野生大熊猫种群数量调查方法研究进展
2016-04-14史雪威张晋东欧阳志云
史雪威,张晋东,欧阳志云
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085 2 西华师范大学生命科学学院, 南充 637009 3 中国科学院大学, 北京 100049
野生大熊猫种群数量调查方法研究进展
史雪威1,3,张晋东2,欧阳志云1,*
1 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085 2 西华师范大学生命科学学院, 南充 637009 3 中国科学院大学, 北京 100049
大熊猫(Ailuropodamelanoleuca)是中国特有珍稀野生动物,被誉为野生动物保护领域的“旗舰物种”。大熊猫种群数量的变化可以实时反应大熊猫种群动态,提供大熊猫分布区域、栖息地质量等最直接的信息,是制定大熊猫保护方案的基础,也是有效实施大熊猫保护措施的前提。综述了几种传统野生大熊猫种群数量调查方法(包括直接计数法、数学模型法、距离-咬节分析法及分子生物学方法),以及近年来最新应用于野生动物种群数量调查的红外相机技术、足迹鉴定法,讨论了传统方法中可能存在的问题、分析新方法的应用前景,并针对今后的野生大熊猫种群数量调查提出了一些建议与展望。
大熊猫;种群数量调查方法;距离-咬节分析法;分子生物学方法;红外相机技术;足迹鉴定法
种群数量是动物生态学、行为生态学和资源管理学最核心的问题之一,通过种群数量调查可以有效的掌握种群动态、种群生活史及进化对策、种群栖息地状况[1]。种群的婚配制度、繁殖策略、繁殖行为的表达也与种群数量调查密切相关[2]。种群数量的变化可以评价濒危物种的保护措施对种群恢复的作用,资源管理部门亦可以依据种群数量的调查结果制订有效的保护管理计划。
大熊猫(Ailuropodamelanoleuca)是中国特有珍稀野生动物,有“国宝”和“活化石”之称,被誉为野生动物保护领域的“旗舰物种”,历史上,大熊猫曾广泛分布于我国南部、中部、西南,向北直到北京周口店等,向南至缅旬北部[3]。然而由于地质、气候的变迁,特别是近代以来人类活动范围的不断扩大和对森林资源的过度利用,大熊猫面临种群隔离、遗传多样性降低以致灭绝的危险[4]。
大熊猫种群数量调查可以实时反应大熊猫种群动态变化,提供大熊猫分布、栖息地状况等最直接的信息,是制定及有效实施大熊猫保护措施的前提。为得到大熊猫种群数量等信息,建国以来,国家林业局先后组织4次全国大熊猫调查,最近的调查结果显示截至2013年底,全国野生大熊猫种群数量达1864只(表1),比第三次调查增加268只,增长16.8%[5]。

表1 历次全国大熊猫调查总结
大熊猫生活在高山峡谷的密林竹丛中,栖息地地形复杂;同时大熊猫种群密度低,其行动也十分隐蔽,对人类的警惕性很高,在野外很难观察到它的实体,因此许多陆栖动物的数量调查方法不能运用于大熊猫种群数量调查[6]。为了得到野生大熊猫数量及分布的确切数据,科研工作人员先后采用直接计数法、数学模型法、路线调查法、距离区分法和咬节区分法、分子生物学方法等多种方法对野生大熊猫种群数量进行调查。历次大熊猫种群数量调查均花费高额费用,调动大量人力、物力参与其中,但由于种种原因,其所得结果常为后人所质疑。目前大熊猫种群调查方法依然缺乏一种同时具备客观性、非损伤性、精确度高、投入产出比高、可实践性强大熊猫种群调查方法。本文对大熊猫种群数量调查方法进展进行综述,梳理野生大熊猫种群数量调查方法,讨论方法中可能存在的问题,并探讨了今后的野生大熊猫种群数量调查方法的发展方向。
1 野生大熊猫种群数量调查方法
野生大熊猫种群数量调查方法大体上可分为两种:直接统计方法和间接统计方法。
1.1 直接统计方法
直接统计方法主要包括直接计数法和数学模型法,其核心思想是通过布设调查路线、样方等直接对野生大熊猫直接进行观察,根据实际观察到的野生大熊猫数量估算样方内密度,进而推算研究区域内总体数量。
1.1.1 直接计数法
1968年,我国首次野外大熊猫调查,采用直接计数法(又名哄赶法或围赶法)调研了四川省平武县王朗自然保护区的大熊猫数量[7]。调查步骤如下:(1)调查目标动物的主要栖息地并选好调查样地;(2)在样地内对目标动物进行哄赶并计数;(3)计算调查样地内目标动物栖息密度;(4)根据样地占研究区面积比例,推算研究区域内目标动物种群数量。
王朗自然保护区大熊猫调查组选择两条比较小又具有代表性的山沟(机械工棚沟和澡堂沟)作为样地,分两路携带猎犬由沟底小溪两侧向上搜索驱赶,间隔一定距离统计所发现大熊猫实体数量,依据样地面积,计算大熊猫在有林地面积和箭竹丛面积中的栖息密度,然后根据有林地面积和箭竹丛面积所占保护区的百分比,推算出保护区大熊猫的种群数量。
直接计数比较原始,数量统计精确度也较低,仅适用于对大型动物的调查,例如有蹄类动物的数量[8],能在一定意义上说明该区域存在野生大熊猫。直接计数法人力物力花费较大,对大熊猫的惊扰较严重,野外观测时存在重复计数和漏计问题,现该方法已被淘汰[6]。
1.1.2 数学模型法
1988年,北京大学、陕西长青林业局联合大熊猫研究小组在出版的《秦岭大熊猫的自然庇护所》一书中提出了用数学模型推导在针阔叶混交林-巴山木竹林-大熊猫生态系统中大熊猫种群密度的方法[9]。模型为:
式中,P为单位面积内熊猫的平均数量,即密度(只/km2);T为单位面积内所设立的一个样方的面积(km2);V为一个观测时间单位中,在一个样方里观察到熊猫的平均数量(只·样方面积-1·观测时间-1);K为在一个观测时间中,一只熊猫可能独立进入一个样方的次数。

式中,T值选取依据竹林中能见度的最大范围,K值根据大熊猫在观测时间内的移动范围与样方面积的比值得出。
为了验证此模型的精确性,调查组利用数学模型法于1985年对佛坪自然保护区三官庙地区、1986年对长青林业局华阳林场境内的大熊猫数量进行调查,得出与以往调查资料相近的结果。
数学模型法是一种简单易行的种群调查方法,调查人员只需记录路线长度、样方数、直观熊猫数等数据即可依据模型估算区域内熊猫数量。但野外直接观察到大熊猫实体十分困难,而且大熊猫往往会通过敏锐的嗅觉、听觉率先发现调查者并改变原有的行进路线,这种行进路线的改变是否会造成直观大熊猫重复计数有待于进一步的调查。另外,在大熊猫的不同生境下,竹林密度会有所差异,因此竹林中能见度的最大范围相应不同,即T值不同。模型参数较多为推测,模型存在一定的模糊性和不确定性。
1.2 间接统计方法
由于大熊猫栖息地地形复杂,大熊猫活动隐秘,不容易直接定位、观察野生大熊猫个体,目前对野生大熊猫的监测、数量调查工作常通过对大熊猫活动产生的衍生物(如粪便等)进行分析,间接统计大熊猫数量。间接统计方法包括路线调查法、距离分析法和咬节区分法以及分子生物学调查法。
1.2.1 路线调查法
路线调查法在大熊猫调查中有着较长时间的应用,1974年—1977年开展的四川省大熊猫等珍稀动物调查(即全国第一次大熊猫调查)采用了路线法。路线调查法简称路线法,又名样线法,是一种在预先划定的调查区域内布设调查路线收集大熊猫活动各种信息的方法,适用于分布面积广、无法进行彻底调查的动物[10]。调查步骤如下:(1)通过查阅资料、走访当地村民等确定重点调查区和一般调查区;(2)根据调查区域内地形地貌、植被状况、竹类分布状况、大熊猫的生态习性等划分调查小区,布设调查路线;(3)调查人员依据调查路线收集、记录大熊猫活动的各种信息[3]。调查路线布设的原则是:保证调查路线尽可能多地穿过大熊猫活动的各种生境(包括沟谷、山坡、山梁等),收集的信息能够有效代表该调查小区[11]。
全国第一次大熊猫调查过程中,调查人员在岷山、邛崃山、凉山、相岭及秦岭南坡中段等大熊猫栖息地内,按“V”字形进行搜索,记录遇到的大熊猫实体以及各种新鲜活动痕迹(如粪便、咬痕、卧穴、足迹等)。统计分析时,只以新鲜粪便为指标(陈旧粪便不作统计),将新鲜粪便的直径、组成成分、消化状况、咬节长度等作为区分大熊猫个体的重要依据。指标相同则视为同一只个体,否则视为不同个体[12]。在这次调查中,咬节长度虽然作为区分大熊猫个体的指标之一得到应用,但区分咬节长度的判别标准较为粗糙(没有设置区分阈值),后经完善逐步形成了系统的咬节区分法。
路线调查法是一种原理简单、容易推广和易于操作的方法,同时也是是其他调查方法的基础。通过路线调查法可以收集到野生大熊猫的相关信息,这些信息不但可以用于分析大熊猫的种群数量和分布范围,还囊括了大熊猫栖息地及潜在栖息地受干扰状况。但第一次全国大熊猫调查中应用的路线法过多依靠调查人员的主观视觉评价(例如对大熊猫实体大小、粪便新鲜程度的判断等),不同调查人员之间调查结果存在较大差异,所得数量仅供参考。
大熊猫调查工作目前细化分为外业调查方法和内业分析方法。在现阶段研究工作中,路线调查法常作为外业调查方法一种,收集所需要的资料。全国第三次大熊猫调查外业调查即采取拉网式路线调查法,首要搜集大熊猫的粪便,作为分析大熊猫种群数量的依据[11]。
1.2.2 距离区分法和咬节区分法
1999—2003年全国第三次大熊猫调查外业调查采取拉网式路线调查法收集大熊猫活动的各种信息,而内业分析方法采用距离区分法和咬节区分法结合综合分析大熊猫种群数量。内业分析方法具体为:(1)首先利用距离区分法初步区分。通过GIS软件计算出任意两处痕迹点之间的距离,将此距离与区分阈值进行比较,如果距离大于区分阈值,则将其区分为两个不同个体。(2)再利用咬节区分法进一步区分。对于小于距离区分阈值的痕迹点集合,则计算不同痕迹点的咬节平均值之差,通过与咬节区分阈值比较,进行个体识别(图1)[11]。
距离区分法是指当两个痕迹点之间的间距超出一定范围时,可基本判断这两个痕迹点不属于同一只大熊猫。大熊猫属于独栖动物,除繁殖季节外,野生大熊猫间常保持着一定距离,每只大熊猫通过嗅觉主动回避其他个体的近日活动范围[13],除了发情期和季节性垂直迁移期及突发事件(如冷箭竹开花、地震)外,野生大熊猫基本都在一个有限的区域内摄食、饮水或寻找配偶,这个区域即巢域(家域)[14],如果两个痕迹点间距不在这个范围内,那么它们属于同一只大熊猫的概率就变得很低。另外大熊猫为节省能量消耗不爱走动,日移动直线平均距离不到500m[3]。由于大熊猫在一定时间内的移动距离是相对稳定的,大熊猫很可能只出现在一个以某一痕迹点为圆心,该时间段内大熊猫最远移动距离为半径的区域内。因此如果两个痕迹点之间的距离超过发现这两个痕迹点的时间间隔段内大熊猫最远移动距离(距离区分阈值),基本上可以判断这两个痕迹点不属于同一只大熊猫所留。全国第三次大熊猫调查中采用的距离区分阈值分别为:1d为D1=1.0km;1—3d为D3=1.5km;4—15d为D7=2.5km;15d以上为DMAX=3.5km[11]。
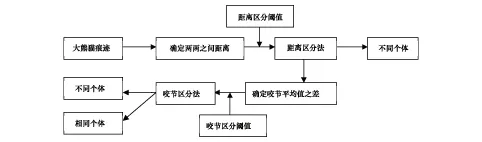
图1 第三次全国大熊猫调查大熊猫种群数量分析流程图[11]Fig.1 The flow chart of Giant Panda population analysis in The Third Nation′s Giant Panda Survey[11]
采用距离区分法时,对于大尺度上距离显著大于区分阈值的两处痕迹点是很容易区分开来的,但是对于在小尺度上距离小于或接近区分阈值的两处痕迹点的区分能力不强。Zhang等对陕西省佛坪自然保护区的4只野生大熊猫GPS追踪发现,大熊猫冬季巢域范围显著大于夏季巢域范围,并且不同的野生大熊猫家域之间存在很高程度的巢域重叠现象[15]。距离区分阈值同日行走距离密切相关,而大熊猫日行走距离受繁殖因素、寻找食物难易程度等多方面因素影响,不同个体、季节有很大差异,日行走距离很难有一个确定的数字[16]。胡锦矗等通过无线电监测测定四川卧龙保护区大熊猫每天活动平均约为600—1500m左右,移动的直线距离不超过500m;潘文石等通过无线电项圈对秦岭6只大熊猫日常活动进行监测,发现任何个体在同一个时间间隔内的迁移距离都存在很大差别,并且30d内同一个体迁移距离随时间间隔的增加其平均距离也增大[10];Zhang等对卧龙自然保护区5只野生大熊猫的监测显示大熊猫日活动范围在(241±341)m。目前大熊猫移动距离、巢域大小等多利用无线电监测进行,通过判断信号强弱以确定目标个体的方位。但山区地形复杂、植被丰富,无线电信号反射现象较为严重,而且定位目标个体方位易受监测者主观意识影响,因此可能存在较大的定位误差[4],现在GPS追踪的应用揭示了比无线电项圈更广的大熊猫家域和活动核心区域[16,17]。距离区分阈值常采用调查时期内的大熊猫平均最远移动距离,因此分析出的大熊猫种群数量是一个保守的数字。另外,大熊猫在空间利用上存在显著的互相影响。Hull等发现1个熊猫对(由1只雄性和两只雌性大熊猫组成)的叠合巢域同时显著吸引熊猫对里的3只大熊猫(即熊猫对里3只大熊猫会同时出现在他们的叠合巢域内)[17]。仅通过痕迹点之间的距离就判断是否为1只大熊猫值得商榷,大熊猫行为生态学的进一步发展将有助于距离区分法的改进与应用。
20世纪80年代以来,咬节区分法作为判断大熊猫种群数量的重要方法之一得到了广泛的应用。咬节(Bamboo Stem Fragments)即为大熊猫粪便中未消化的竹茎,普遍认为具有个体特征[3]。对粪便中的咬节长度进行多元方差分析,可以区分相邻样线上不同个体,进而得出区域内大熊猫数量。尹玉峰等提出目前使用的咬节区分法阈值(2mm)可较准确判断大熊猫种群数量[18]。
用咬节区分个体的前提是在野外,大熊猫彼此间隔离,区域范围内数量很少的情况下进行。在较大区域上,由于会存在更多的年龄相同或相仿、咀嚼能力接近的大熊猫个体,彼此之间咬节平均长度有可能差异不显著[3]。同时大熊猫食性存在季节性变化,在某些季节全采食竹叶而在其粪便中不留下咬节,因此咬节区分法具有一定时间局限性[19]。另外,咬节区分法判断两只个体所使用的平均咬节长度差别阈值是1.5mm或2mm(如第三次全国大熊猫调查),这对测量的精度要求很高,容易受到主观因素和测量误差的影响[20]。2003年魏荣平等对卧龙自然保护区中的15只不同年龄(2—16岁)大熊猫粪便中的竹子咬节进行了测定,通过竹子咬节平均值的方差分析,发现粪便中竹子咬节长短受个体、日期及它们的交互作用影响,与年龄、性别,山系,野外和饲养环境都没有显著的相关关系,也易受环境(日期,竹子质量等许多不清楚的因素)的影响[21]。因此通过咬节法所得到的数量调查结果存在争议。
第三次全国大熊猫调查对大熊猫种群数量的分析,采用距离区分法和咬节区分法相结合的综合分析方法[11]。调查结果显示,目前野外共有1596只大熊猫(不包括1.5岁以下大熊猫幼体数量)。为了使调查数据更具有可比性,第四次调查基本上沿用了第三次调查采用的距离区分法和咬节区分法对种群数量进行了统计,结果显示截至2013年年底,全国野生大熊猫种群数量达1864只,增加了268只,增长了16.8%。
1.2.3 分子生物学调查法
近些年随着分子生物学、保护遗传学的发展,有专家陆续通过大熊猫DNA进行个体识别,进而统计大熊猫种群数量。其基本原理是:大熊猫粪便在经消化道排出时,会有部分脱落的细胞混杂在粪便表面的黏液中。提取脱落细胞的DNA全基因组,进行个体遗传信息的分析,区分不同大熊猫个体,最终统计种群数量(图2)[22]。高等生物的基因组中含有大量的串联重复序列,主要由两种类型组成,即小卫星DNA和微卫星DNA,对应的采用的分子生物学遗传标记监测技术分别为DNA指纹图谱技术和微卫星标记技术。
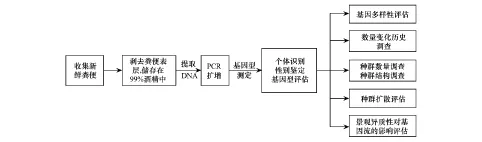
图2 分子生物学在大熊猫调查研究中的流程图及应用[22]Fig.2 Flow chart of genetics method and its application in the Giant Panda research[22]
小卫星DNA是一些重复单位在11—60bp、重复次数在成百以上、总长度由几百至几千个碱基组成的串联重复序列。DNA指纹图谱技术是由DNA指纹探针产生的、多个RFLP图带组成的、具有高度变异性和个体专一性的,并能稳定遗传的限制性片段长度多态性图谱[23]。DNA指纹图谱技术利用多位点探针与基因组中的多个小卫星位点上的等位基因杂交,产生多态性图带,每条可分辨的图带代表一个等位基因位点。因此DNA指纹图谱能够全面地反应基因组的变异性,具有高度的个体特异性。20世纪90年代,方盛国、冯和文等尝试将DNA指纹图谱技术应用于大熊猫数量调查,首次利用分子生物学技术判断野生大熊猫种群数量。2006年余建秋建立了从大熊猫粪便样品中提取高质量DNA的技术体系,并得到清晰易辨的DNA指纹图谱[24]。
DNA指纹图谱技术具有可供分析图带多、并能同时检测多个位点,反映动物的遗传背景的变异程度更精确等优点,而且通过分级离心法可以有效降低粪便样品中污染DNA含量[24]。但该技术存在着工作量大、带型比较误差大、提取DNA数量质量的要求比较严格、成本相对较高、设备要求较多等缺点。
微卫星DNA是指以1—6bp的短核苷酸序列为核心单位、长度小于100bp的小片段,又称为简单重复序列(Simple Sequence Repeats)。微卫星DNA两端的序列一般是相对保守的单拷贝序列, 并且不同等位基因间的重复数存在丰富的差异,通过设计特异引物,对基因组总DNA进行PCR扩增,用以揭示扩增片段的长度多态性(即微卫星标记)[25]。相对于其它分子标记而言,微卫星标记具有分布广泛、选择中性、共显性遗传、多态性高、实验结果稳定可靠等优点。1997年张亚平等对大熊猫微卫星初次进行筛选并应用于亲子鉴定[26]。2006年詹祥江等人通过PCR技术扩增微卫星位点,利用多个位点的微卫星分子标记进行大熊猫的个体鉴定。在对四川王朗自然保护区大熊猫种群数量调查研究中,詹祥江等人得出结论共有66只大熊猫(35只雄性,31只雌性)生存,远高于全国第三次大熊猫调查中的27只[20]。2008年Wu Hua等发现可用于大熊猫非损伤取样样品分析的33个多态性微卫星位点[27]。2010年在《Nature》杂志上公布的大熊猫基因组精细图谱研究成果,为分子生物学鉴定个体提供便利[28]。2015年黄杰等人通过扫描大熊猫全基因组搜索和输出微卫星序列,进一步对微卫星序列进行统计和分析,逐步筛选出粪便样品中同时具备多态性、稳定性、可重复性的四碱基微卫星共15个建立大熊猫微卫星标记系统,同时选择此系统中期望杂合度最高的6个位点建立大熊猫个体识别方法[29]。
微卫星DNA标记克服了DNA指纹技术需求大量高质量DNA的限制[20],但仍有以下问题值得注意。微卫星标记具有种族特异性,通常需针对不同的物种,在了解其基因序列的基础上进行特异性引物设计,此过程包括基因文库构建、基因测序、引物设计与多态性检测等,费时费力[30]。另外采用不同微卫星标记得出的不同结果之间很难横向对比[29]。微卫星侧翼序列变异会带来等位基因大小同源异形和无效等位基因,对群体遗传多样性参数、亲本分析结果等相关分析带来不良影响[29]。同时微卫星标记在处理低浓度、少数量、污染DNA模板时可能会出现等位基因丢失、错印等现象,引起种群数量统计上的偏差。设计恰当基因分型实验方案、减少基因分型过程中的误差、基因分型后的数据清理、数据分析时误差说明等可在很大程度上最终减少误差[31]。现在国际上通常采用哈迪-温伯格检验和孟德尔遗传规律检测无效等位基因;利用重复基因实验来检测等位基因丢失现象[31-32]。
分子生物学调查方法不但可以提供较精确、可信性较强的大熊猫个体鉴定、区别方法,也能提供大熊猫的性别、遗产多样性、种群间基因交流、扩散格局等其他信息,对野生大熊猫保护和栖息地的规划有着至关重要的作用[26]。但其调查成本较高、样品处理要求高等缺点仍没有得到进一步的改善。国内已有许多文献描述如何在粪便中提取DNA[33-34],但提取过程相对于普通保护区/监测站点仍是一个难题。在现有的技术方法、人员配置以及经费支持条件下,监测站只能对粪便进行简单预处理,初步提取DNA,等待送往专业的科研机构进行下一步的工作。这个过程对最终的分析结果是否有影响值得讨论。如何简化DNA提取操作以及如何减少调查过程中的开销是解决分子生物学调查法推广的关键。另外,分子生物学调查法对粪便质量要求较高,如何获得新鲜的大熊猫粪便,也是利用分子生物学手段进行大熊猫种群数量调查需要解决的问题。
2 总结与展望
大熊猫保护的相关政策和管理行动减缓了数量下降,同时狩猎、栖息地破坏等行为几乎消失,野生大熊猫种群和栖息地均得到了有效保护,野生大熊猫的未来光明并充满希望[35]。王朗自然保护区的首次调查揭开了大熊猫种群数量调查的帷幕,经过数十年积累,大熊猫种群数量调查取得巨大进步;调查范围由最初的自然保护区演变为全国范围;调查目的由单一种群数量和分布情况演变为野生大熊猫种群数量、分布现状、栖息地状况、野生大熊猫种群及栖息地受保护状况、野生大熊猫栖息地受干扰或威胁状况、圈养大熊猫种群状况、野生大熊猫分布区经济状况、大熊猫保护管理状况与成效七大方面内容综合调查;调查方法由直接观测过渡到间接识别,由扰动较大到非损伤,由主观判断到客观分析,由单一方法判别个体发展为多种技术手段综合分析鉴定个体;调查工具也逐步采用3S(GPS、GIS、RS)技术等国际最前沿的方法。我国大熊猫种群数量调查体系初步形成。在此,我们根据各种调查方法发展中存在的问题和目前野生大熊猫种群调查发展需求,提出3种未来可能利用的大熊猫种群调查方法。
2.1 分子生物学调查法
经过20余年的沉淀后,分子生物学调查法可以提供较精确、可信性较强的种群数量,新的遗传工具同样揭示了种群结构、量化大熊猫扩散格局,已经成为目前比较值得信任的调查方法之一,预计在今后较长的一段时间内将会作为大熊猫种群数量调查的主要方法。大熊猫基因组测序的发布为基因标记的研发提供便利,在将来使用SNP类型进行标记可能会比微卫星标记更加容易。粪便中提取出成千上万SNP将进一步转变大熊猫的生态学、进化学和保护学的研究[22]。
2.2 红外相机技术
红外相机陷阱技术是指通过自动相机系统来获取野生动物图像数据(如照片和视频),这些图像数据可以用来分析野生动物的物种组成、分布、种群数量、行为和生境利用等基础信息[36]。国外使用红外相机技术开展野生动物调查研究已有较长历史,在20世纪90年代逐步发展成熟;我国使用红外相机技术进行相关研究的工作起步较晚[37],在国内,研究者利用红外相机记录野生动物的活动模式,探测野生动物的多样性,或者记录特定的野生动物,如东北虎、大熊猫、雪豹及华南虎等[38]。相对于传统调查方法,红外相机技术不但具有客观性、非损伤性、监测类群全面等优点,它还可以提供更为丰富的信息(例如物种出现的具体时间、先后顺序、社会结构等)[37]。红外相机装置隐秘和能持续观测等特点适合于观测栖息地地形复杂、采用直接观察难以执行、活动隐秘数量稀少的动物,而且红外相机获取的调查数据可在“标记-重捕”模型(Mark-Recapture Model)的理论框架下,使用专业数据分析软件估算此物种的种群大小和密度[36]。
红外相机技术具有直观监测大熊猫迁移动态、提供准确影像资料、补充丰富监测库数据、观测人为/自然干扰及伴生动物、减轻巡护人员工作压力等优点,在大熊猫行为研究、大熊猫保护中的应用前景光明,但对于大熊猫种群数量调查而言还属于一种相对新的技术[39]。究其原因,主要是大熊猫只有黑白两色,无法根据体侧斑纹模式进行个体识别,仅能通过伤疤或脸部花纹等其它个体特征进行初步识别。另外红外相机技术需要在观测物种主要通道上布设大量相机并定期进行维护,而且后期对数据的分析整理也需要专业人士,这对人力物力是一个极大的考验。现已有部分保护区安装大量红外相机用于监测、保护等目的,尚无涉及种群数量调查,因此,红外相机技术应用于野生大熊猫种群调查需要进一步的探讨与尝试。
2.3 足迹鉴定法
足迹鉴定法通过收集区域内目标动物足迹信息(包括足迹图像、足迹位置、发现时间等),输入基于JMP(SAS insitute)软件上的特定插件,自动提取足迹图像上的几何信息并进行统计分析,结合其他信息综合区分不同个体,最终统计区域内种群数量(图3)[40- 44]。足迹鉴定法的主要分析过程为软件自动完成,不存在主观干扰,国外已利用足迹法对多种动物进行个体识别或种群数量调查,例如老虎(Pantheratigris)和雪豹(Pantherauncia)[41]、黑犀牛(Dicerosbicornis)[42]、美洲狮(Pumaconcolor)[43]等,在我国也有对东北虎(Panthemtigrisaltaica)雪地足迹进行性别鉴定及个体识别的研究[40]。WildTrack动物保护组织与美国杜克大学已合作开展大熊猫足迹识别相关插件的研究工作。
相对于传统调查方法,足迹鉴定法优势不仅在于其客观性、非损伤性,而且它具有很高的投入产出比[45]。足迹鉴定法主要设备为相机与电脑,将符合要求的足迹图片导入程序即可完成鉴定工作,普通监测人员即能胜任对目标物种长时间监测。足迹鉴定法适用于一些夜间出没或是难以直接观察物种的数量调查,并且足迹鉴定法可以更直接监测目标物种的迁移路线,这在一定程度上弥补现有调查方法的不足。同时足迹鉴定法也不会受昂贵仪器或者时间问题限制,因此可以提供比无线遥测技术和红外相机监测技术更真实的动物活动范围[45]。

图3 Sky K.Alibhai等在白犀足迹上做的标记点及衍生点 (a)白犀牛的左后脚(b)白犀牛的左后脚足迹,包含标记点(○)以及衍生点(■)[45]Fig.3 The landmark points and derived points made by Sky K. Alibhai on White rhinofootprint(a) White rhino left hind foot. (b) White rhino lefthind footprint showing landmark points(○)and derived points(■)[45]
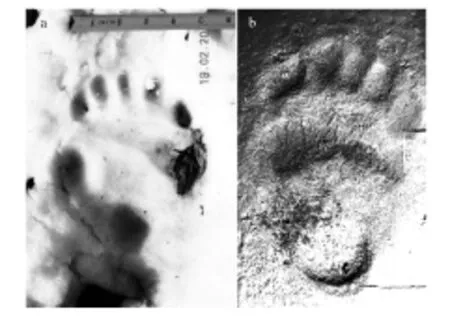
图4 (a)雪作为基质收集的足迹(图片来源:熊猫国际官网)(b)泥土作为基质收集的足迹 Fig.4 (a) Collect footprints in the substrate of snow.(Picture comes from:Panda International)(b) Collect footprints in the substrate of soil
应用足迹法的首要条件是调查区拥有适当的基质,目标种群足底结构足够复杂,这样才会收集到足够清晰、满足分析要求的图片。大熊猫行走习性和脚底特征使得大熊猫足迹可以轻易的同其他物种区分开来(如图4所示,雪地上、泥地上大熊猫的足迹均可以清晰的反映出趾垫的轮廓)[3]。泉眼附近泥塘和冬季有积雪覆盖地面是较为理想的基质。但泥塘分布的局限性和积雪作为基质的不稳定性(容易融化或者被继续的降雪覆盖等)对足迹质量影响很大,因此需要收集大量的足迹图片用于筛选、分析。足迹收集工作可与保护区的日常巡护工作、粪便收集工作同时开展。同时,大熊猫足迹识别相关插件正在研发中,需要进行大量的实验进行校准与验证。
本文通过对几种大熊猫种群数量调查方法进行了较全面的对比分析(表2)。未来开展大熊猫种群数量调查时应综合考虑这些方法的调查范围、时间尺度、人员配置、资金投入等因素,针对不同情形选用不同方法,发挥各种调查方法的优势,保证调查数据的可靠性。另外,对于有条件保护区选用的调查方法应具有延续性,这样历次调查结果可以进行横向对比,反应出的种群数量变化更具有说服力,为有关部门及时调整大熊猫保护策略提供科学依据。
[1] 戈峰. 现代生态学. 北京: 科学出版社, 2008.
[2] 李博, 杨持, 林鹏. 生态学. 北京: 高等教育出版社, 2000.
[3] 胡锦矗. 大熊猫研究. 上海: 上海科技教育出版社, 2001.
[4] 魏辅文, 张泽钧, 胡锦矗. 野生大熊猫生态学研究进展与前瞻. 兽类学报, 2011, 31(4): 412- 421.
[5] 国家林业局. 中国大熊猫现状展——全国第四次大熊猫调查简况. (2015-03-03)[2016-03- 21]. http://202.99.63.178/main/69/content- 743562.html.
[6] 冯文和, 方盛国, 张安居, 赵佳, 藤原昇. 大熊猫种群数量调查方法的新突破. 大自然探索, 1998, 17(2): 44- 48.
[7] 王朗自然保护区大熊猫调查组. 四川省平武县王朗自然保护区大熊猫的初步调查. 动物学报, 1974, 20(2): 162- 173.
[8] 马建章, 罗泽珣. 野生动物数量的调查. 野生动物, 1983, (6): 24- 28.
[9] 潘文石, 高郑生, 吕植. 秦岭大熊猫的自然庇护所. 北京: 北京大学出版社, 1988.
[10] 潘文石, 吕植, 朱小健, 王大军, 王昊, 龙玉, 付达莉, 周欣. 继续生存的机会. 北京: 北京大学出版社, 2001.
[11] 国家林业局. 全国第三次大熊猫调查报告. 北京: 科学出版社, 2006.
[12] 四川省珍贵动物资源调查队. 四川省珍贵动物资源调查报告. 成都: 四川省林业局, 1977.
[13] 胡锦矗, 夏勒. 卧龙的大熊猫. 成都: 四川科学技术出版社, 1985.
[14] 胡锦矗, 胡杰. 大熊猫研究与进展. 四川师范学院学报: 自然科学版, 2003, 24(3): 253- 257.
[15] Zhang Z J, Sheppard J K, Swaisgood R R, Wang G, Nie Y G, Wei W, Zhao N X, Wei F W. Ecological scale and seasonal heterogeneity in the spatial behaviors of giant pandas. Integrative Zoology, 2014, 9(1): 46- 60.
[16] Zhang J D, Hull V, Huang J Y, Zhou S Q, Xu W H, Yang H B, McConnell W J, Li R G, Liu D, Huang Y, Ouyang Z Y, Zhang H M, Liu J G. Activity patterns of the giant panda (Ailuropodamelanoleuca). Journal of Mammalogy, 2015, 96(6): 1116- 1127.
[17] Hull V, Zhang J D, Zhou S Q, Huang J Y, Li R G, Liu D, Xu W H, Huang Y, Ouyang Z Y, Zhang H M, Liu J G. Space use by endangered giant pandas. Journal of Mammalogy, 2015, 96(1): 230- 236.
[18] 尹玉峰, 王昊, 陈艾, 刘国琪. 对大熊猫数量调查方法中咬节区分机制的准确性评价. 生物多样性, 2005, 13(5): 439- 444.
[19] 张泽钧, 杨旭熤, 吴华, 李明, 魏辅文. 大熊猫粪便宽径与咬节平均长度的关系. 兽类学报, 2005, 25(4): 351- 354.
[20] Zhan X J, Li M, Zhang Z J, Goossens B, Chen Y P, Wang H J, Bruford M W, Wei F W. Molecular censusing doubles giant panda population estimate in a key nature reserve. Current Biology, 2006, 16(12): R451-R452.
[21] 魏荣平, 张贵权, 王鹏彦, 李德生, 曹家林. 大熊猫粪便中竹子咬节长短与年龄和种群数量关系. 生态学报, 2003, 23(10): 2153- 2162.
[22] Wei F W, Hu Y B, Zhu L F, Bruford M W, Zhan X J, Zhang L. Black and white and read all over: the past, present and future of giant panda genetics. Molecular Ecology, 2012, 21(23): 5660- 5674.
[23] 陈振文. DNA指纹图与微卫星DNA技术在近交系大、小鼠遗传监测中的应用研究[D]. 北京: 中国农业大学, 2004.
[24] 余建秋. 大熊猫基因资源库构建及粪便高质量DNA提取方法的建立[D]. 杭州: 浙江大学, 2006.
[25] 孙波, 鲍毅新, 赵庆洋, 张龙龙, 胡知渊. 微卫星位点获取方法的研究进展. 生态学杂志, 2009, 28(10): 2130- 2137.
[26] 张亚平, 王文, 宿兵, Ryder O A, 范志勇, 张和民, 何廷美. 大熊猫微卫星DNA的筛选及其应用. 动物学研究, 1995, 16(4): 301- 306.
[27] Wu H, Zhan X J, Zhang Z J, Zhu L F, Yan L, Li M, Wei F W. Thirty-three microsatellite loci for noninvasive genetic studies of the giant panda (Ailuropodamelanoleuca). Conservation Genetics, 2009, 10(3): 649- 652.
[28] Li R Q, Fan W, Tian G, Zhu H M, He L, Cai J, Huang Q F, Cai Q L, Li B, Bai Y Q, Zhang Z H, Zhang Y P, Wang W, Li J, Wei F W, Li H, Jian M, Li J W, Zhang Z L, Nielsen R, Li D W, Gu W J, Yang Z T, Xuan Z L, Ryder O A, Leung F C C, Zhou Y, Cao J J, Sun X, Fu Y G, Fang X D, Guo X S, Wang B, Hou R, Shen F J, Mu B, Ni P X, Lin R M, Qian W B, Wang G D, Yu C, Nie W H, Wang J H, Wu Z G, Liang H Q, Min J M, Wu Q, Cheng S F, Ruan J, Wang M W, Shi Z B, Wen M, Liu B H, Ren X L, Zheng H S, Dong D, Cook K, Shan G, Zhang H, Kosiol C, Xie X Y, Lu Z H, Zheng H C, Li Y R, Steiner C C, Lam T T Y, Lin S Y, Zhang Q H, Li G Q, Tian J, Gong T M, Liu H D, Zhang D J, Fang L, Ye C, Zhang J B, Hu W B, Xu A L, Ren Y Y, Zhang G J, Bruford M W, Li Q B, Ma L J, Guo Y R, An N, Hu Y J, Zheng Y, Shi Y Y, Li Z Q, Liu Q, Chen Y L, Zhao J, Qu N, Zhao S C, Tian F, Wang X L, Wang H Y, Xu L Z, Liu X, Vinar T, Wang Y J, Lam T W, Yiu S M, Liu S P, Zhang H M, Li D S, Huang Y, Wang X, Yang G H, Jiang Z, Wang J Y, Qin N, Li L, Li J X, Bolund L, Kristiansen K, Wong G K S, Olson M, Zhang X Q, Li S G, Yang H M, Wang J, Wang J. The sequence anddenovoassembly of the giant panda genome. Nature, 2010, 463(7279): 311- 317.
[29] Huang J, Li Y Z, Du L M, Yang B, Shen F J, Zhang H M, Zhang Z H, Zhang X Y, Yue B S. Genome-wide survey and analysis of microsatellites in giant panda (Ailuropodamelanoleuca), with a focus on the applications of a novel microsatellite marker system. BMC Genomics, 2015, 16: 61.
[30] 文亚峰, Uchiyama K, 韩文军, Ueno S, 谢伟东, 徐刚标, Tsumura Y. 微卫星标记中的无效等位基因. 生物多样性, 2013, 21(1): 117- 126.
[31] Pompanon F, Bonin A, Bellemain E, Taberlet P. Genotyping errors: causes, consequences and solutions. Nature Reviews Genetics, 2005, 6(11): 847- 859.
[32] Guichoux E, Lagache L, Wagner S, Chaumeil P, Léger P, Lepais O, Lepoittevin C, Malausa T, Revardel E, Salin F, Petit R J. Current trends in microsatellite genotyping. Molecular Ecology Resources, 2011, 11(4): 591- 611.
[33] 魏辅文, 饶刚, 李明, 方盛国, 冯祚建. 分子粪便学及其应用——可靠性、局限性和展望. 兽类学报, 2001, 21(2): 143- 152.
[34] 钟华, 赖旭龙, 魏荣平, 刘中来. 一种从大熊猫粪便中提取DNA的改进方法. 动物学报, 2003, 49(5): 670- 674.
[35] Wei F W, Swaisgood R, Hu Y B, Nie Y G, Yan L, Zhang Z J, Qi D W, Zhu L F. Progress in the ecology and conservation of giant pandas. Conservation Biology, 2015, 29(6): 1497- 1507.
[36] 肖治术, 李欣海, 王学志, 周岐海, 权锐昌, 申小莉, 李晟. 探讨我国森林野生动物红外相机监测规范. 生物多样性, 2014, 22(6): 704- 711.
[37] 李晟, 王大军, 肖治术, 李欣海, 王天明, 冯利民, 王云. 红外相机技术在我国野生动物研究与保护中的应用与前景. 生物多样性, 2014, 22(6): 685- 695.
[38] 刘芳, 李迪强, 吴记贵. 利用红外相机调查北京松山国家级自然保护区的野生动物物种. 生态学报, 2012, 32(3): 730- 739.
[39] 卢学理, 蒋志刚, 唐继荣, 王学杰, 向定乾, 张建平. 自动感应照相系统在大熊猫以及同域分布的野生动物研究中的应用. 动物学报, 2005, 51(3): 495- 500.
[40] 顾佳音. 东北虎雪地足迹个体识别技术研究[D]. 哈尔滨: 东北林业大学, 2013.
[41] Riordan P. Unsupervised recognition of individual tigers and snow leopards from their footprints. Animal Conservation, 1998, 1(4): 253- 262.
[42] Jewell Z C, Alibhai S K, Law P R. Censusing and monitoring black rhino (Dicerosbicornis) using an objective spoor (footprint) identification technique. Journal of Zoology, 2001, 254(1): 1- 6.
[43] Lewison R, Fitzhugh E L, Galentine S P. Validation of a rigorous track classification technique: identifying individual mountain lions. Biological Conservation, 2001, 99(3): 313- 321.
[44] Funston P J, Frank L, Stephens T, Davidson Z, Loveridge A, Macdonald D M, Durant S, Packer C, Mosser A, Ferreira S M. Substrate and species constraints on the use of track incidences to estimate African large carnivore abundance. Journal of Zoology, 2010, 281(1): 56- 65.
[45] Alibhai S K, Jewell Z C, Law P R. A footprint technique to identify white rhinoCeratotheriumsimumat individual and species levels. Endangered Species Research, 2008, 4: 205- 218.
Research progress on population investigation methods for wild giant panda
SHI Xuewei1,3, ZHANG Jindong2, OUYANG Zhiyun1,*
1StateKeyLaboratoryofUrbanandRegionalEcology,ResearchCenterforEco-environmentalScience,ChineseAcademyofSciences,Beijing100085,China2CollegeofLifeSciences,ChinaWestNormalUniversity,Nanchong637009,China3UniversityofChineseAcademyofSciences,Beijing100049,China
The giant panda (Ailuropodamelanoleuca) is endemic to China and is a flagship for wildlife protection. Changes in panda population numbers reflect population dynamics in real-time, and also provide direct information about panda distribution and habitat. Panda population numbers are a basic requirement for giant panda resource surveys, and ensure the effective implementation of conservation measures. This paper reviews the progress of panda population survey methods, including the Direct Counting Method, the Mathematical Model, Distance-Bamboo Stem Fragments Method, and the Molecular Biology Method. In addition, we discuss new methods, such as camera-trapping and the Footprint Identification Technique, used to investigate populations of wild pandas in recent years. This paper reviews possible problems with traditional methods, and examines the prospects for the application of new methods in the future. Suggestions and prospects are proposed for future research into wild populations of giant pandas.
the giant panda; population investigation method; distance-bamboo stem fragments method; molecular biology method; infrared camera-trapping; footprint identification technique
国家林业局大熊猫国际资金项目(CM1417)
2015- 10- 29;
2016- 04- 25
10.5846/stxb201510292185
*通讯作者Corresponding author.E-mail: zyouyang@rcees.ac.cn
史雪威,张晋东,欧阳志云.野生大熊猫种群数量调查方法研究进展.生态学报,2016,36(23):7528- 7537.Shi X W, Zhang J D, Ouyang Z Y.Research progress on population investigation methods for wild giant panda.Acta Ecologica Sinica,2016,36(23):7528- 7537.
