滆湖大型底栖动物群落分布和氮磷因子的相关分析
2016-04-18陈志宁张红高周崴沈丽娟
陈志宁,张红高,周崴,沈丽娟
(1.常州市环境监测中心,江苏 常州 213014;2.江苏省环境保护水环境生物监测重点实验室,江苏 常州 213014)
·解析评价·
滆湖大型底栖动物群落分布和氮磷因子的相关分析
陈志宁1,2,张红高1,2,周崴1,2,沈丽娟1,2
(1.常州市环境监测中心,江苏 常州 213014;2.江苏省环境保护水环境生物监测重点实验室,江苏 常州 213014)

大型底栖动物;氮;磷;时空分布;滆湖
底栖动物是水生生态系统食物链的重要组成部分,影响着物质分解和营养循环,有相对固定的栖息地且生命周期较长,其不同种类对水质的敏感性差异大,在受外界干扰后生物群落的变化(如群落多样性的减少、优势种群个体体型的变化)可反映水体受污染的性质和程度,因此可以有效地指示水质的生态状况。底栖动物,尤其是水栖昆虫,已广泛应用于各种环境污染的指示,如有机污染[1]、重金属污染[2]、水生生态系统退化[3-4]等。欧盟《水框架指令》和美国EPA也都明确地将底栖动物列为指示生物。
现于2012年12月—2013年11月对滆湖各湖区进行采样调查,初步了解滆湖大型底栖动物的空间分布特征及其同氮磷元素之间的关系,为滆湖水域生态学的研究提供基础资料。
1 研究方法
1.1 采样点布设
滆湖(31°29′—31°42′N,119°44′—119°53′E)是苏南地区仅次于太湖的第二大淡水湖,位于太湖流域西部。现有水面积16 600 hm2,常年平均水深约1.3 m。根据滆湖的形状及周边环境,在滆湖设采样点20个,分别为:S1 ~ S20。
1.2 样品采集与处理
2012年12月—2013年11月对滆湖各采样点进行大型底栖动物采样,样品采集利用1/16 m2采集面积的彼得逊采泥器,经60目尼龙筛洗净后置于白色搪瓷盘中进行底栖动物的挑选。样品的保存、计数、称重等按文献[5]操作。
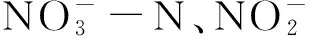

1.3 数据分析
TN、TP时空分布采用surfer 9.0进行绘图,空间插值采用普通克里格(Kriging)插值法;运用典范对应分析(canonical correspondence analysis,简称CCA)分析环境因子与底栖群落结构的关系,在Canoco for windows 4.5软件包上进行[6],CCA分析时底栖动物数据进行平方根转换,环境因子进行lg(x+1) 转换(pH值除外)。
2 结果分析
2.1 滆湖水质特征及TN、TP时空分布

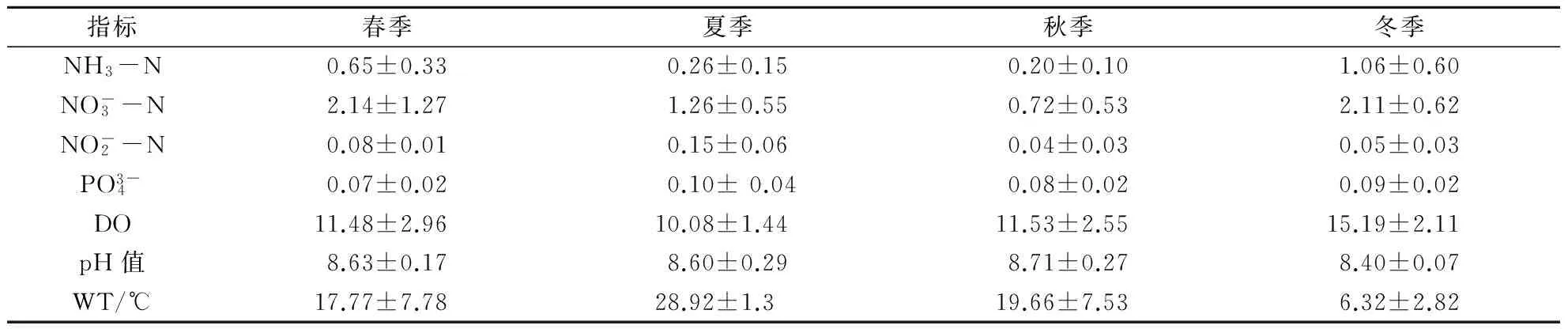
表1 滆湖理化指标 mg/L
滆湖水体TN四季的空间分布见图1。由图1可见,滆湖水体TN值除秋季以外,都已超过地表水Ⅴ类水质标准,其中冬季水体的TN值高于其他季节。湖北区和湖南区水体TN值差异显著,湖北区的TN值显著高于湖南区,且湖区TN值从北至南有递减的趋势,其中秋季的湖南区基本可达Ⅲ类水标准。
滆湖TP四季的空间分布见图2。滆湖TP值在春季、夏季、秋季、冬季均已超过地表水Ⅳ类水标准。其中,夏季水体的TP值相对较高,其他季节的TP值差异不大。与TN值的空间分布规律类似,TP值也呈现出湖北区高于湖南区的规律。
2.2 大型底栖动物的种类与数量
调查共获得大型底栖动物15种,隶属于3门8科13属。其中,环节动物有2科3属5种,占总种数的33.3%,软体动物有2科2属2种,占总种数的13.3%,节肢动物有4科8属8种,占总种数的53.3%,见表2。
由表2可见,大型底栖动物在出现区域的标准差较大,如中国长足摇蚊,总均丰度标准差为399,而总均丰度仅为130,此外,克拉伯水丝蚓与贝蠓仅在一个样点出现,标准差为0。
2.3 主要优势种的时空分布
滆湖底栖动物中霍甫水丝蚓和中国长足摇蚊具有较高的出现率,是主的要优势种。滆湖大型底栖动物优势种的时空分布情况见图3(a)(b)(c)(d)。[(b)(c)单位为102m-2)]
由图3可见,霍甫水丝蚓的丰度在时间上的变化显著,总体表现为秋季>夏季>冬季>春季,且在夏季的出现频率最高,冬季的出现频率最低;霍甫水丝蚓在空间上无显著的变化规律。

图1 滆湖水体TN四季的空间分布特征

图2 滆湖水体TP四季的空间分布特征

表2 滆湖各类大型底栖动物的出现率和丰度

续表

图3 滆湖大型底栖动物优势种四季分布
中国长足摇蚊在时间上的分布规律与霍甫水丝蚓类似,其丰度表现为秋季>夏季>冬季>春季,且夏季的出现频率最高,冬季的出现频率最低。站点S8的中国长足摇蚊丰度全年都显著较高,变化范围为104~2 272 m-2,其中春季丰度最低,秋季最高。
2.4 大型底栖动物的分布与水体氮磷的关系
对滆湖氮磷营养元素及基本理化指标与底栖动物群落结构进行CCA分析,CCA排序见图4。前2个排序轴共解释了整个底栖动物群落结构变异程度的53.0%,其中Axis1解释了37.7%。
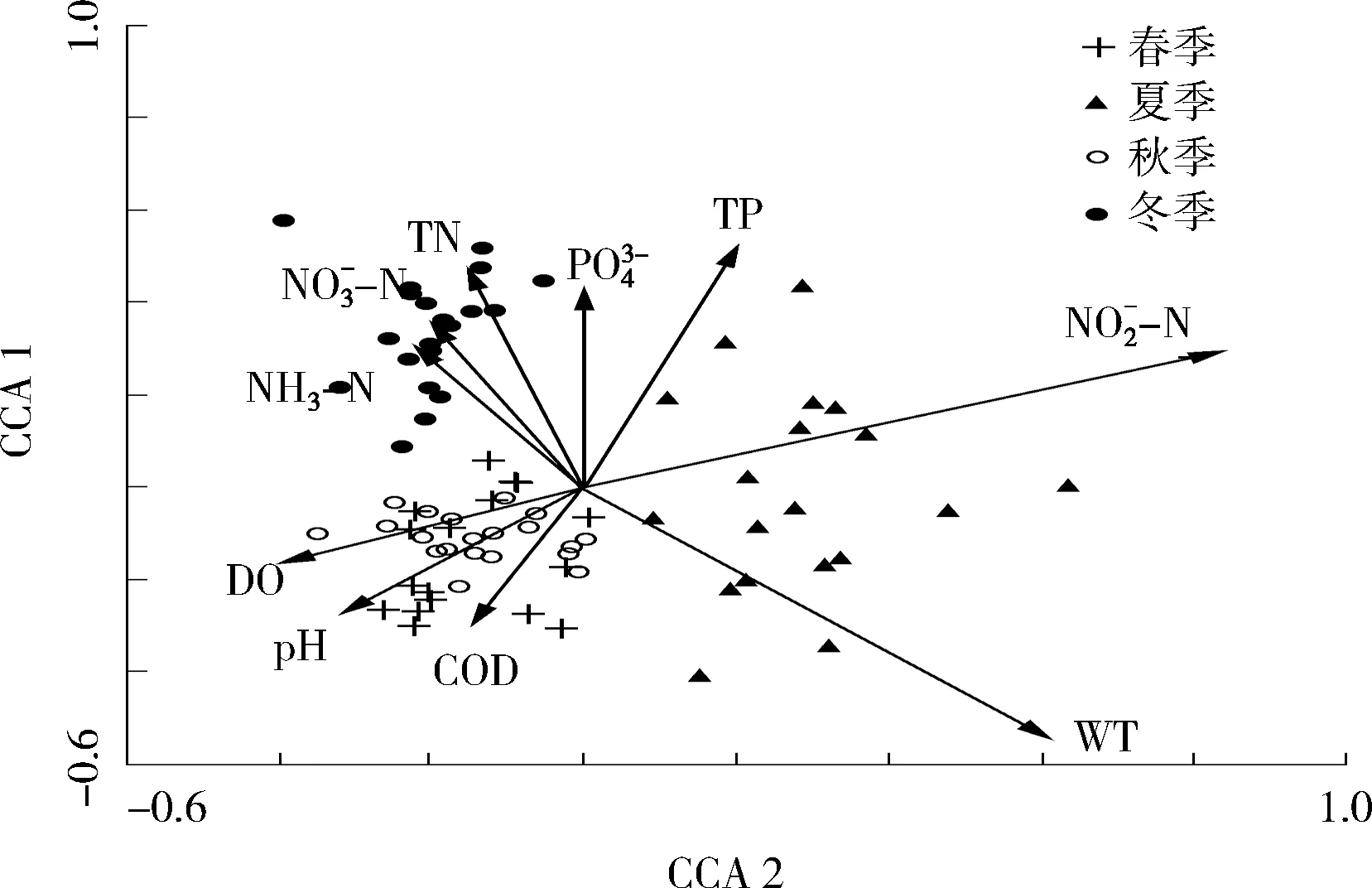
图4 滆湖大型底栖动物与环境因子的CCA 排序


表3 优势种密度与水体氮磷浓度的Pearson相关性分析①
①**:p<0.01;*:p<0.05。
3 讨论
TN、TP是滆湖的主要污染物[7-8],滆湖水体的氮磷主要来源于:(1)滆湖周边的工业企业污水处理不到位,包括印染、化工企业等;(2)滆湖上游大部分乡镇污水收集系统落后导致生活污水的排入;(3)农业面源污染严重[9];(4)人工养殖投饵量的增加。TN、TP在空间上呈现北高南低的规律,可能与滆湖上游快速发展的工业企业有关,此外,上游非武进地区的污染排放也是重要的影响因素[10]。TN在冬季浓度较高这一规律与太湖类似,滆湖为浅水湖泊,冬季风多且大对沉积物有扰动,促进沉积物中氮的释放,另外,冬季水生植物生长缓慢,对氮的利用率较低,也可能引起氮含量的升高。
滆湖属于施肥养殖湖泊兼具城市湖泊的特征,氮磷污染严重。湖泊的富营养化程度对底栖动物群落结构的优势种、丰度、生物量等均存在显著影响[11]。调查发现颤蚓类和摇蚊科幼虫是主要的特征种类(包括霍甫水丝蚓,巨毛水丝蚓,克拉伯水丝蚓和苏氏尾鳃蚓),与前人的研究结果相符。与以往对滆湖底栖动物的调研数据对比,所得物种种类显著降低。2002—2003年采集的滆湖底栖动物有31种[12],2009—2010年报道的滆湖底栖动物有35种[13]。究其原因,湖泊水体的污染、富营养化的加剧是导致底栖动物多样性急剧降低的重要因素[14]。滆湖水体的主要优势种为霍甫水丝蚓和中国长足摇蚊,其出现率分别达到71.3%和53.8%。霍甫水丝蚓是寡毛类常见的物种,耐污性较强,在低氧环境下可以正常生长繁殖,常被用作有机污染或富营养化的标志性指示种[15]。有研究表明,霍甫水丝蚓丰度与湖泊富营养化程度有关,有随着富营养程度的增加而增加的趋势[16-17]。而研究中霍甫水丝蚓与总氮、硝酸盐氮关系呈负相关,与这一结论不符,可能与污染的浓度范围有关。调查中,北部湖区与南部湖区的氮磷浓度虽有显著的差异,但霍甫水丝蚓在空间上无显著的差异,因为随着湖泊深度的增加以及过高的污染物浓度,霍甫水丝蚓的生长繁殖也有可能会受到一定程度的抑制[18]。龚志军等[17]认为中国长足摇蚊可作为超富营养水体的指示种,适合在一定程度的超富营养化环境中生存。站点S8的中国长足摇蚊丰度显著高于其他站点,但该站点处于湖南区,氮磷浓度较低,造成这种差异性的原因有多种可能性,底栖动物的分布除了受氮磷营养元素的控制,区域因素和空间位置等都是不可忽视的环境因素。

4 结论
(1)滆湖各湖区水体氮磷污染严重。ρ(TN)在时间上表现为冬季>春季>夏季>秋季,ρ(TP)在时间上表现为夏季>春季、冬季>秋季;在空间上,北部湖区的TN、TP值均高于南部湖区,从北至南有递减的趋势;
(2)滆湖大型底栖动物出现频率最高的为霍甫水丝蚓和中国长足摇蚊,分别为71.3%和53.8%。霍甫水丝蚓和中国长足摇蚊的丰度在时间上的变化显著,均表现为秋季>夏季>冬季>春季,在空间上的规律性不明显;
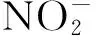
[2] SMOLDERS A J P,LOCK R A C,VAN DER VELDE G,et al.Effects of mining activities on heavy metal concentrations in water,sediment and macroinvertebrates indifferent reaches of the Pilcomayo River,South America[J].Archives of Environmental Contamination and Toxicology,2003,44(3):314-323.
[3] LORENZ A,HERING D,FELD C K,et al.A new method for assessing the impact of hydromorphological degradation on the macroinvertebrate fauna of five German stream types [J].Hydrobiologia,2004,516(1-3):107-127.
[4] FRIBERG N,SANDIN L,PEDERSEN M L.Assessing the effects of hydromorphological degradation on macroinvertebrate indicators in rivers:examples,constraints,and outlook[J].Integrated Environmental Assessment and Management,2009,5(1):86-96.
[5] GROWNS J,CHESSMAN B,JACKSON J,et al.Rapid assessment of Australian rivers using macroinvertebrates:cost and efficiency of 6 methods of sample processing[J].Journal of the North American Benthological Society,1997,16:682-693.
[6] 冯素珍,李畅游.基于水生生物的湖泊营养类型评价[J].环境监测管理与技术,2010,22(2):228-232.
[5] 国家环境保护总局.水和废水监测分析方法[M].4版.北京:中国环境科学出版社,2012.
[6] BRAAK C J F T,SMILAUER P.CANOCO reference manual and user′s guide to canoco for windows:software for canonical community ordination[M].Version 4.Centre for Biometry,1998.
[7] 王兆群,张宁红,张咏,等.洪泽湖水质富营养化评价[J].环境监控与预警,2010,2(6):31-35.
[8] 韦忠,王晓杰.滆湖氮磷来源及总量分析[J].治淮,2013(1):92-93.
[9] 吴云波,郑建平.滆湖入湖污染物控制对策研究[J].环境科技,2010,23(1):12-14.
[10] 汪院生.滆湖水环境演变及其原因分析[J].水利规划与设计,2013(8):37-40.
[11] 蔡永久,姜加虎,张路,等.长江中下游湖泊大型底栖动物群落结构及多样性[J].湖泊科学,2010,22(6):811-819.
[12] 刘其根,孔优佳,陈立侨,等.网围养殖对滆湖底栖动物群落组成及物种多样性的影响[J].应用与环境生物学报,2005,11(5):566-570.
[13] 王丽卿,吴亮,张瑞雷,等.滆湖底栖动物群落的时空变化及水质生物学评价[J].生态学杂志,2012,31(8):1990-1996.
[14] 熊金林,梅兴国,胡传林.不同污染程度湖泊底栖动物群落结构及多样性比较[J].湖泊科学,2003,15(2):160-168.
[15] ASTON R.Tubificids and water quality:a review[J].Environmental Pollution,1973,5:1-10.
[16] 李艳,蔡永久,秦伯强,等.太湖霍甫水丝蚓(LimnodrilushoffmeisteriClaparède) 的时空格局[J].湖泊科学,2012,24(3):450-459.
[17] 龚志军,谢平,唐汇涓,等.水体富营养化对大型底栖动物群落结构及多样性的影响[J].水生生物学报,2001,25(3):210-216.
[18] VOLPERS M,NEUMANN D.Tolerance of two tubificid species (TubifextubifexandLimnodrilushoffmeisteri) to hypoxic and sulfidic conditions in novel,long-term experiments[J].Archivfür Hydrobiologie,2005,164:13-38.
[19] 邵卫伟,张勇,于海燕,等.不同土地利用对溪流大型底栖无脊椎动物群落的影响[J].环境监测管理与技术,2012,24(3):18-23.
[20] 赵升,刘旭东,张爱君,等.黄岛电厂温排水对大型底栖生物群落的影响[J].环境监测管理与技术,2013,25(4);18-23.
[21] 渠晓东,曹明,邵美玲,等.雅砻江(锦屏段)及其主要支流的大型底栖动物[J].应用生态学报,2007,18(1):158-162.
[22] 季相星,姜毅,王晨波,等.连云港市泥砂质潮间带大型底栖动物群落调查[J].环境监控与预警,2014,6(6):41-44.
[23] SANDIN L,JOHNSON R K.Local,landscape and regional factors structuring benthic macroinvertebrate assemblages in Swedish streams[J].Landscape Ecology,2004,19(5):501-514.
[24] COURTNEY L A,CLEMENTS W H.Effects of acidic pH on benthic macroinvertebrate communities in stream microcosms[J].Hydrobiol,1998,379:135-145.
[25] 汪星,郑丙辉,刘录三,等.洞庭湖典型断面底栖动物组成及其与环境因子的相关分析[J].中国环境科学,2012,32(12):2237-2244.
[26] DAUER D M,RANASINGHE J A,WEISBERG S B.Relationships between benthic community condition,water quality,sediment quality,nutrient loads,and land use patterns in Chesapeake Bay[J].Estuaries,2000,23(1):80-96.
[27] DÉVAI G,MOLDOVN J.An attempt to trace eutrophication in a shallow lake using chironomids[J].Paleolimnology,1983,15:169-175.
[28] 段学花,王兆印,余国安.以底栖动物为指示物种对长江流域水生态进行评价[J].长江流域资源与环境,2009,18(3):241-247.
[29] 刘玉,VERMAAT J E,RUYTER ED,等.珠江、流溪河大型底栖动物分布和氮磷因子的相关分析[J].中山大学学报:自然科学版,2003,42(1):95-99.
[30] ASHTON M J,MORGAN R P,STRANKO S.Relations between macroinvertebrates,nutrients,and water quality criteria in wadeable streams of Maryland,USA[J].Environmental Monitoring and Assessment,2014,186:1167-1182.
Analysis on the Relationship between the Distribution of Macrobenthos Community and Nitrogen-Phosphorus Factor in the Gehu Lake
CHEN Zhi-ning1,2,ZHANG Hong-gao1,2,ZHOU Wei1,2,SHEN Li-juan1,2
(1.ChangzhouEnvironmentalMonitoringCenter,Changzhou,Jiangsu213014,China;2.JiangsuProvincialEnvironmentalProtectionKeyLaboratoryofWaterEnvironmentalBiologicalMonitoring,Changzhou,Jiangsu213014,China)

Macrobenthos; Nitrogen; Phosphorus; Spatial and temporal variation; Gehu Lake
2015-04-03;
2015-05-13
江苏省环境监测基金资助项目(1313);国家科技重大专项基金资助项目(2012ZX07506-003,2012ZXO7101-007)
陈志宁(1983—),男,工程师,本科,从事环境监测与研究工作。
X826
B
1674-6732(2016)01-0045-06
