型、基因型、所处的气候环境、地理位置等的不同而有所差异,使得目前的大部分研究(局限于单一因子的影响)显得极为单薄,也偏离了作物生长的真实环境。近年来陆续有一些两因子耦合对作物影响的试验研究,但三因子耦合甚至多因子耦合的协同影响研究也还很少,多因子协同作用下作物反应和适应性如何,相互间如何反馈和调控?这些都是亟待解决的科学问题。
(2)目前大多数研究侧重于作物生理水平上的响应,并且研究结果有时存在分歧,而对于细胞及分子水平的响应研究,虽然证明了高温干旱协同胁迫效应并非其单独胁迫效应的简单相加,但对于高温和干旱胁迫同时发生时的互作效应以及植物对其协同胁迫的抵御机制尚不清楚,这些不能满足对作物逆境胁迫响应的认识。因此,需要继续深入研究气温升高、降水变化对作物细胞水平和分子水平的影响,并阐明作物对环境因素变化响应的分子机理,从而对作物响应暖干化气候得出更加合理、科学的解释和适应对策。
(3)高温和干旱缺水对作物的影响研究中,测试的各种响应指标和阈值都是在特定的土壤类型、土壤肥力水平、气候、作物种类、品种等因素下进行的,因而在一定程度上削弱了其影响的代表性。所以,迫切需要研究同一作物或不同作物在多地区对气温升高、降水波动、CO2浓度升高等逆境胁迫的响应,以期找出对逆境响应的异同以及对逆境响应出现最大值或最小值反应的指标及其临界点,以利于作物对高温干旱应对技术的制定和防灾减灾工作的进行。
7研究展望
IPCC第五次评估报告指出,预计2016~2035年全球平均地表温度将继续升高0.3~0.7 ℃,全球降水将呈现“干者愈干、湿者愈湿”的趋势[1,7]。气候变暖很容易产生不可逆转的影响[2]。与 20 世纪末相比,如果气温升高2 ℃,将会给热带、温带地区的小麦、玉米、水稻生产产生负面影响;如果升温超过4 ℃,有可能对全球粮食安全产生重大影响[2,5]。因此,为了最大限度地减小气温升高、降水减少对作物的不利影响,利用气候变暖的有利因素,需要更进一步了解气候变化对主要粮食作物的影响,为不同区域因地制宜地制定适应对策提供理论依据,希望继续着重进行以下几个方面的研究。
(1)充分利用各种研究手段,如大田控制试验、遮雨棚、水分池、温室、人工气候室等模拟农作物的生长环境,结合统计分析、作物模型等,综合分析温度、水分等因子的作用和地位,确定各自的影响阈值,是揭示农作物对高温、干旱缺水等胁迫响应的基础。
(2)高温干旱的协同影响比单独胁迫影响更有害,自然界中的协同影响往往是累加的,尤其在降雨波动较大的地区。干旱/湿润条件下的温度、水分阈值及其相互关系如何?协同影响机制怎样?在此基础上,进一步研究高温、水分亏缺以及CO2浓度增加等多因子协同作用下作物的忍耐极限以及相互关系等,是深入认识作物对气候变化适应的关键所在,也会促进植物水分生理、抗性生理、农业干旱、气象干旱等边缘学科的交叉渗透。
(3)植物对水分亏缺的响应特征中,植物可利用的土壤水分PAW已经引起了科学家足够的重视。然而,在已有的对生理关系的共识中,对于给定的过程和种类,PAW的测量值受到蒸发需求、根的分布、土壤结构和土壤容重等因子的影响,因此,在模式模拟中使用固定的土壤水分阈值时要做一些相应的假设,是客观评估作物受胁迫的重要途径之一。
(4)作物在适度干旱后复水存在一定的补偿或超补偿效应,水分亏缺对冬小麦等作物生长造成的滞后效应在复水后将成为作物快速生长的驱动因素。已有的研究较少考虑作物的旱后补偿效应或者仅仅是在土壤水分亏缺下的补偿效应。那么在气温升高的情况下,作物的这种补偿效应是被削减还是增加?值得继续研究探讨。
(5)在研究开展作物高温、干旱胁迫致灾机理的相关研究时,需重视胁迫程度、时期、历时与作物自身生理生化过程的关系,系统认识作物受害过程中不同生育期各种生理生态参数、形态建成等指标持续变化特征以及发生突变的动态轨迹,并对其进行定量描述,了解致灾的生理机制和关键影响因子,是科学制定适应对策的前提。
(6)高温和干旱缺水是我国农业生产健康发展的主要限制因素,尤其在西北干旱半干旱区,降水少,水资源短缺问题尤为突出。在该地区研究缺水条件下遇到高温胁迫时作物生长发育和产量形成的规律和机理,了解高温干旱对主要粮食作物伤害的生理生化机制以及细胞和分子水平上的响应机制,为防御作物高温热害和干旱伤害提供科学依据,同时也为进一步研究高温干旱环境下作物的抗性机制和抗逆作物优质育种打下基础,对于科学利用当地有限的降水资源、有效抗御干旱、提高农作物产量等具有重要意义。因此,缺水条件下作物受到的高温胁迫研究具有广阔的应用前景。
参考文献:
[1] IPCC. Climate Change 2013: the physical science basis. Contribution of Working Group I to the fifth assessment report of the Intergovernmental Panel on Climate Change[R]. Cambridge & New York: Cambridge University Press, 2013.
[2] IPCC. Climate Change 2014: impacts, adaptation and vulnerability. Contribution of Working Group II to the fifth assessment report of the Intergovernmental Panel on Climate Change[R]. Cambridge & New York:Cambridge University Press, 2014.
[3] 吴普特,赵西宁. 气候变化对中国农业用水和粮食生产的影响[J]. 农业工程学报, 2010,26(2):1-6.
[4] 张强,陈丽华,王润元,等. 气候变化与西北地区粮食和食品安全[J]. 干旱气象, 2012,30(4):509-513.
[5] 周广胜. 气候变化对中国农业生产影响研究展望[J]. 气象与环境科学,2015,38(1):80-94.
[6] 郑冬晓,杨晓光. ENSO对全球及中国农业气象灾害和粮食产量影响研究进展[J]. 气象与环境科学,2014,37(4):90-101.
[7] 秦大河. 气候变化科学与人类可持续发展[J]. 地理科学进展,2014,33(7):874-883.
[8] Sadras V O, Milroy S P. Soil-water thresholds for the responses of leaf expansion and gas exchange: A review[J]. Field Crops Research, 1996,47:253-266.
[9] 许振柱. 植物的水分阈值与全球变化[J]. 水土保持学报,2003,17(3):155-158.
[10] FAO. Rice in Human Nutrition[M]. Food and Agriculture Organization of the United Nations, Rome, 1993.
[11] Hakim M A, Hossain A, Teixeira da Silva J A, et al. Yield, protein and starch content of 20 wheat (Triticum aestivum L.) genotypes exposed to high temperature under late sowing conditions[J]. J Sci Res, 2012,4(2):477-489.
[12] Hossain A, Lozovskaya M V, Zvolinsky V P, et al. Effect of soil and climatic conditions on phenology of spring wheat varieties in the northern Bangladesh[J]. Nat Sci:J Fund Appl Sci, 2012,2(39): 78-86.
[13] Wang R Y, Zhang Q, Wang Y L, et al. Response of corn to climate warming in arid areas in northwest China[J]. Acta Botanica Sinica, 2004,46(12):1387-1392.
[14] 赵鸿,王润元,王鹤龄,等. 西北干旱半干旱区春小麦生长对气候变暖响应的区域差异[J]. 地球科学进展,2007,22(6):636-641.
[15] Porter J R. Rising temperatures are likely to reduce crop yields[J]. Nature, 2005,436:174.
[16] 赵鸿,肖国举,王润元,等. 气候变化对半干旱雨养农业区春小麦生长的影响[J]. 地球科学进展,2007,22(3):322-327.
[17] Llorens L, Penuelas J, Beier C, et al. Effects of an experimental increase of temperature and drought on the photosynthetic performance of two ericaceous shrub species along a north-south European gradient[J]. Ecosystems, 2004,7(6):613-624.
[18] Tian Y L, Chen J, Deng A X, et al. Effects of asymmetric warming on the growth characteristics and yield components?of winter wheat under free air temperature increased[J]. Chinese Journal of Applied Ecology, 2011,22(3):681-686.
[19] 房世波,谭凯炎,任三学. 夜间增温对冬小麦生长和产量影响的实验研究[J]. 中国农业科学杂志,2010,43(15):3251-3258.
[20] 王润元,杨兴国,赵鸿,等. 半干旱雨养区小麦叶片光合生理生态特征及其对环境的响应[J]. 生态学杂志,2006,10:1161-1166.
[21] Rawson H M. Effect of high temperatures on the development and yield of wheat and practices to reduce deleterious effects[M]. Klatt A R, eds. Wheat Production Constraints in Tropical Environments. Mexico, D.F, CIMMYT, 1988.44-62.
[22] Zhou J B, Wang C Y, Zhang H, et al. Effect of water saving management practices and nitrogen fertilizer rate on crop yield and water use efficiency in a winter wheat-summer maize cropping system[J]. Field Crops Research, 2011,122(2):157-163.
[23] Ben-Asher J, Garcia A G Y, Hoogenboom G. Effect of high temperature on photosynthesis and transpiration of sweet corn (Zea mays L. var. rugosa)[J]. Photosynthetica, 2008,46(4):595-603.
[24] Rodin J W. Reconciling water-use efficiencies of cotton in field and laboratory[J]. Crop Science, 1992,32:13-18.
[25] Hossain A, DA SILVA J A T. Phenology, growth and yield of three wheat (Triticum aestivum L.) varieties as affected by high temperature stress[J]. Notulae Scientia Biologicae, 2012,4(3):97-109.
[26] Peng S, Huang J, Sheehy J E, et al. Rice yields decline with higher night temperature from global warming[J]. P Natl Acad Sci, USA, 2004,101(27):9971-9975.
[27] Slafer G A, Rawson H M. Base and optimum temperatures vary with genotype and stage of development in wheat[J]. Plant, Cell & Environment, 1995,18:671-679.
[28] Van Ittersum M K, Howden S M, Asseng S. Sensitivity of productivity and deep drainage of wheat cropping systems in a Mediterranean environment to changes in CO2, temperature and precipitation[J]. Agriculture, Ecosystems & Environment, 2003,97(1):255-273.
[29] Wardlaw I F, Moncur L. The response of wheat to high temperature following anthesis. I The rate and duration of kernel filling[J]. Australian Journal of Plant Physiology, 1995,22:391-397.
[30] Asseng S, Foster I, Turner N C. The impact of temperature variability on wheat yields[J]. Global Change Biology, 2011,17:997-1012.
[31] Reynolds M P, Delgado M I, Gutiérrez-Rodrguez M, et al. Photosynthesis of wheat in a warm, irrigated environment. I: Genetic diversity and crop productivity[J]. Field Crops Research, 2000,66:37-50.
[32] 刘军铭,赵琪,尹赜鹏,等. 利用蛋白质组学技术揭示的植物高温胁迫响应机制[J]. 应用生态学报,2015,26(8):2561-2570.
[33] Wang H L, Gan Y T, Wang R Y, et al. Phenological trends in winter wheat and spring cotton in response to climate changes in northwest China[J]. Agricultural and Forests Meteorology, 2008,148:1242-1251.
[34] Parry M L, Swaminathan M S. Effects of Climate Change on Food Production[M]. Cambridge:Cambridge University Press, 1992.
[35] 王鹤龄,王润元,张强,等. 甘肃省作物布局演变及其对区域气候变暖的响应[J]. 自然资源学报,2012,27(3):413-421.
[36] 张彬,郑建初,田云录,等. 农田开放式夜间增温系统的设计及其在稻麦上的试验效果[J]. 作物学报,2010,36(4):620-628.
[37] 田云录,陈金,邓艾兴,等. 非对称性增温对冬小麦籽粒淀粉和蛋白质含量及其组分的影响[J]. 作物学报,2011,37(2):302-308.
[38] Zhang X C, Liu W Z. Simulating potential response of hydrology, soil erosion, and crop productivity to climate change in Changwu tablel and region on the Loess Plateau of China[J]. Agricultural and Forest Meteorology, 2005,131:127-142.
[39] 吴姝,张树源,沈允钢. 昼夜温差对小麦生长特性的影响[J]. 作物学报,1998,24(3):333-337.
[40] 刘振山. 小麦苗期干旱、高温和旱热共胁迫转录表达谱及ABD部分同源基因表达分化分析[D]. 北京:中国农业大学,2015.
[41] Boyer J S, Westgate M E. Grain yields with limited water[J]. Journal of Experimental Botany, 2004,25:2385-2394.
[42] Hasanuzzaman M, Hossain M A, Teixeira da Silva J A, et al. Plant response and tolerance to abiotic oxidative stress: antioxidant defense is a key factor[M]. Bandi V, Shanker A K, Shanker C, et al, eds. Crop Stress and its Management: Perspectives and Strategies. The Netherlands:Springer, 2012.261-315.
[43] Hossain A, Teixeira da Silva J A, Lozovskaya M V, et al. The effect of high temperature stress on the phenology, growth and yield of five wheat (Triticum aestivum l.) genotypes[J]. Asian Australasian J Plant Sci Biotech,2012,6(1):14-23.
[44] McWilliam J R. The national and international importance of drought and salinity effects on agricultural production[J]. Aust J Plant Physiol, 1986,13:1-13.
[45] 刘宗元,张建平,罗红霞,等. 基于农业干旱参考指数的西南地区玉米干旱时空变化分析[J]. 农业工程学报,2014,30(2):105-115.
[46] Beáta Barnabás, Katalin Jger1, Attila Fehér. The effect of drought and heat stress on reproductive processes in cereals[J]. Plant, Cell and Environment, 2008,31:11-38.
[47] 张强,姚玉璧,李耀辉,等. 中国西北地区干旱气象灾害监测预警与减灾技术研究进展及其展望[J]. 地球科学进展,2015,30(2):196-213.
[48] Araus J L, Slafer G A, Reynolds M P, et al. Plant breeding and drought in C3 cereals: what should we breed for[J]. Annals of Botany, 2002,89:925-940.
[49] Bartels D, Souer E. Molecular responses of higher plants to dehydration[M]. Hirt H, Shinozaki K, eds. Plant Responses to Abiotic Stress. Berlin and Heidelberg, Germany, Springer-Verlag, 2004. 9-38.
[50] Zhu X, Gong H, Chen G, et al. Different solute levels in two spring wheat cultivars induced by progressive field water stress at different developmental stages[J]. Journal of Arid Environments, 2005, 62:1-14.
[51] Akram M. Growth and yield components of wheat under water stress of different growth stages[J]. Bangladesh Journal of Agricultural Research, 2011,36(3):455-468.
[52] 任丽雯,马兴祥. 石羊河流域水分胁迫对玉米生长发育指标和产量的影响[J]. 干旱气象,2014,32(5):760-764.
[53] Rampino P, Pataleo S, Gererdi C, et al. Drought response in wheat: physiological and molecular analysis of resistant and sensitive genotypes[J]. Plant, Cell & Environment, 2006,29:2143-2152.
[54] Cominelli E, Galbiati M, Vavasseur A, et al. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance[J]. Current Biology, 2005,15:1196-1200.
[55] Zhou J, Wang X, Jiao Y, et al. Global genome expression analysis of rice in response to drought and high-salinity stresses in shoot, flag leaf, and panicle[J]. Plant Molecular Biology, 2007,63:591- 608.
[56] Battaglia M, Solorzano R.M, Hernandez M, et al. Proline-rich cell wall proteins accumulate in growing regions and phloem tissue in response to water deficit in common bean seedlings[J]. Planta, 2007,225:1121-1133.
[57] Nicolas M E, Lambers H, Simpson R J, et al. Effect of drought on metabolism and partitioning of carbon in two wheat varieties differing in drought-tolerance[J]. Annals of Botany, 1985,55:727-747.
[58] 白莉萍,隋方功,孙朝晖,等. 土壤水分胁迫对玉米形态发育及产量的影响[J]. 生态学报,2004,24(7):1556-1560.
[59] 黄明丽,邓西平,周生路,等. 二倍体、四倍体和六倍体小麦产量及水分利用效率[J]. 生态学报,2007,27(3):1113-1121.
[60] 毕建杰,刘建栋,叶宝兴,等. 干旱胁迫对夏玉米叶片光合及叶绿素荧光的影响[J]. 气象与环境科学,2008,31(1):10-15.
[61] Kobata T, Palta J A, Turner T C. Rate of development of postanthesis water deficits and grain filling of spring wheat[J]. Crop Science, 1992,32:1238-1242.
[62] 赵鸿,李凤民,熊友才,等. 土壤干旱对作物生长发育过程及产量的影响研究进展[J]. 干旱气象,2008,26(3):67-71.
[63] Zhang J, Sui X, Li B, et al. An improved water-use efficiency for winter wheat grown under reduced irrigation[J]. Field Crops Research, 1998,59:91-98.
[64] Yang J C, Zhang J H. Grain filling of cereals under soil drying[J]. New Phytologist, 2006,169:223-236.
[65] 张玉书,米娜. 土壤水分胁迫对玉米生长发育的影响研究进展[J]. 中国农学通报,2012,28(3):1-7.
[66] Turner N C. Crop water deficits:a decade of progress[J]. Adv Agron, 1986,39:1-51.
[67] Machado Stephen, Paulsen G M. Combined effects of drought and high temperature on water relations of wheat and sorghum[J]. Plant and Soil, 2001,23(2):179-187.
[68] Morgan J M. Osmoregulation and water stress in higher plants[J]. Annu Rev Plant Physiol, 1984,35:299-319.
[69] Gunasekera D, Berkowitz G A. Evaluation of contrasting cellular-level acclimation responses to leaf water deficits in three wheat genotypes[J]. Plant Sci, 1992,86:1-12.
[70] 山仑,徐萌. 节水农业及其生理生态基础[J]. 应用生态学报,1991,2(1):70-76.
[71] Craufurd P Q, Peacock J M. Effect of heat and drought stress on sorghum (Sorghum bicolor). II. Grain yield[J]. Experimental Agriculture, 1993,29:77-86.
[72] Savin R, Nicolas M E. Effects of short periods of drought and high temperature on grain growth and starch accumulation of two malting barley cultivars[J]. Australian Journal of Plant Physiology, 1996,23:201-210.
[73] Pradhan G P, Prasad P V V, Fritz A K, et al. Effects of drought and high temperature stress on synthetic hexaploid wheat[J]. Functional Plant Biology, 2012,39(3):190-198.
[74] Shah N, Paulsen G. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat[J]. Plant and Soil, 2003,257:219-226.
[75] Hossain A, da Silva J A T, Lozovskaya M V, et al. High temperature combined with drought affect rainfed spring wheat and barley in South-Eastern Russia: I. Phenology and growth[J]. Saudi Journal of Biological Sciences, 2012,19(4):473-487.
[76] 张凯,王润元,冯起,等. 模拟增温和降水变化对半干旱区春小麦生长及产量的影响[J]. 农业工程学报,2015,31(增刊1):161-170.
[77] Olufayo A, Baldy C, Ruelle P, et al. Diurnal course of canopy temperature and leaf water potential of sorghum (Sorghum bicolor L. Moench) under a Mediterranean climate[J]. Agric For Meteorol, 1993,64:223-236.
[78] Blum A, Ramaiah S, Kanemasu E T, et al. The physiology of heterosis in sorghum with respect to environmental stress[J]. Ann Bot, 1990,65:149-158.
[79] 王石立,娄秀荣. 气候变化对华北地区冬小麦水分亏缺状况及生长的影响[J]. 应用气象学报,1996,7(3):308-315.
[80] 李广,李玥,黄高宝,等. 基于APSIM模型旱地春小麦产量对温度和CO2浓度升高的响应[J]. 中国生态农业学报,2012,20(8):1088-1095.
[81] Aprile A, Havlickova L, Panna R, et al. Different stress responsive strategies to drought and heat in two durum wheat cultivars with contrasting water use efficiency[J]. BMC Genomics, 2013,14(1): 821-838.
[82] Mittler R. Abiotic stress, the field environment and stress combination[J]. Trends in Plant Science, 2006,11:15-19.
[83] Vikender K, Behl R K. Grain yield in wheat as affected by short periods of high temperature, drought and their interaction during pre- and post-anthesis stages[J]. Cereal Research Communications, 2010,38(4):514-520.
[84] 宋松. 幼苗期小麦热激蛋白的诱导研究(英文)[J]. Agricultural Science & Technology, 2010,11(1):28-30.
[85] Ristic Z, Uilliams G, Yang G E, et al. Dehydration, damage to cellular membranes, and heat-shock proteins in maize hybrids from different climates[J]. Journal of Plant Physiology, 1996,149(3-4):424-432.
[86] Slatyer R O. Climatic control of plant water relations[M]. Evans LT, eds. Environmental Control of Plant Growth. Academic Press, New York, NY. 1963.34-54.
[87] Acevedo E H, Silva P C, Silva H R, et al. Wheat production in Mediterranean environments[M]. Satorre E H, Slafer G A, eds. Wheat:Ecology and Physiology of Yield Determination. Food Products Press, Binghamton, NY. 1999,295-331.
[88] Svihra J, Brestic M, Olsovska K. The effect of water and temperature stresses on productivity of winter wheat varieties[J]. Rostlinna Vyroba, 1996,42:425-429.
[89] Saini H S, Westgate M E. Reproductive development in grain crops during drought[M]. Spartes D L, eds. Advances in Agronomy. Academic Press, SanDiego, CA, USA, 2000,68:59-96.
[90] Nouri A, Etminan A, Teixeira da Silva J A, et al. Assessment of yield, yield-related traits and drought tolerance of durum wheat genotypes (Triticum turjidum var. durum Desf.)[J]. Austr J Crop Sci, 2011,5(1):8-16.
[91] Al-Karaki G N. Phenological development-yield relationships in durum wheat cultivars under late-season high-temperature stress in a semiarid environment[J]. ISRN Agronomy, 2012:1101-1107.
[92] Metcalfe D S, Elkins D M, Hughes H D M. Crop production: principles and practices[M]. New York:Macmillan, 1980.
[93] 中国农业百科全书总编辑委员会, 畜牧业卷编辑委员会, 中国农业百科全书编辑部编. 中国农业百科全书(农业气象卷)[M]. 北京:中国农业出版社,1996.
[94] Porter J R, Gawith M. Temperatures and the growth and development of wheat:a review[J]. Eur J Agron, 1999,10:23-36.
[95] Luo Q Y. Temperature thresholds and crop production:a review[J]. Climatic Change, 2011,109(3-4):583-598.
[96] 王建林,林日暖. 中国西部-农业气象灾害(1961-2000)[M]. 北京:气象出版社,2003.
[97] 段若溪,姜会飞. 农业气象学[M]. 北京:气象出版社,2009.
[98] Wheeler T R, Craufurd P Q, Ellis R H, et al. Temperature variability and the yield of annual crops[J]. Agric Ecosyst Environ, 2000,82:159-167.
[99] Dupuis L, Dumas C. Influence of temperature stress on in vitro fertilization and heat shock protein synthesis in maize (Zea mays L.) reproductive systems[J]. Plant Physiol, 1990, 94: 665-670.
[100] Prasad P V V, Boote K J, Allen L H. Adverse high temperature effects on pollen viability, seed-set, seed yield and harvest index of grain-sorghum [Sorghum bicolor (L.) Moench] are more severe at elevated carbon dioxide due to high tissue temperature[J]. Agric For Meteorol, 2006,139:237-251.
[101] 周曙东,周文魁,朱红根,等. 气候变化对农业的影响及应对措施[J]. 南京农业大学学报(社会科学版),2010,10(1):34-39.
[102] 谢云. 粮食产量对气候变化的敏感性分析[J]. 资源科学,1999,21(6):13-17.
[103] 熊伟,居辉,许吟隆,等. 气候变化对中国农业温度阈值影响研究及其不确定性分析[J]. 地球科学进展,2006,21(1):70-76.
[104] 夏江宝,张淑勇,赵自国,等. 贝壳堤岛旱柳光合效率的土壤水分临界效应及其阈值分级[J]. 植物生态学报,2013,37(9):851-860.
[105] 吴元芝,黄明斌. 土壤质地对玉米不同生理指标水分有效性的影响[J]. 农业工程学报,2010,26(2):82-88.
[106] Nable R O, Robertson M J, Berthelsen S. Response of shoot growth and transpiration to soil drying in sugarcane[J]. Plant Soil, 1999,207:59-65
[107] 张喜英,裴冬,由懋正. 几种作物的生理指标对土壤水分变动的阈值反应[J]. 植物生态学报,2000,24(3):280-283.
[108] 陈家宙,王石,张丽丽,等. 玉米对持续干旱的反应及红壤干旱阈值[J]. 中国农业科学,2007,40(3):532-539.
[109] Lacape M J, Wery J, Annerose D J M. Relationships between plant and soil water status in five field-grown cotton (Gossypium hirsutum L.) cultivars[J]. Field Crops Research, 1998,57:29-43.
[110] Denmead O T, Shaw R H. Availability of soil water to plants as affected by soil moisture content and meteorological conditions[J]. Agronomy Journal, 1962,45:385-390.
[111] Ray J D, Gesch R W, Sinclair T R, et al. The effect of vapor pressure deficit on maize transpiration response to a drying soil[J]. Plant and Soil, 2002,239(1):113-121.
[112] Novák V, Havrila J. Method to estimate the critical soil water content of limited availability for plants[J]. Biologia, 2006,61(19):S289-S293.
[113] Ghosh A, Majumder A. Incidence of drought in relation to rainfed rice production: An analysis[J]. International Journal of Bio-resource and Stress Management, 2014,5(1):113-115.
Progress and Perspectives in Studies on Responses and Thresholds of Major Food Crops to High Temperature and Drought Stress
ZHAO Hong1, WANG Runyuan1, SHANG Yan2, WANG Heling1,ZHANG Kai1, ZHAO Funian1, QI Yue1, CHEN Fei1
(1.KeyLaboratoryofAridClimaticChangeandReducingDisasterofGansuProvince,KeyLaboratoryofAridclimateChangeandDisasterReductionofCMA,InstituteofAridMeteorology,ChinaMeteorologicalAdministration,Lanzhou730020,China; 2.CollegeofLifeScienceandTechnology,GansuAgricultureUniversity,Lanzhou730070,China)
Abstract:In recent years, climate change with the main indicator of air temperature increasing and precipitation fluctuations had an important impact on agricultural production. Researches about the responses of crop growth, morphogenesis, physiological and biochemical process to temperature increase and water decrease are not only play an important role in revealing mechanism about impact of climate change on crops, but also an important prerequisite of making counter measures. In this paper, the effects of temperature increase and water decrease and their combined influence on the development, physiological ecology and yields of main grain crops like rice, wheat and maize etc at home and abroad in recent years were reviewed, and their change process, thresholds and the consequences from change of temperature and water were discussed. At the same time, some current problems in these studies were also found. In addition, based on the hot study issues about influence and their thresholds of warmer temperature and water stress on crops at present, some key scientific problems were proposed in the future study, including:the thresholds of temperature and water under drought/wet conditions, and the endurance limit of crops under multi-factors collaborative stress; the relationships between stress level, time, duration and crop physiological, biochemical processes, as well as the cellular and molecular level response mechanism; the compensation effect of crops to moderate drought at higher temperature.
Key words:food crops; high temperature; drought stress; development; physiological ecology; yields; thresholds
中图分类号:S51
文献标识码:A
文章编号:1006-7639(2016)-01-0001-12
doi:10.11755/j.issn.1006-7639(2016)-01-0001
作者简介:赵鸿(1977-),女,甘肃临洮人,副研究员,博士,主要从事干旱气候变化与农业生态等方面的研究. E-mail:zhaohonglt@126.com
基金项目:国家自然科学基金(41505099, 41275118)及公益性行业(气象)科研重大专项(GYHY201506001-2)共同资助
收稿日期:2015-12-13;改回日期:2016-02-05
赵鸿,王润元,尚艳,等.粮食作物对高温干旱胁迫的响应及其阈值研究进展与展望[J].干旱气象,2016,34(1):1-12, [ZHAO Hong, WANG Runyuan, SHANG Yan, et al. Progress and Perspectives in Studies on Responses and Thresholds of Major Food Crops to High Temperature and Drought Stress[J]. Journal of Arid Meteorology, 2016, 34(1):1-12], doi:10.11755/j.issn.1006-7639(2016)-01-0001
