低温胁迫对两种樟树抗寒性生理的影响
2016-04-08袁美丽河南科技大学林学院河南洛阳47000洛阳市隋唐城遗址植物园河南洛阳4700
王 宁,袁美丽,丁 博.河南科技大学林学院,河南洛阳47000.洛阳市隋唐城遗址植物园,河南洛阳4700
低温胁迫对两种樟树抗寒性生理的影响
王宁1,袁美丽2,丁博2
1.河南科技大学林学院,河南洛阳471000
2.洛阳市隋唐城遗址植物园,河南洛阳471002
摘要:以猴樟和芳樟叶片为试材,通过低温处理研究了两种树木叶片MDA、脯氨酸、可溶性蛋白质及SOD活性和电导率的变化。结果表明:随着处理温度的降低,两种樟树的电解质外渗率呈“S”型上升;两种樟树的SOD活性及脯氨酸含量呈现先升后降的变化;可溶性糖、可溶性蛋白质及MDA含量则均呈现先降后升再降的变化;采用Fuzzy数学隶属函数综合评判法对猴樟和芳樟的抗寒性进行综合评判猴樟的抗寒性大于芳樟,这与利用电解质外渗率结合Logistic方程得出猴樟和芳樟的半致死温度(LT50)分别为-12.43、-10.71℃结果相一致;以猴樟为例,对低温胁迫过程中各抗寒指标隶属函数值进行的综合评判结果显示其抗寒性分抗寒性减弱期、增强期及减弱期。
关键词:樟树;抗寒性;SOD活性;综合评价
低温是限制植物北引的主要环境胁迫因子[1]。随着全球气候的逐渐变暖,南方树种有望逐渐向北方地区引种。我国常绿阔叶树种资源较为丰富,但主要分布于亚热带温暖湿润地区。目前,常绿阔叶树种以其四季常青、革质光亮及花色果色丰富等特点,已被广泛应用于我国南方城市绿化。对于常绿阔叶树种相对匮乏的北方城市而言,樟树作为优良的园林绿化树种越来越受到北方城市绿化部门的重视。关于樟树的抗寒性研究相对较少,薛建辉等[2]结合自然降温过程中叶片相对电导率和低温半致死温度(LT50)测定,对南京地区引种的浙江樟等5种常绿阔叶树种进行了抗寒性研究,将5种树种的抗寒适应性分为抗寒性较弱期、抗寒性增强期、抗寒性最强期和抗寒性减弱期4个阶段,并得出浙江樟抗寒能力较强,且能在南京地区广泛栽植的结论。谢晓金等[3]借助电导法,对南京地区露地引种的包括香樟、浙江樟等12种常绿阔叶树种进行了抗寒性研究,研究结果表明1月中旬抗寒能力达到最强,而通常10月份或3月份期间抗寒能力最弱。而对冬季低温条件相应的快慢程度的研究表明,香樟、浙江樟属于低温快速响应型。尤扬等[4]研究认为,当温度低于-10℃时香樟容易发生冻害。毛春英[5,6]在山东泰安对香樟幼苗及种子分别进行了引种研究,结果表明引种种子进行播种繁殖更易成功,并能获得抗寒性强的苗木。本研究以实生繁殖的猴樟(Cinnamonum bodinieri Levl)及芳樟(C. camphora L.(linalool-type))为试材,通过对不同低温处理中其叶片膜透性、保护酶活性及渗透调节物含量动态变化的研究,以期更加全面科学的鉴定两种樟树的抗寒性,揭示植物在不同低温胁迫的适应机制。
1 材料与方法
1.1实验材料
本试验采用的材料为实生繁殖的1 a生猴樟及芳樟幼苗,种子均来源于湖南省长沙县跳马镇(东经113°12′~113°13′,北纬28°22′~28°23′)。试验地位于河南省郑州市河南农业大学第三生活区内。郑州市位于东经112°42′~114°14′,北纬34°16′~34°58′,属北温带季风型大陆性气候,年均气温为14.2℃,一月份均温0.2°C,极端低温-19.7℃。全年平均无霜期为227 d,初霜期在每年11月。
1.2实验设计
2011年12月,选取两种樟树的1 a健壮苗木各10株,并在东南方向生长健康的枝条,取枝条上倒2和倒3叶片,先用自来水冲洗干净,再用去离子水漂洗3次,在滤纸上吸干。剪碎后分成7份,每份约3 g左右,用自封袋包装置于低温冰箱中。实验设置6个低温梯度,分别为0、-5、-10、-15、-20、-25℃,处理均为24 h,取样进行各项生理指标的测定,以采样当天(2011年12月5日,0/4℃)的叶片为对照(CK)。每个处理叶片3份,3次重复。
1.3测定项目及方法
参考李合生[7]方法,蒽酮比色法测定叶片内可溶性糖,考马斯亮蓝法测定可溶性蛋白质含量,酸性茚三酮测定脯氨酸含量,丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)法,超氧化物歧化酶(SOD)活性的测定采用NBT光照化学还原法,参照邹琦[8]电导法进行电解质渗透率的测定。
1.4数据处理
应用Fuzzy数学中隶属函数法进行综合评判[9,10],其计算公式如下:(1)对与抗寒性正相关的可溶性糖、可溶性蛋白质和脯氨酸,采用公式:f(xij)=(Xij-Xj min)/(Xj max-Xj min);(2)对与抗寒性呈负相关的电导率及MDA含量,采用下列公式:f(xij)=1-(Xij-Xj min)/(Xj max-Xj min)。式中,f(xij)为i树种的j项指标隶属函数值;Xij为其测定值;Xj max和Xj min分别为j指标最大测定值和最小测定值。
试验数据用Excel及SPSS统计软件进行相关分析和方差分析。
2 结果分析
2.1电导率的变化
低温胁迫下,两种樟树叶片的电导率均随胁迫温度的降低而增大(图1)。其中,以芳樟的上升幅度较大。一般认为,相对电导率达到50%时的温度可作为植物的LT50。由图中可以看出,猴樟及芳樟在-10℃时的电导率分别为48.69%、52.99%,均较接近50%。由此表明,两种樟树的半致死温度均在-10℃左右。
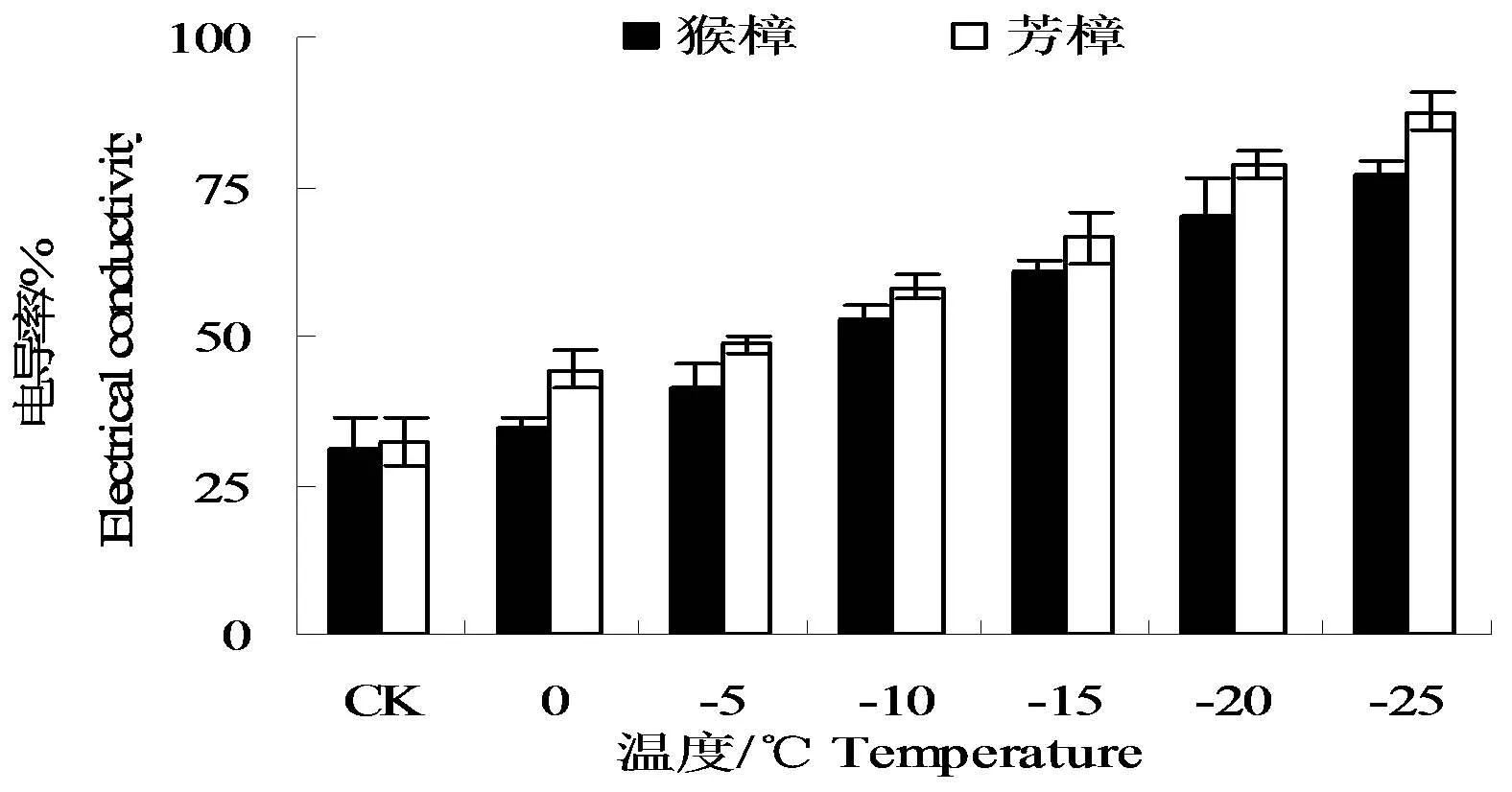
图1 低温处理后两种樟树的电导率变化Fig.1 The changes of electrical conductivity in leaves of two camphor species under low temperature
为进一步准确判断两种树木的半致死温度,将各处理温度下两种樟树叶片的电导率用Logistic方程y=K/(1+αe-bx)进行拟合,求出猴樟和芳樟的LT50分别为-12.43、-10.71℃(表1)。从表中可以看出,猴樟及芳樟的拟合度(R2)均达到0.99以上。综上所述,两种樟树细胞膜低温稳定性以猴樟相对较强。
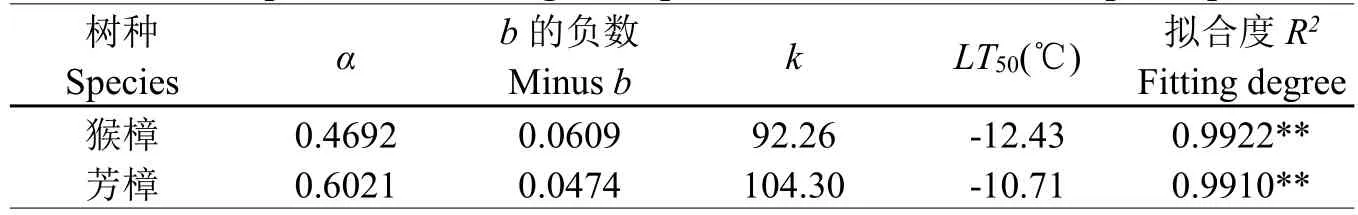
表1 两种樟树电导率的Logistic方程参数及LT50Table 1 The parameters of Logistic equation and LT50of two camphor species
2.2MDA含量
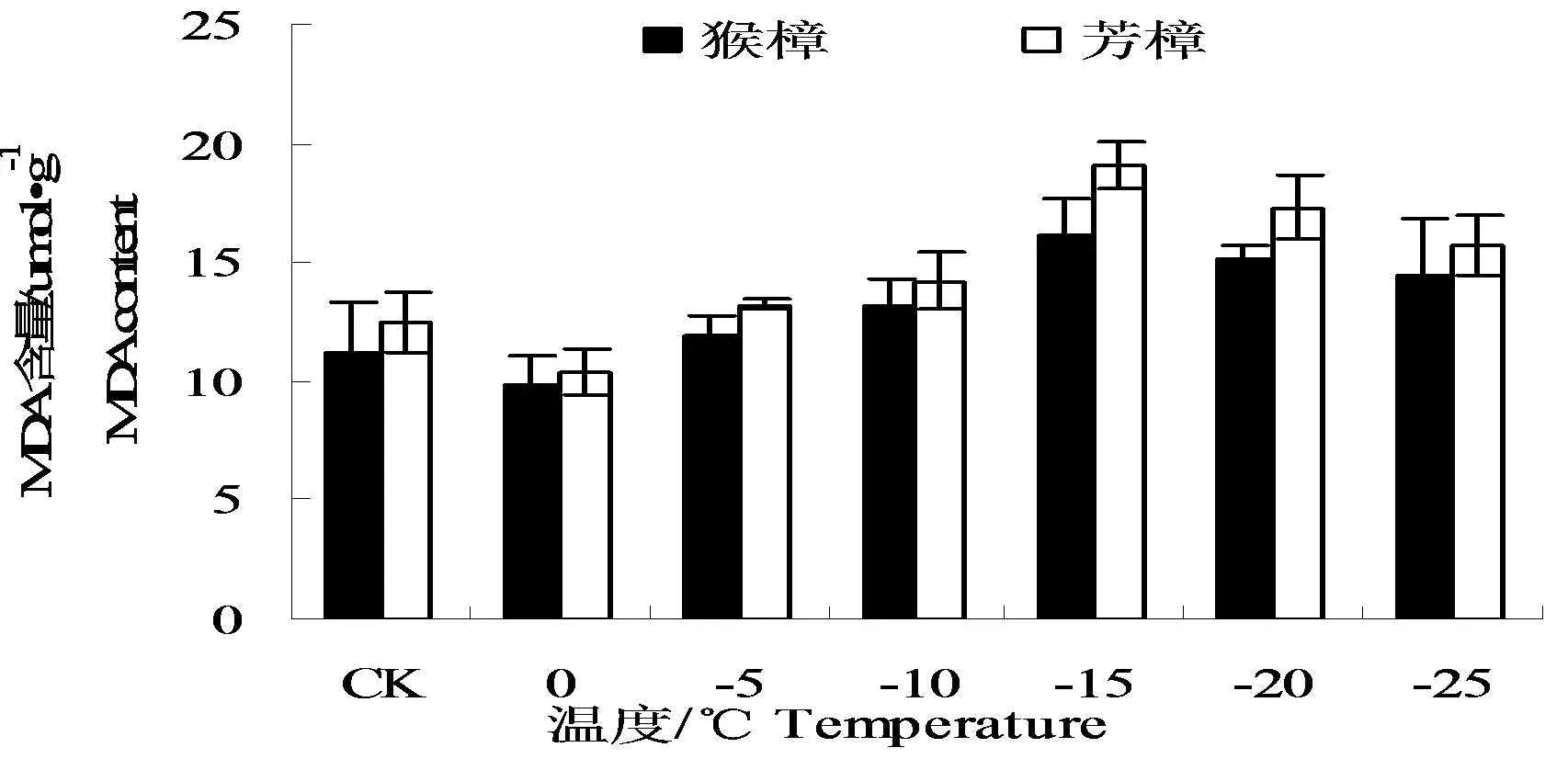
图2 低温处理后两种樟树的MDA含量的变化Fig.2 The changes of MDAcontent in leaves of two camphor species under low temperature
由图2中可以看出,随着温度的降低,两种樟树叶片MDA含量呈先降后升又降的变化。MDA含量前期的增加可能与樟树前期保护酶活性及渗透调节物质的增加相关,而随后的上升是低温胁迫的进一步增大的结果,最后的下降可能与低温超过其承受范围,造成植物组织逐渐死亡相关。图中显示,芳樟的MDA含量在各低温下均高于猴樟,说明芳樟的膜脂过氧化程度均高于猴樟。两种樟树MDA含量均在0℃时最低,与最低值相比,猴樟和芳樟在-15℃时MDA含量分别为16.20、19.11 μmol·g-1,分别增加了64.47、84.46%。
2.3脯氨酸含量的变化
游离脯氨酸作为重要的渗透调节物质,低温胁迫下其含量的增加,有利于稳定蛋白质结构和保护细胞内大生物分子的作用。从图3中可以看出,低温胁迫初期两种樟树的脯氨酸含量均出现不断的增加,且以猴樟增加较为明显,至-15℃时脯氨酸含量达到最大,分别为0.3089、0.2548 mg·g-1,分别比对照增加了24.14%和10.49%。当低温超过-15℃时,脯氨酸含量有所下降,这可能与樟树在重度胁迫下的耐受性有限相关。
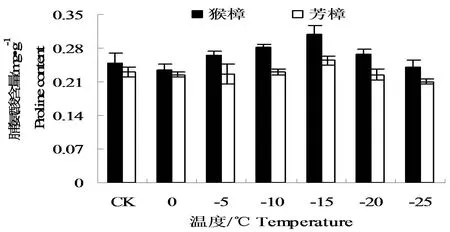
图3 低温处理后两种樟树的脯氨酸含量的变化Fig.3 The changes of proline content in leaves of two camphor species under low temperature
2.4可溶性糖、可溶性蛋白含量的变化
从图中4可以看出,芳樟和猴樟叶片内可溶性糖和可溶性蛋白含量均呈先降后升再降的变化,前期的下降可能是两种樟树对低温的应激反应,随后可溶性糖和可溶性蛋白含量的增加,至-15℃时出现峰值,且均以猴樟的含量相对较高,分别比芳樟高16.42%及17.74%,表明两种樟树在适度的低温胁迫中,通过可溶性糖和可溶性蛋白的含量提高来增强其抗冻能力;然而,随着温度的进一步降低,两种渗透物质的含量均出现明显的下降,表明其调节能力的下降,这可能是低温胁迫超过了两种樟树的承受范围。
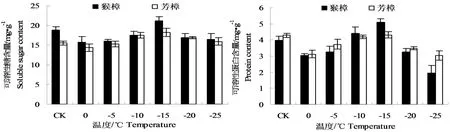
图4 低温处理后两种樟树的可溶性糖、可溶性蛋白含量的变化Fig.4 The changes of soluble sugar and protein content in leaves of two camphor species under low temperature
2.5SOD活性的变化
SOD作为植物体内清除O2-重要保护酶类之一,其活性的大小反映了植物对逆境胁迫的适应能力。低温胁迫对两种樟树SOD活性的影响见图5。从图中可以看出,两种樟树的SOD活性均呈现先升后降的变化,且峰值均出现-5℃,与对照相比,分别增加了61.86、85.65%,而猴樟的峰值则比芳樟高出了13.27%。随着胁迫温度的持续降低,SOD活性出现大幅下降,这可能与胁迫低温超过了其承受的范围,而导致保护酶活性失活相关。
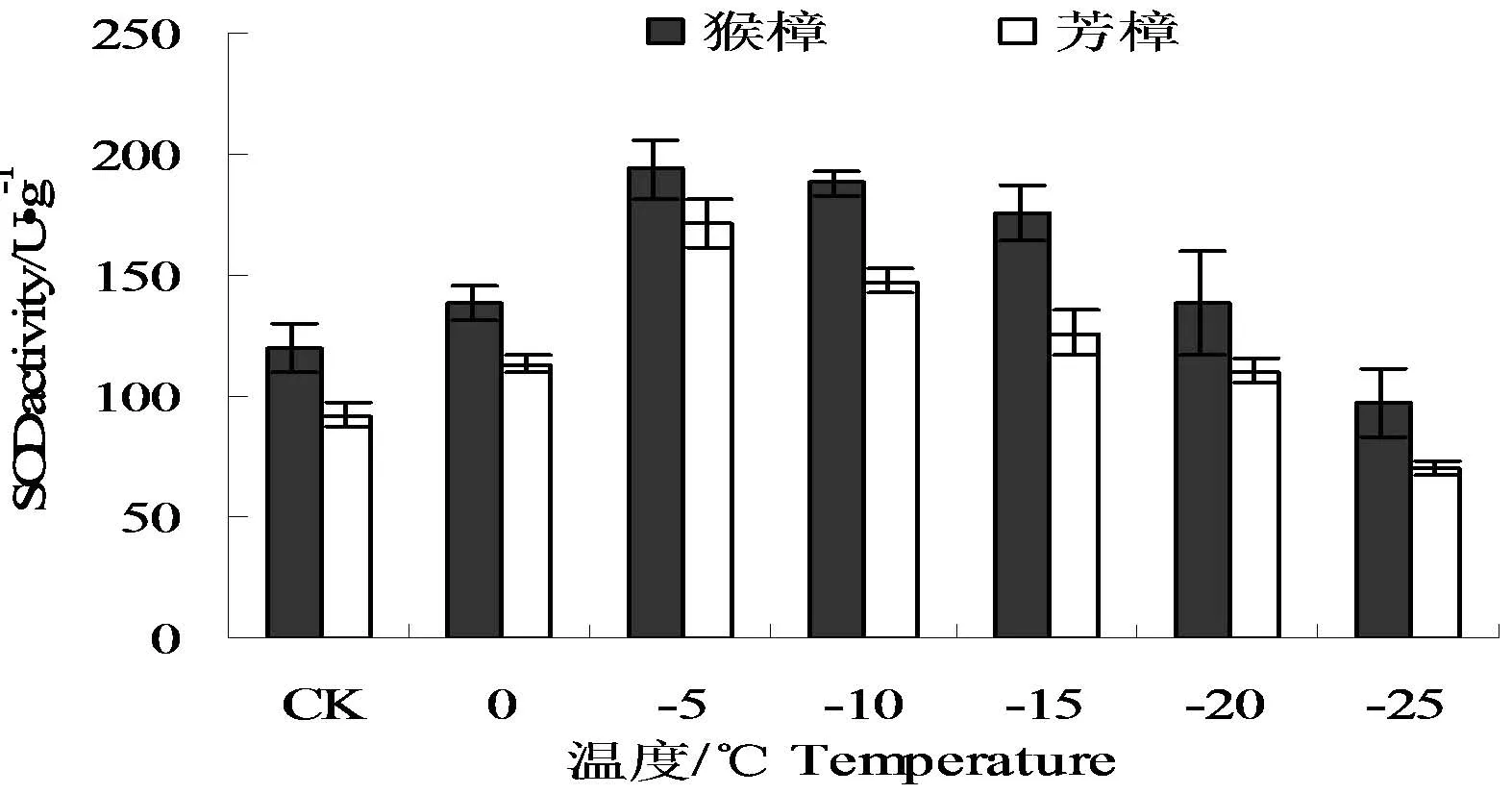
图5 低温处理后两种樟树的SOD活性的变化Fig.5 The changes of SOD activity in leaves of two camphor species under low temperature
2.6隶属函数法综合分析抗寒性
2.6.1低温胁迫过程中各抗寒指标隶属值以猴樟为例,低温胁迫过程中各抗寒指标隶属函数值见表2。综合评价值呈现出“降-升-降”的变化趋势,表明低温胁迫过程中,猴樟的抗寒性呈减弱、增强及减弱的变化。由此可将低温胁迫过程中猴樟的抗寒性分为2个时期,分别为:(1)抗寒性减弱期。此期包括两个阶段,从CK到0℃,猴樟抗寒性的减弱,可能与其应激反应有关;至-15℃后,其抗寒性的大幅下降,可能与胁迫低温超过了其承受范围,导致了植物细胞的不可逆性伤害,从而造成其抗寒性的减弱;(2)抗寒性增强期(0℃至-10℃)。猴樟抗寒性呈逐渐增强的变化,表明在对低温短暂的适应后,其通过渗透调节物质含量的积累及保护酶活性的增加,从而增强对低温的适应能力。
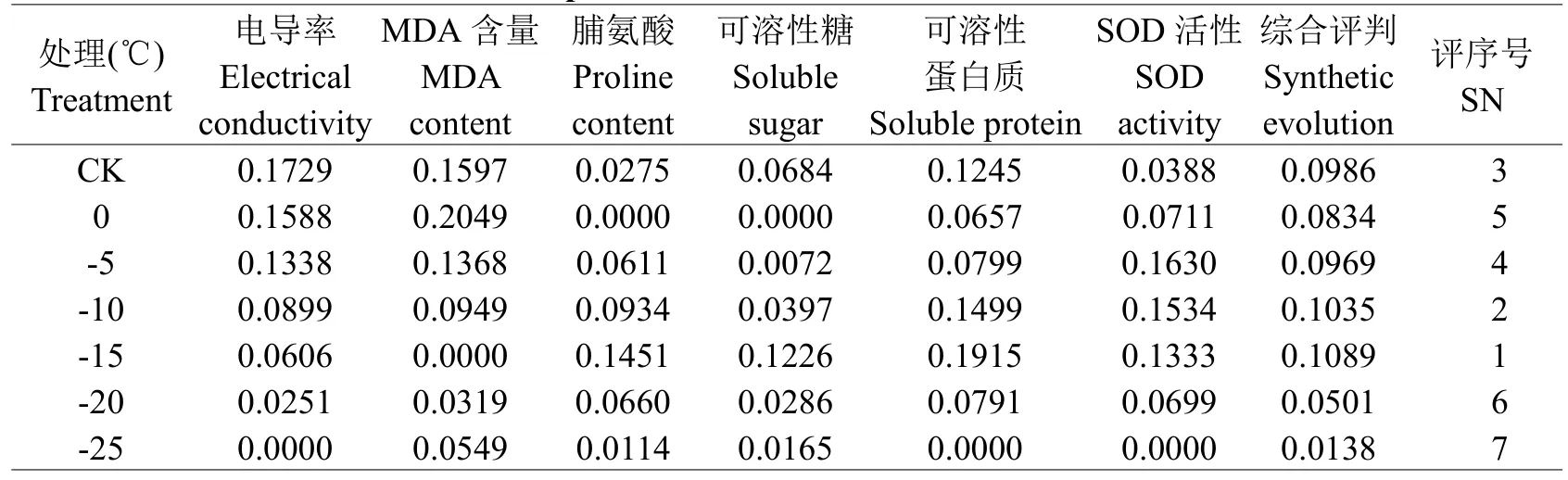
表2 猴樟低温胁迫过程中各抗寒指标隶属值Table 2 The membership values of cold resistance indicators under cold stress
2.6.2抗寒性综合评价植物抗寒性时多指标综合作用的结果,运用上述公式(1)、(2)求出猴樟及芳樟各指标参数的隶属函数值,再乘以各指标权重后求其平均值得其综合评价值,该值越大,则抗寒性即越强。2种樟树6项抗寒指标参数的综合评价结果如表3所示,得出抗寒性强弱顺序为:猴樟>芳樟。
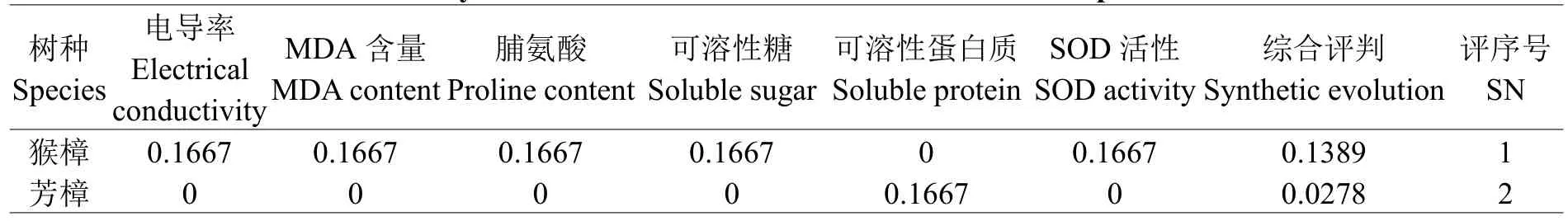
表3 两种樟树抗寒性综合评判Table3 Synthetic evaluation of cold resistance of two camphor trees
3 结论与讨论
3.1不同低温对两种樟树的抗寒性影响
3.1.1低温对细胞膜透性的影响细胞膜是植物细胞与环境发生物质交换、信息传递以及能量交换的重要场所和媒介,也是细胞感受外界环境胁迫最为敏感的部位[11],是低温伤害的原初部位[12,13]。许多研究已经证明,不同梯度低温胁迫中,随着低温的下降,植物电导率持续的上升[14-16]。本试验结果表明,随着处理温度的下降,两种樟树的电导率呈“S”型曲线上升,这与前人研究结果一致。自Rajashekar C等[17]利用Logistic曲线描述低温对植物细胞膜的伤害过程,提出以曲线拐点为半致死温度(LT50)的观点,此法已广泛的应用于不同植物的抗寒性研究[18-20]。本研究中,两种樟树的回归方程相关系数R2分别为0.9922、0.9910,效果较好。研究认为,拐点温度上表现出的差异可以衡量其忍受低温的能力,拐点温度越低,表明其半致死温度越低,则该树种的抗寒性就越强[21]。本研究中,猴樟的半致死温度为-12.43℃,而芳樟的为-10.71℃,由此判断猴樟的抗寒性强于芳樟。
3.1.2低温对渗透调解物质的影响研究表明,脯氨酸、可溶性糖及蛋白参与调控抗寒能力的形成[22-24],可溶性糖可以提高细胞的渗透浓度,降低水势,增加保水能力,从而使冰点下降,提高植物的抗寒性,而脯氨酸作为细胞质的渗透调节物质,在植物对抗低温胁迫时起到平衡细胞代谢的作用,以保持细胞内环境的相对稳定,其含量的增加有利于植物抗寒性的提高[25]。
本研究表明,不同低温胁迫中,两种樟树的可溶性糖、可溶性蛋白及脯氨酸含量均呈先降后升又降的变化,前期的下降可能是对低温应激反应的结果,这与张纪林等[26]对枸骨等冬青植物的研究结果相一致,随后的上升,表明两种通过渗透调节物质含量的增加来提高其抗冻能力,随着胁迫温度的进一步降低,渗透调节物质含量出现下降趋势,这可能与低温超过了两种樟树的承受范围,其调节能力降低相关,这与冯献宾等[16]对黄连木和黄山栾树的研究结果一致。
3.1.3低温对膜脂过氧化的影响MDA作为膜脂过氧化的主要产物质之一,其含量的高低可以反映植物遭受逆境伤害的程度[27]。本研究中,不同低温胁迫中两种樟树的MDA含量前期先降后升的变化与周建等对广玉兰[28]、郑国华等对枇杷[29]的研究结果一致,而最后的下降,表明胁迫低温超过了两种樟树的承受范围,造成组织逐渐死亡,导致细胞膜脂过氧化程度的降低,这与高京草等对枣树[30]、胡春霞等对南国梨[31]、郑元等对几种仁用杏[32]的抗寒性研究结果一致。
3.1.3低温对保护酶活性的影响SOD是植物细胞内最重要的清除活性氧的酶之一,对膜系统的稳定性有着重要的作用,其活性的高低直接反映了活性氧的清除能力大小,与植物的抗逆性关系密切[33,34]。在强度的逆境胁迫中,酶活性一般随胁迫的增加而增加,或者呈先增加后降低的变化[35]。本研究表明,不同低温的胁迫中,SOD活性同样呈先升后降的变化,后期的下降则可能是胁迫低温达到临界温度,导致SOD活性出现钝化的结果,造成其活性的下降,这与石雪辉等对柑桔[36]、詹福建等对马占相思树[37]、段肖翠等对阿月浑子[38]的研究结果一致。
3.1.4抗寒性综合评价植物的抗寒性是受其生理生化特征综合作用的遗传表现,因此,单一抗寒指标难以判断植物对低温的综合适应能力[9]。韩宏伟等[39]、柳新红等[40]运用Fuzzy数学隶属函数综合评判法对中国刺槐种源间和翅荚木种源的抗寒性进行了综合评判,并得出了满意的结果。本研究运用同样的方法,对不同低温胁迫中MDA、SOD、可溶性糖、可溶性蛋白及脯氨酸5个生理生化参数进行了综合分析,得出猴樟抗寒性大于芳樟的结果。
以猴樟为例,对不同低温胁迫过程中各抗寒指标隶属函数值进行的综合评判结果显示,低温胁迫过程中,其抗寒性又分抗寒性减弱期、增强期及减弱期,前期抗寒性的减弱可能是对低温胁迫的应激反应结果,随着温度的持续降低,通过渗透调节物质含量及保护酶活性的增加提高其对低温的适应能力,抗寒性也逐渐增强,随后抗寒性的降低可是胁迫低温超过了其承受的范围,造成植物组织不可恢复的损伤所至。
参考文献
[1] Stushnoff C,Junttila O. Seasonal Development of Cold Stress Resistance in Several Plant Species at a Coastal and a Continental Location in North Norway [J]. Polar Biology,1986,5(3):129-133
[2]薛建辉,苏敬,刘金根,等.5个常绿阔叶园林树种对低温变化的生理响应[J].南京林业大学学报:自然科学版,2009,33(4):38-42
[3]谢晓金,郝日明.南京地区12种常绿阔叶树种冬季抗寒性动态变化[J].生态学报,2009,29(4):2149-2154
[4]尤扬,刘弘,吴荣升,等.低温胁迫对香樟幼树抗寒性的影响[J].广东农业科学,2008,(11):23-25
[5]毛春英.香樟的引种与驯化研究[J].山东农业大学学报:自然科学版,2004,35(4):534-539
[6]毛春英,张纪德,王秀梅.樟树引种驯化及抗寒育苗栽培技术[J].林业科技,2001,26(6):10-12
[7]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000
[8]邹琦.植物生理生化实验指导[M].北京:中国农业出版社,2000
[9]李轶冰,杨顺强,任广鑫,等.低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J].生态学报,2009,29(3):1341-1347
[10]田钦谟.模糊综合评价中的若干问题[J].模糊系统与数学,1996,10(2):62-69
[11]王洪春.植物抗逆性与生物膜结构功能研究进展[J].植物生理学通讯,1985,1:60-66
[12]何若韫.植物低温逆境生理[M].北京:中国农业出版社,1995
[13]简令成.生物膜与植物寒害和抗寒性的关系[J].植物学通报,1983,1(1):17-23
[14]徐康,夏宜平,徐碧玉,等.以电导法配合Logistic方程确定茶梅‘小玫瑰’的抗寒性[J].园艺学报,2005,32(1):148-150
[15]王永格,丛日晨.常绿阔叶树种小果卫矛引种北京的抗寒性比较研究[J].北京农学院学报,2007,22(4):13-16.
[16]冯献宾,董倩,李旭新,等.黄连木和黄山栾树的抗寒性[J].应用生态学报,2011,22(5):1141-1145
[17] Rajashekar C,Gusta LV,Burke MJ. Membrane structural transition: probable relation to frostdamage in hardyherbaceous species [M]//Lyons J M,Graham D,Raison J K. Low temperature stress in crop plants the role of membrane. New York: Academin Press,1979:255-274
[18]缴丽莉,路丙社,白志英,等.四种园林树木抗寒性的比较分析[J].园艺学报,2006,33(3):667-670
[19]李刚,姜卫兵,翁忙玲,等.木兰科6种常绿树幼苗抗寒性的初步研究[J].园艺学报,2007,34(3):783-786
[20]侯渝嘉,唐敏,胡翔.茶树种质资源的抗寒性鉴定[J].西南农业学报,2010,23(1):137-140
[21]贾遂民,陈辉,陈玉珍,等.以电导法配合Logistic方程确定香椿种源间抗寒能力[J].河南科学,2010,28(1):45-48
[22]艾希珍,于贤昌,王绍辉,等.低温胁迫下黄瓜嫁接苗与自根苗某些物质含量变化[J].植物生理学通讯,1999,35(1):26-28
[23]陈杰忠,徐春香,梁立峰.低温对香蕉叶片中蛋白质和脯氨酸的影响[J].华南农业大学学报,1999,20(3):54-58
[24]柴团耀,张玉秀.菜豆富含脯氨酸、蛋白质基因在生物和非生物胁迫下的表达[J].植物学报,1994,41(1):111-113
[25]李明玉,曹辰兴,于喜艳.低温锻炼对冷胁迫下黄瓜幼苗保护性酶的影响[J].西北农业学报,2006,15(1):160-164
[26]张纪林,谢晓金,教忠意,等.几种冬青属植物抗冻能力比较[J].园艺学报,2005,32(3):477-481
[27]高述民,陈朋军,郭惠红,等.日本桃叶珊瑚的冷驯化及抗寒机制研究[J].西北植物学报,2003,23(12):2113-2119
[28]周建,杨立峰,郝峰鸽,等.低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J].西北植物学报,2009,29(1):0136-0142
[29]郑国华,张贺英,钟秀荣.低温胁迫下枇杷叶片细胞超微结构及膜透性和保护酶活性的变化[J].中国生态农业学报,2009,17(4):739-745
[30]高京草,王长柱,王进国,等.枣树抗寒性测定方法研究[J].西北林学院学报,2011,26(5):72-75
[31]胡春霞,王丽,汤杰.低温对南果梨的生理生化指标的影响[J].沈阳农业大学学报,2009,40(3):349-352
[32]郑元,杨途熙,魏安智,等.低温胁迫对仁用杏几个抗寒生理指标的影响[J].西北农林科技大学学报:自然科学版,2008,36(1):163-167
[33] Prasad TK. Mechanism of chilling-induced oxidative stress injury and tolerance: changes in antioxidant system,oxidation of proteins and lipids and protease activities [J]. Plant J,1996,10(6):1017-1026
[34] Prasad TK,Anderson MP,Martin BA,et al. Evidence for chilling induced oxidative stress in maize seedling and a regulatory role for hydrogen peroxide [J]. Plant Cell,1994,6(1):65-74
[35]任安芝,高于葆,刘爽.铬、镉、铅胁迫对青菜叶片几种生理生化指标的影响[J].应用于环境生物学报,2000,6(2):112-116
[36]石雪晖,陈祖玉,杨会卿,等.低温胁迫对柑桔离体叶片中SOD及其同工酶活性的影响[J].园艺学报,1996,23(4):384-386
[37]詹福建,巫光宏,黄卓烈,等.马占相思树对低温冻害的抗性研究[J].林业科学,2003,39(1):56-61
[38]段肖翠,倪志云,路丙社,等.低温胁迫对阿月浑子抗寒生理指标的影响[J].河北农业大学学报,2005,28(4):48-50
[39]韩宏伟,张世红,徐兴兴,等.中国刺槐种源间抗寒性地理变异研究[J].河北农业大学报,2008,31(2):57-60
[40]柳新红,何小勇,苏冬梅,等.翅荚木种源抗寒性综合评价体系的构建与应用[J].林业科学,2007,43(10):45-50
The Effect of Cold Stress on the Cold Resistance Physiology of Two Camphor Species
WANG Ning1,YUAN Mei-li2,DING Bo2
1. College of Forestry/Henan University of Science and Technology, Luoyang 471000,China
2. The Sui&Tang Dynasties Relics Botanic Garden of Luoyang, Luoyang 471002,China
Abstract:Taking leaves of Cinnamonum bodinieri Levl and C. camphora L.(linalool-type)treated by cold temperature as test materials,the changes of MDA,Proline,Soluble protein and SOD activity,electrical conductivity in them were studied. The results indicated that for all the materials,with the decrease of temperature,the ion leakage percentage of the two species increased in S-shape,the SOD and POD activities decreased after an initial increase. The content of soluble protein,MDA and soluble sugar began to drop,then to increase and at last to drop again. By using fuzzy mathematics method,the cold resistance of two camphor tree species was evaluated as Cinnamonum bodinieri Levl>C. camphora L.(linalool-type),the semi-lethal temperature(LT50)of two camphor tree species calculated by the Logistic equation of ion leakage percentage was -12.43,-10.71℃,respectively. At last,by using a fuzzy mathematics method,the cold resistance of C. bodinieri Levl was divided into three periods,namely,the reducing stage,the enhancement setting stage,and the reducing stage.
Keywords:Camphor tree;cold resistance;SOD activity;comprehensive evaluation
作者简介:王宁(1979-),男,河南鄢陵人,讲师,博士,主要从事园林植物抗逆性相关研究. E-mail:hnkjdxwangning2013@163.com
基金项目:河南省教育厅高等学校重点科研项目(16A220001);河南科技大学青年科学基金项目(2013QN018)
收稿日期:2013-10-12修回日期:2013-12-18
中图法分类号:S722.7
文献标识码:A
文章编号:1000-2324(2016)01-0037-06
