不同土地利用方式下亚热带花岗岩小流域碳汇潜力及其影响因素
2016-04-05左双苗杨金玲张甘霖黄来明31土壤与农业可持续发展国家重点实验室中国科学院南京土壤研究所江苏南京210008中国科学院大学北京100049生态系统网络观测与模拟院重点实验室中国科学院地理科学与资源研究所北京100101
左双苗,杨金玲*,张甘霖,黄来明,31.土壤与农业可持续发展国家重点实验室//中国科学院南京土壤研究所,江苏 南京 210008;2.中国科学院大学,北京 100049;3.生态系统网络观测与模拟院重点实验室//中国科学院地理科学与资源研究所,北京 100101
不同土地利用方式下亚热带花岗岩小流域碳汇潜力及其影响因素
左双苗1,2,杨金玲1,2*,张甘霖1,2,黄来明1,2,3
1.土壤与农业可持续发展国家重点实验室//中国科学院南京土壤研究所,江苏 南京 210008;2.中国科学院大学,北京 100049;3.生态系统网络观测与模拟院重点实验室//中国科学院地理科学与资源研究所,北京 100101
摘要:了解小流域尺度上植物生长与岩石风化对CO2吸收的相对贡献对评估生态系统碳汇功能有重要意义,但过去的研究大多集中在某一单一过程且多基于特定土地利用类型。本研究以亚热带丘陵地区不同土地利用方式下的3个花岗岩小流域(F-100%森林、FA1-82%森林/18%农田和FA2-76%森林/24%农田)为研究区,自2010年3月至2012年2月定期监测了流域内的雨水、径流水,并采集了植物样品,分析其化学组成,系统研究了小流域尺度下植物生长和岩石风化的碳汇潜力及其影响因素。结果表明,F流域中不同林分(马尾松Pinus massoniana Lamb.阔叶树混交林、杉木Cunninghamia lanceolata(Lamb.) Hook.阔叶树混交林、马尾松林、杉木林、灌木林和竹Bambusoideae林)的碳密度和年均CO2吸收通量均不相同,碳汇潜力存在差异。不考虑施肥的影响,F、FA1和FA2流域植被的碳密度分别为44.5、37.3和35.0 t·hm-2,植被年均单位面积吸收CO2的量分别为10.5、11.8和12.4 t·hm-2·a-1,岩石风化消耗CO2的量分别为54.7×10-3、99.8×10-3和109.2×10-3t·hm-2·a-1,均随农田比例的增加而增加。施肥对农田水稻(Oryza sativa)碳截留的直接贡献很小,但可通过多种途径间接影响农田的碳汇潜力。3个流域径流水中HCO3-的物质的量浓度随农田比例增加而增加,在一定程度上受到施肥的影响,扣除施肥对径流水中HCO3-的贡献外,FA1和FA2流域土壤风化吸收CO2的量分别为84.4×10-3和88.6×10-3t·hm-2·a-1,仍高于F流域土壤风化吸收的大气CO2的量,说明农田土壤和森林土壤通过风化对CO2的固定存在差异。因此,农业活动在一定程度上影响了流域碳汇,不同土地利用方式下流域的碳汇潜力存在差异。尽管短时间尺度上植物生长对流域碳汇的贡献远高于岩石风化,但植物的收获与利用也可能加剧生态系统的碳排放,而硅酸岩风化在任何尺度上都是净碳汇,因而在地质时间尺度上硅酸盐风化对全球碳循环的影响不容忽视。
关键词:流域;植物固碳;岩石风化;风化固碳;碳汇潜力
ZUO Shuangmiao,YANG Jinling,ZHANG Ganlin,HUANG Laiming.Carbon Sequestration Potential in Granitic Watersheds under Different Land Uses in Subtropical China [J].Ecology and Environmental Sciences,2016,25(1):1-8.
工业革命以来,随着化石燃料的大量使用、森林砍伐和土地利用方式的改变,大量已被固定的碳被重新释放进入大气,使得大气CO2浓度呈持续上升趋势(Falkowski et al.,2000)。大气中CO2浓度已由工业革命前的280 mg·kg-1升高到2010年的389 mg·kg-1,并以每年约2 mg·kg-1的速度继续增加(刘再华,2012),由CO2排放增多带来的温室效应使得全球变暖已成为最严重的环境问题之一。研究发现目前已知的碳源和碳汇不能达到平衡,存在很大的碳失汇(刘建栋等,2007)。相比大气和海洋生态系统,同为碳汇的陆地生态系统更复杂,不确定性更大,因此寻找陆地碳汇成为当前研究的焦点(Gaillardet et al.,1999;韩冰等,2008;邱冬生等,2004)。
陆地生态系统中,充当碳汇的两个主要过程是岩石风化对大气CO2的消耗和植物光合作用对大气CO2的吸收(Gaillardet et al.,1999)。早在上世纪80年代初,Walker et al.(1981)即提出硅酸盐岩风化是大气CO2的一个主要的净汇。硅酸盐岩风化过程将CO2全部转化为HCO3-并将其作为碳酸盐矿物埋藏在海洋,已知碳酸盐矿物的存留时间超过百万年,因而在地质时间尺度上硅酸盐矿物的风化是消耗大气CO2的重要途径(刘建栋等,2007)。植物作为陆地生物圈碳循环的主体,通过光合作用吸收CO2制造碳氢化合物,将大气中的CO2以有机碳的形式固定于机体中,在减少全球CO2排放、维护区域乃至全球气候稳定中起着重要作用(Sedjo,1993;杨昆等,2006)。与此同时,岩石风化和植物生长相互影响,岩石风化可为植物提供矿质养分,间接促进植物光合作用对CO2的吸收;而植物通过分泌有机酸和吸收风化产物加速岩石风化进程,进而加快岩石风化对CO2的消耗(Berner,1992;Perrin et al.,2008;Hinsinger et al.,2001)。过去研究陆地生态系统的碳汇主要针对岩石风化和植物光合作用中的某一过程单独开展。此外,尽管前人分别针对林地和农田的碳汇进行了广泛研究(Fang et al.,2001;Schlesinger et al.,2001;Zhang et al.,2001),但是关于小流域尺度上不同土地利用方式下植物生长与岩石风化对CO2吸收的对比研究较少,不能系统地理解小流域尺度下岩石风化和植物吸收对碳汇的相对贡献。
本研究选取我国亚热带丘陵地区不同土地利用方式下的3个相邻流域(F-100%森林、FA1-82%森林/18%农田和FA2-76%森林/24%农田),通过定期监测与采集流域内雨水、径流水以及植物样并分析其化学组成,系统研究了小流域尺度上不同土地利用方式下岩石风化和植物吸收的碳汇潜力及其影响因素。
1 材料与方法
1.1研究区概况
研究区位于安徽省南部的泾县包合乡厚岸村,属亚热带湿润季风气候,年平均气温为16.5 ℃,年均降雨量为1585 mm。该地区最具代表性的岩石类型为花岗岩,其抗风化能力强,形成的土层较薄,主要土壤类型为湿润雏形土、正常新成土和水耕人为土。研究区内农作物主要为单季水稻(Oryza sativa)且休耕时不种植其它作物;而森林中林分类型较多,主要包括:马尾松Pinus massoniana Lamb.阔叶树混交林(I)、杉木Cunninghamia lanceolata (Lamb.) Hook.阔叶树混交林(II)、马尾松林(III)、杉木林(IV)、灌木林(V)和竹Bambusoideae林(VI)。除灌木林外,不同林分类型下灌木层和草本层物种丰富度差异性不大。根据调查结果,林下灌木树种主要有山矾Symplocos caudata、山胡椒Lindera glauca (Sieb.et Zucc.) Bl和檵木Loropetalum chinensis (R.Br.) Oliv.、野茶树Camellia assamica、桑树Morus alba L.和乌药Lindera aggregate (Sims) Kosterm.等,还有少量的六月雪Serissa japonica (Thunb.) Thunb.、野鸦椿Euscaphis japonica (Thunb.) Dippel、荆条Vitex negundo L.var.heterophylla (Franch.) Rehd.、茅栗Castanea seguinii Dode和映山红Rhododendron simsii Planch.。草本主要是铁芒萁Dicranopteris dichotoma、铁线蕨Adiantum capillus-veneris L.和显子草Phaenosperma globosa Munro ex Benth.等,而香附Cyperus rotundus L.、淡竹叶Lophatherum gracile、胡枝子Lespedeza bicolor Turcz.和白茅Imperata cylindrical (Linn.) Beauv.较少见。灌木林主要由山矾Symplocos caudata和矮竹Shibataea chinensis Nakai组成,草本极少。
选取不同土地利用方式下的3个相邻小流域(F-100%森林、FA1-82%森林/18%农田和FA2-76%森林/24%农田),流域的面积分别为359、72和101 hm2。2012年10月中旬对森林流域(F)手持GPS进行全流域踏查,确定了主要林分类型及其边界,选取了6个具有代表性的标准样地(选取依据详见本文1.2)(图1)。由于研究区3个小流域(F,FA1,FA2)紧连,且前期调查表明3个流域森林林分构成基本一致,因此本研究中FA1和FA2流域森林部分各指标参照纯森林流域F,并在FA1和FA2流域各选取了3块稻田标准样地(2 m×2 m)。
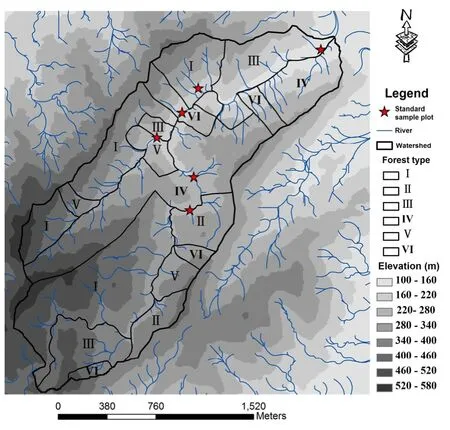
图1 森林流域 (F) 林分类型和标准样地示意图Fig.1 Schematic diagram of forest types and standard sample plot in the forest watershed (F)
1.2样品采集与分析
2010年3月至2012年2月对3个小流域(F,FA1,FA2)进行定期采样。雨水样品逢雨采用干湿沉降自动采样仪(青岛普仁PSC-Ⅲ)收集,用小型气象站(美国Watch Dog小型便携式自动气象站900ET)监测降雨量。在流域出口处分别修建了标准堰,用标准堰法测定径流量(黄来明等,2012),径流水样品采样周期为每周1次。
由于本研究区位于九华山和黄山风景区附近,林木采伐被限制,为尽量满足采样的科学性和代表性,在对整个流域卫星影像解读的基础上,对其进行实地踏勘,针对每个林分类型(马尾松阔叶树混交林、杉木阔叶树混交林、马尾松林、杉木林、灌木林和竹林),根据林分长势和立地条件分别选取3 个20 m×20 m的样地并对样地内的主要树种进行每木尺检,最终各林分分别选取主要树种的树高和胸径居中的样地为标准样地。根据标准样地每木尺检结果,在每个样地内,主要树种分别选取树高和胸径接近、高于和低于平均值的树木各1株作为标准木进行砍伐取样,选取的标准木具有较强的代表性。其中,马尾松林、杉木林和竹林各选3株标准木,马尾松阔叶树混交林选取马尾松和枫香Liquidambar formosana Hance各3株,杉木阔叶树混交林选取杉木和苦槠(Castanopsis sclerophylla (Lindl.) Schott.各3株。将选取的树木伐倒后分别对其干、枝和叶称重并取样。在样地的对角线4个角和中心设5个2 m×2 m小样方,记录样方内的灌木、草本种类,用全收获法分别测定鲜重并取样。灌木林仅需设小样方,用全收获法称重取样。2010年水稻收获时在FA1和FA2流域各随机选取3块稻田样地(2 m×2 m),在样地内用全收获法采集稻谷、稻秆和稻根样品。
雨水和径流水样品采集后立即用pH计(PHS型)测定pH值,过滤后立即用HCl滴定法测定HCO3-。植物样分析前先将样品清洗干净,置于80 ℃恒温烘箱内烘干后磨成粉备用。用碳氮分析仪测定所有植株中的碳。
1.3流域碳密度与CO2吸收通量计算方法
1.3.1植物碳密度及年均吸收CO2通量
不同植被碳密度根据森林或水稻的生物量(B)及其含碳率(Cc)计算得到,公式如下:

式中,C为植被碳密度(t·hm-2),Cc为含碳率(%),B为生物量(t·hm-2),其中B包括地上生物量(Ba)和地下生物量(Bu)。本研究中森林植被的Ba用收获法估算,具体步骤是在F流域按林分类型测定标准样地内树木的干、枝和叶,灌木、草本的地上部分及枯落物的鲜重,分别取样带回室内测干鲜比,算出样地的地上生物量,再乘以林分的相应面积即可求得。各林分面积通过手持GPS对F流域踏查确定相应的斑块边界,结合流域地形图用Arcgis软件定位获取,用该方法分别获取FA1和 FA2流域农田和森林面积。根据地下和地上部分生物量之比(Bu/Ba)乘以实测的地上生物量可估算地下生物量(姜萍等,2005)。由于多数植物的Bu/Ba值较稳定(Perala et al.,1994),本研究植物的地下部分生物量根据临近研究区相应植物的Bu/Ba值乘以实测的地上生物量获得(李海奎等,2010;张贵啅,2009;樊后保等,2006;孙刚,2009;汪永文等,2010)。水稻及森林植物地上部分的含碳率通过实测求得,森林植物根系的含碳率按如下原则替代:乔木用树干,灌木用灌木干枝,草本用地上部分替代。FA1和FA2流域中水稻生物量根据小样方(2 m×2 m)收获的水稻根系、籽粒和秸秆的鲜重,室内测定的干鲜比及农田面积计算而得,森林部分各项指标参照F流域。
各流域单位面积植被年均CO2吸收量可用下述公式估算:
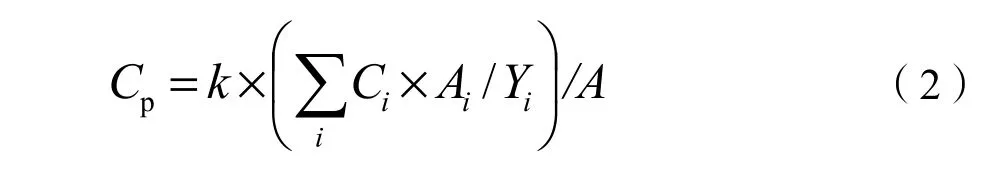
式中,Cp为流域单位面积植被年均CO2吸收量(t·hm-2·a-1),Ci为植被类型i的碳密度(t·hm-2),Ai为植被i的面积(hm2),Yi为植被i的年龄(a),A为流域面积(hm2),k为CO2与C的摩尔质量比(3.67)。马尾松阔叶树混交林、杉木阔叶树混交林、马尾松林、杉木林和灌木林的平均生长年龄根据具有平均树高和胸径的优势树种的地径年轮估算,竹林的平均年龄由有经验的植物学家判断得来,水稻以一年计。
1.3.2花岗岩风化消耗CO2的量
根据Yang et al.(2013)的研究,本流域内花岗岩的主要矿物为斜长石,可用化学反应式:2NaAlSi3O8+2CO2+11H2O→2Na++2HCO3-+4H4SiO4+Al2Si2O5(OH)4来表示。由此可见,研究区岩石的碳汇功能实际上是1 mol大气或土壤中的CO2通过风化转变为1 mol碳酸盐的过程,该过程中有1 mol CO2的净消耗。根据风化产生的碳酸盐量,利用水化学方法,参照刘玉等(2008)的公式,由于研究区雨水呈较强酸性检测不到无机碳存在,而径流水中的无机碳以HCO3-的形态为主,故本研究区花岗岩单位面积风化消耗的CO2量可简化为:
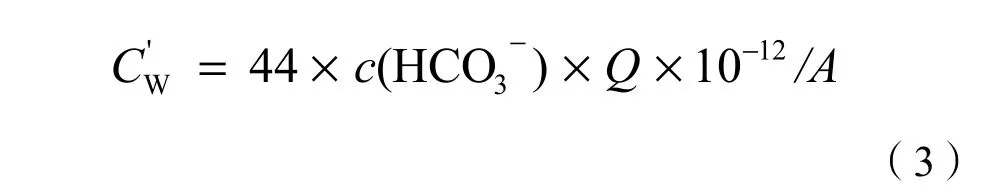
式中,C’w代表单位面积岩石风化消耗CO2的量(t·hm-2·a-1),44为CO2的摩尔质量(g·mol-1),[c(HCO3)-为径流水中HCO3-年加权平均物质的量浓度(μmol·L-1),Q代表年加权平均流量(L·a-1),A代表流域面积(hm2)。
2 结果
2.1流域碳汇
2.1.1植物含碳率分析
研究区乔木的含碳率都在40%以上,其中马尾松阔叶树混交林(I)中马尾松的加权平均含碳率最高,而枫香最低(表1),研究区乔木的含碳率略低于前人在安徽池州和江西千烟洲人工林的研究结果(张乐勤,2011;沈文清等,2006)。同种乔木的不同器官(干、枝、叶)间及不同乔木的含碳率差异都很小,变异系数均小于5%(表1)。
林下植被层中,灌木枝条、灌木叶、草本和枯落物的含碳率均值分别为44.8%、41.7%、42.0%和45.0%,各林分间的变异系数分别为1.5%、3.5%、2.0%和5.5%。不同林分中,枯落物因其组成成分差异大,含碳率变异系数最高。流域内不同林分及其枯落物含碳率高低表现为:乔木层>枯落物>灌木>草本,与前人在江西人工林的研究结果一致(沈文清等,2006)。水稻样品中,稻根、稻谷和稻秆含碳率均值分别为37.3%、39.0%和37.1%。
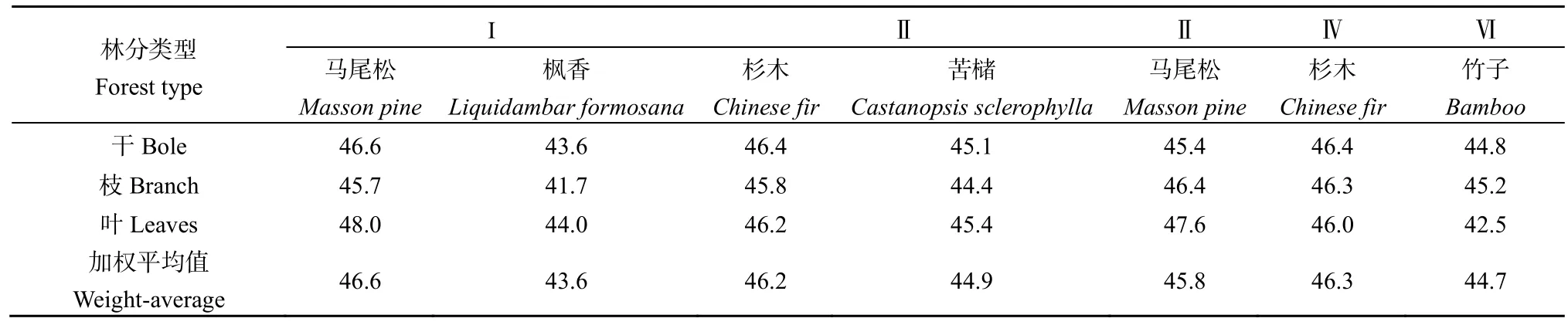
表1 流域内乔木的含碳率Table 1 Carbon content rates of trees in watershed %
2.1.2流域内植被的碳储量
由于3个小流域的森林林分类型相同且各林分的面积占森林总面积比例基本一致,因此本研究以森林流域(F)为例估算不同林分的碳密度。研究区不同林分地上部分碳密度存在较大差异(表2),其中以马尾松纯林(III)最高(56.5 t·hm-2),竹林(VI)最低(10.4 t·hm-2)。流域(F)内不同林分地上部分的总碳储量为13.2×103t,其中覆盖面积最大的马尾松阔叶树混交林(I)贡献最高(39.2%),而占流域面积最少的竹林贡献最低(1.7%)(表2)。F流域单位面积地上部碳密度平均值为36.9 t·hm-2。
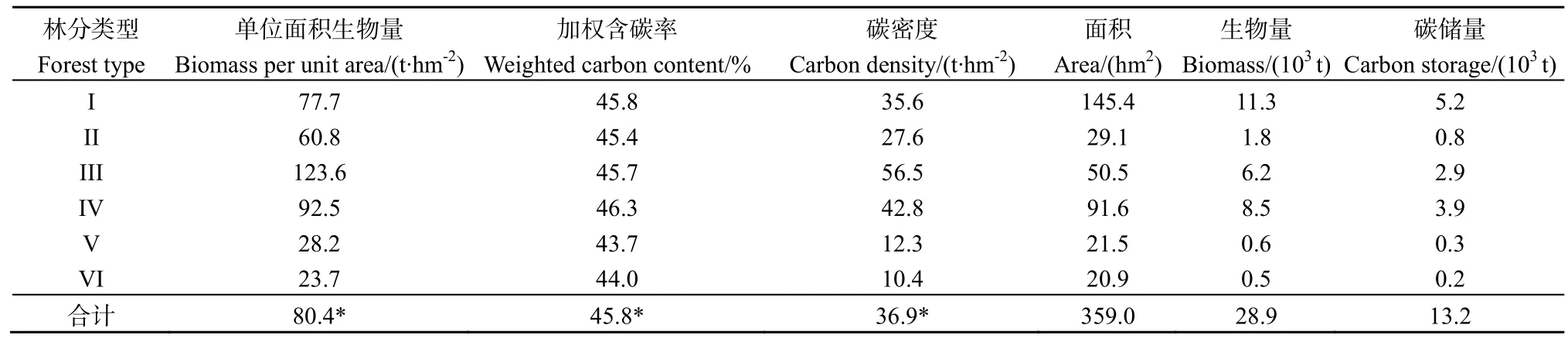
表2 F流域植被地上部分的生物量和碳储量Table 2 Above-ground biomass and carbon storage of plants in the forested watershed (F)
流域内不同林分地下生物量占总生物量的比例为12.4%~28.5%,各林分地下部分碳储量占总碳储量的比例为12.4%~28.6%,与生物量的变化规律一致(图2)。F流域植被地上与地下部分碳储量之和为16.0×103t,植被碳密度为44.5 t·hm-2,安徽省乔木林碳密度为29.0 t·hm-2(李海奎等,2010),研究区森林碳密度是安徽省乔木林平均碳密度的1.5倍,与临近区域池州的研究结果相近(张乐勤,2011)。
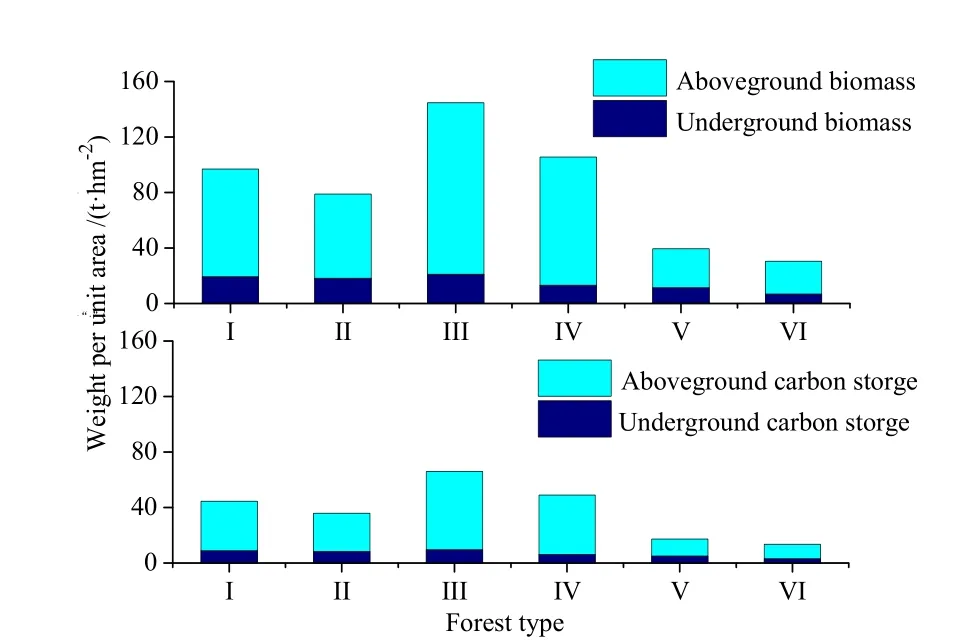
图2 不同林分类型单位面积生物量和单位面积碳储量Fig.2 Biomass and carbon storage per unit area of different vegetation types
农林流域(FA1,FA2)内水稻的碳密度为4.9 t·hm-2,低于任意林分类型。农林流域(FA1,FA2)植物的碳储量通过农田和森林的面积及其碳密度求得。FA1和FA2流域植物的碳储量分别为2.7×103和3.5×103t,碳密度分别为37.3和35.0 t·hm-2,均低于森林流域(F)。
2.1.3植物年均吸收大气CO2的量
研究区各林分年均吸收大气CO2的量相差较小,其中以杉木阔叶树混交林最大为11.9 t·hm-2·a-1,灌木林最小为7.9 t·hm-2·a-1(图3),与前人在闽江流域的研究结果差别不大(姬桂珍等,2003)。森林流域(F)植物年均吸收CO2的量为10.7 t·hm-2·a-1,与曾曙才等(2003)提出的生产1 g干物质需要吸收1.63 g CO2(10.18 t·hm-2·a-1)的结果基本一致。研究区水稻年均吸收的CO2量为17.9 t·hm-2·a-1,与按曾曙才等(2003)的方法计算的结果20.99 t·hm-2·a-1相差较大,说明该方法可能更适于计算森林植物CO2的吸收量。FA1和FA2流域内植物年均吸收的CO2量根据农田和森林在流域中的面积及农田和森林单位面积年均吸收的CO2量求得,分别为11.8和12.4 t·hm-2·a-1,流域中农田比例越高则流域内植被单位面积年均吸收的CO2量越高。
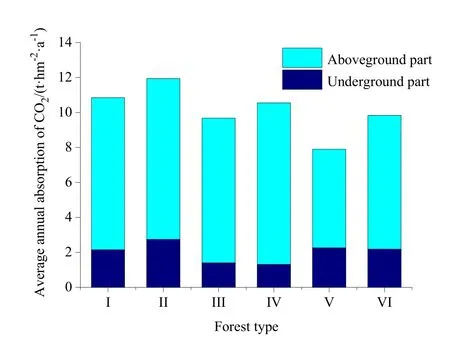
图3 不同林分类型年均吸收大气CO2的量Fig.3 Average annual absorption of CO2of different forest types
2.2流域花岗岩风化过程消耗的CO2量
2.2.1径流水中HCO3-的物质的量浓度
监测期间,F、FA1和FA2流域径流水中HCO3-变异系数分别为34.4%、30.8%和36.5%,3个小流域径流水中HCO3-物质的量浓度最高值与最低值均相差10倍以上,说明径流水中HCO3-物质的量浓度受环境气候条件等的影响而差异较大(图4)。3个流域(F、FA1和FA2流域)的HCO3-含量加权平均物质的量浓度随农田比例的增加而增加,分别为178.7,310.8和360.7 μmol·L-1(图4)。
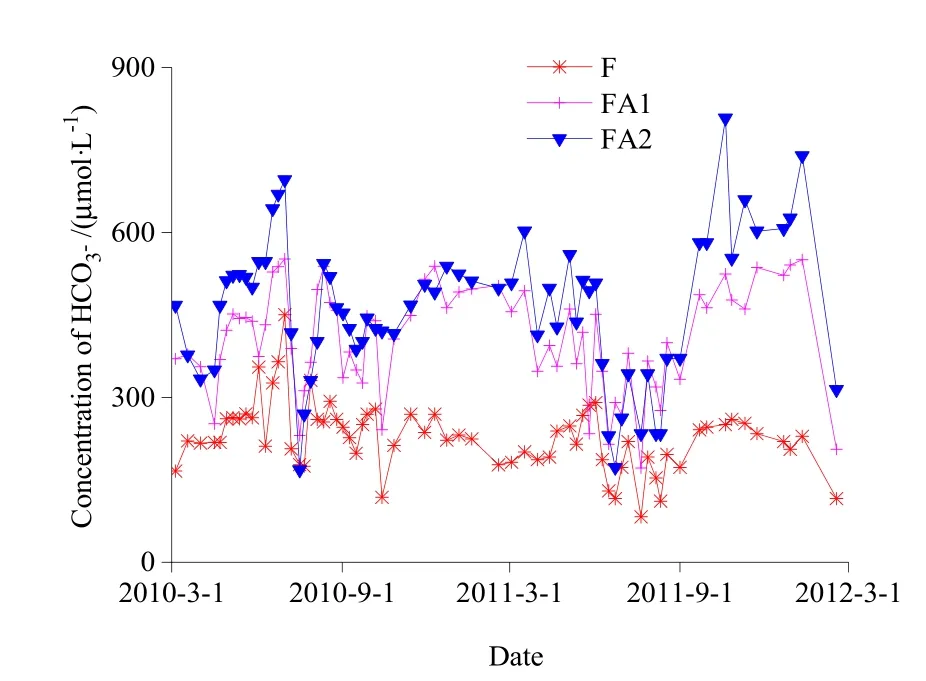
图4 径流水中HCO3-浓度动态变化Fig.4 Dynamic change of concentration of HCO3- in runoff water
监测期间,径流水中HCO3-物质的量浓度大小的一般趋势为FA2>FA1>F,2010年度和2011年度径流水中HCO3-季平均物质的量浓度变化规律分别为:F流域(夏季>春季>秋季>冬季)、FA1和FA2流域(冬季>夏季>春季>秋季);F流域(秋季>春季>冬季>夏季)、FA1和FA2流域(秋季>冬季>春季>夏季)。结果表明,含部分农田的两个流域(FA1和FA2)的HCO3-季平均物质的量浓度变化规律一致,而与纯森林流域(F)不同,说明土地利用方式的不同可能会影响岩石矿物风化消耗大气CO2的量。
2.2.2岩石风化消耗大气CO2的量
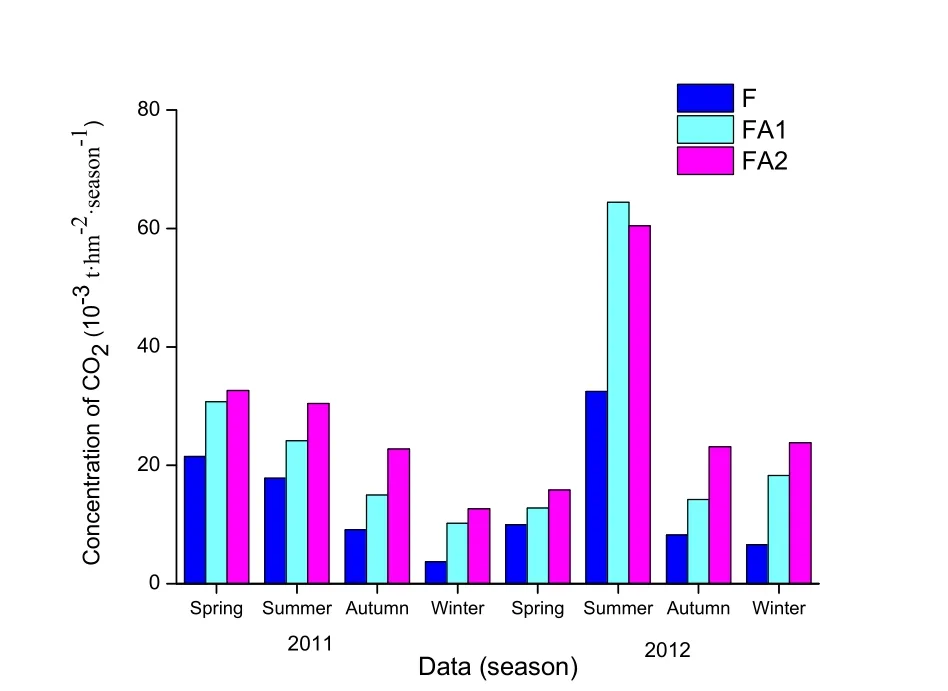
图5 各流域岩石风化消耗CO2的量 (季度)Fig.5 Seasonal adsorption of CO2of rock weathering in all watersheds
若将径流输出的HCO3-全部归因于岩石风化消耗大气CO2,则3个流域岩石风化吸收大气CO2的趋势为FA2>FA1>F,这与流域径流水中HCO3-的物质的量浓度的变化规律一致(图5)。3个流域(F、FA1和FA2),2010年岩石风化吸收大气CO2量的规律均为春季>夏季>秋季>冬季,而2011年的规律为:F流域(夏季>春季>秋季>冬季)、FA1和FA2流域(夏季>冬季>秋季>春季),这与各流域径流水中的HCO3-季物质的量浓度的规律不完全一致。监测期间,F、FA1和FA2流域岩石风化消耗大气CO2的量与降雨量的相关性分别为0.96(n=8,P<0.01)、0.99(n=8,P<0.01)和0.97(n=8,P<0.01),均具有明显的相关性,说明流域岩石风化吸收大气CO2还受降雨量的影响。监测期间,流域(F,FA1和FA2)岩石风化吸收的大气CO2通量分别为54.7×10-3、99.8×10-3和109.2×10-3t·hm-2·a-1,与我国花岗岩单位面积平均碳汇量的数值相近(刘玉,2006),但3个流域呈现出随农田比例增加而增加的规律(图5)。
3 讨论
3.1农业活动对流域碳汇的影响
本研究选取的3个小流域土地利用方式不同,3个流域中植物年均吸收大气CO2的量随流域农田比例的增高而增加,而流域内植被碳密度表现出相反的规律。这是因为虽然FA1和FA2流域中水稻的碳密度低于森林各林分类型中植被的碳密度,但由于水稻的生长周期仅1年,其年均吸收的CO2量高于所有森林林分类型,因而3个流域中植被碳密度和年均吸收大气CO2的量随农田比例不同呈现出相反的规律。
根据调查,研究区农田在水稻生长期间会被施入尿素,按照当地的尿素施用量,随尿素进入FA1 和FA2流域中农田的碳为39.0×10-3t·hm-2·a-1,由于水稻对尿素的利用率为25%~40%(林新坚等,1992),假定40%施入的尿素被水稻吸收,将这部分碳折算为水稻对CO2的截留量为57.2×10-3t·hm-2·a-1,水稻生长年截留的CO2是该数值的200多倍,这说明施肥对水稻碳截留的直接贡献很小。但施肥可通过促进水稻生长,提高其光合作用效率从而间接增加水稻对大气CO2的截留,此外,通过改进和优化耕作措施,如采用保护性耕作措施、扩大水田种植面积、增加秸秆还田和增加有机肥施用等,可以减少农田土壤的CO2净排放,稳定甚至增加土壤有机碳贮量(韩冰等,2008)。前人研究表明,近年来我国南方稻田生态系统土壤有机碳含量普遍呈升高趋势,表现为重要的碳汇(张琪等,2004),因此潘根兴(2008)认为,我国农田具有显著的固碳减排潜力,其中稻作农业的固碳潜力十分突出。
FA1和FA2流域径流水中HCO3-物质的量浓度显著高于森林流域(F)也与农田施肥有关,这是因为尿素施入土壤后,除少量以分子态被土壤胶体吸附外,大部分在土壤脲酶的作用下,水解成碳酸铵(胡霭堂等,2003)。由于稻田氮肥利用率仅25%~40%,剩余的氮肥除氨挥发外,一部分转化为碳酸氢铵的形式进入径流水中。流域农田比例越高,施肥量越多,因此研究区3个流域径流水中HCO3-的物质的量浓度呈现随农田比例增加而增加的趋势。假定尿素中碳被水稻利用外全部以HCO3-的形式进入径流水中,扣除施肥对径流水中HCO3-的贡献外,FA1和FA2流域土壤风化吸收的大气CO2量分别为84.4×10-3和88.6×10-3t·hm-2·a-1,仍高于森林流域(F)土壤风化吸收的大气CO2的量(54.7×10-3t·hm-2·a-1),说明农田土壤和森林土壤通过风化对大气CO2的固定本身也存在一定的差异。
3.2植物生长与岩石风化消耗大气CO2的差异
监测期间,研究区3个小流域(F,FA1和FA2)中植物生长吸收大气CO2的量均为岩石风化消耗大气CO2的100多倍,因此,植物生长消耗大气CO2的量远高于岩石风化的贡献。这一方面与研究区植被的覆盖度高、植物单位面积的生物量相对较高有关;另一方面由于研究区的母岩为花岗岩,属于溶解速度较慢的硅酸盐类(刘再华,2012),且本研究未测定径流水中的有机碳,而前人研究(刘再华等,2012)表明相当一部分输出的有机碳源于岩石风化的贡献,本研究在一定程度上了低估岩石风化的碳汇潜力。
值得注意的是,虽然监测期间研究区植物固定的碳远高于岩石风化固定的碳,但是植物被利用的过程同时也是CO2排放的过程(徐德应等,1992)。例如森林被砍伐后,其地下生物量未被有效利用,而地上部分的树叶和树皮等通常也未被利用,加工后的木制品只有少部分“永久地”保存下来,其余最终以CO2的形式归还大气。同时,土地利用方式改变、生态系统退化可导致生态系统由碳汇变为碳源,如每年我国森林砍伐开荒所释放的CO2量为1.66 t(刘玉,2006)。此外,研究表明全球每年约1%的森林遭受火干扰(Crutzen et al.,1979),森林火灾排放到大气中的碳相当于每年化石燃料燃烧排放量的70%(Taylor et al.,1990),而随着全球气候变暖,森林火灾频率和强度将加剧(Running,2006),势必进一步弱化森林的碳汇潜力。因此,虽然短时间尺度上由于森林的总碳贮量很大,它的微小变化就可以引起大气中CO2浓度的较大波动,但从长时间尺度来看森林对大气碳的作用较小。硅酸盐岩风化过程是将大气中的CO2全部转化为HCO3-并随着径流水进入海洋,将其作为碳酸盐矿物埋藏在海洋,而转化后的碳酸盐矿物的存留时间可超过百万年,因而在地质时间尺度上硅酸盐矿物的风化是消耗大气CO2的重要途径,该理论已被大量科学家所验证和认可(刘建栋等,2007;Gaillardet et al.,1999;Walker et al.,1981;李海奎等,2010;刘玉等,2007;邱冬生等,2004),因此在长时间尺度上岩石风化的碳汇潜力不容忽视,今后要加强对这部分碳汇的研究。
在陆地生态系统中,岩石风化和植物生长对大气CO2的吸收作用是相互影响的,岩石的风化产物能给植物生长提供矿质养分,通过促进植物光合作用间接提高对大气CO2的吸收;而植物通过分泌有机酸和吸收风化产物能加速岩石风化进程,进而加快岩石风化对CO2的消耗(Berner,1992;Perrin et al.,2008;Hinsinger et al.,2001),因此,系统研究植被生长和岩石风化的碳汇潜力是很有必要的。
4 结论
(1)3个流域(F,FA1和FA2)中植被的碳密度分别为44.5、37.3和35.0 t·hm-2,年均吸收大气CO2的量依次为10.3、11.4和12.4 t·hm-2·a-1,随流域农林比例的递增碳密度和CO2吸收量表现出相反的变化趋势。施肥对农田水稻碳截留的直接贡献很小,但可通过多种途径间接影响农田的碳汇潜力。不考虑施肥的影响,F、FA1和FA2流域径流水中HCO3-的物质的量浓度和岩石风化吸收大气CO2的量分别为178.7、310.8和360.7 μmol·L-1和54.7×10-3、99.8×10-3和109.2×10-3t·hm-2·a-1,均随农田比例增加而增加。扣除施肥对径流水中HCO3-的贡献外,FA1和FA2流域土壤风化吸收的大气CO2量分别为84.4×10-3和88.6×10-3t·hm-2·a-1,仍高于森林流域(F)土壤风化吸收的大气CO2的量,说明农田土壤和森林土壤通过风化对大气CO2的固定本身也存在一定的差异。因此,农业活动在一定程度上影响了流域碳汇,研究区流域内植被和岩石风化的碳汇潜力因土地利用方式不同而异。
(2)监测期间,3个流域植物年均吸收的大气CO2量均远高于岩石风化年均消耗的大气CO2量,短时间尺度内流域碳汇潜力主要受植物生长的影响。但植物砍伐、土地利用方式的转变等可能加剧生态系统内碳的排放,而硅酸盐岩风化在地质时间尺度上是CO2的净碳汇,二者在短时间尺度上相差较大,但从长时间尺度上的比较还需要更系统的评估。
参考文献:
BERNER R A.1992.Weathering plants and the long term carbon cycle [J].Geochimica Et Cosmochimica Acta,56(13):3225-3231.
CRUTZEN P J,HEIDT L E,KRASNEC J P,et al.1979.Biomass burning as a source of atmospheric gases CO,H2,N2O,NO,CH3Cl and COS [J].Nature,282(5736):253-256.
FALKOWSKI P,SCHOLES R J,BOYLE E,et al.2000.The global carbon cycle:a test of our knowledge of earth as a system [J].Science,290(5490):291-296.
FANG J Y,CHEN A P,PENG C H,et al.2001.Changes in forest biomass carbon storage in China between 1949 and 1999 [J].Science,292(5525):2320-2322.
GAILLARDET J,DUPRÉ B,LOUVAT P,et al.1999.Global silicate weathering and CO2consumption rates deduced from the chemistry of large rivers [J].Chemical Geology,159(1-4):3-30.
HINSINGER P,BARROS O N F,BENEDETTTI M F,et al.2001.Plant-induced weathering of a basaltic rock:Experimental evidence [J].Geochimica Et Cosmochimica Acta,65(1):137-152.
PERALA D A,ALBAN D H.1994.Allometric biomass estimations for aspen-dominated ecosystems in the Upper Great Lakes [M].USDA Forest service Research paper:NC-134.
PERRIN A S,PROBST A,PROBST J C.2008.Impact of nitrogenous fertilizers on carbonate dissolution in small agricultural catchments:Implications for weathering CO2uptake at regional and global scales [J].Geochimica Et Cosmochimica Acta,72(13):3105-3123.
RUNNING S W.2006.Is global warming causing more,larger wildfires [J].Science,313(5789):927-928.
SCHLESINGER W H,LICHTER J.2001.Limited carbon storage in soil and litter of experimental forest plots under increased atmospheric CO2[J].Nature,411(6836):466-469.
SEDJO R A.1993.The carbon cycle and global forest ecosystem [J].Water,Air,and Soil Pollution,70(1-4):295-307.
TAYLOR,L R.,PICKET,S T A.,WHITE,P S.1990.The ecology of natural disturbance and patch dynamics [J].Journal of Animal Ecology,59(4724):434-435.
WALKER J C G,HAYS P B,KASTING J F.1981.A negative feedback mechanism for the long-term stabilization of Earth's surface temperature chemistry of large rivers [J].Journal of geophysical research,86(C10):9776-9782.
YANG J L,ZHANG G L,HUANG L M,et al.2013.Estimating soil acidification rate at watershed scale based on the stoichiometric relations between silicon and base cations [J].Chemical Geology,s 337-338(1),30-37.
ZHANG F,LI C,WANG Z,et al.2001.Modeling impacts of management alternatives on soil carbon storage of farmland in Northwest China [J].Biogeosciences,3(4):451-466.
曾曙才,苏志尧,谢正生,等.2003.广州白云山主要林分的生产力及吸碳放氧研究[J].华南农业大学学报,24(1):16-19.
樊后保,李燕燕,苏兵强,等.2006.马尾松-阔叶树混交异龄林生物量与生产力分配格局[J].生态学报,26(8):2463-2473.
韩冰,王效科,逯非,等.2008.中国农田土壤生态系统固碳现状和潜力[J].生态学报,28(2):612-619.
胡霭堂,周立祥.2003.植物营养学(下册)[M].北京:中国农业大学出版社:22-24.
黄来明,张甘霖,杨金玲.2012.亚热带典型花岗岩小流域径流化学特征与化学风化[J].环境化学,31(7):1-8.
姬桂珍,吴承祯,洪伟,等.2003.闽江流域主要林分生产力的空间分布格局及其吸碳放氧量的研究[J].江西农业大学学报,25(6):828-833.
姜萍,叶吉,吴钢.2005.长白山阔叶红松林大样地木本植物组成及主要树种的生物量[J].北京林业大学学报,27(S2):112-115.
李海奎,雷渊才.2010.中国森林植被生物量和碳储量评估[M].北京:中国林业出版社:42-48.
林新坚,陈济深,郑时利,等.1992.水稻土中脲酶产生菌、脲酶活性及尿素利用率[J].福建省农科院学报.7(l):36-40.
刘建栋,胡泓,张龙军.2007.流域化学风化作用的碳汇机制研究进展[J].土壤通报,38(5):998-1002.
刘玉,刘德深,沈立成.2007.云南维西花岗岩地区水文地球化学特征及形成机制研究[J].地球化学,36(2):161-170.
刘玉,刘德深,沈立成.2008.花岗岩地区碳汇计算及影响因素研究[J].地球化学,37(3):281-289.
刘玉.2006.花岗岩风化碳汇规律及影响因素研究[D].重庆:西南大学:53-63.
刘再华,DREYBROD W.2012.碳酸盐风化碳汇与森林碳汇的对比——碳汇研究思路和方法变革的必要性[J].中国岩溶,31(4):345-348.
刘再华.2012.岩石风化碳汇研究的最新进展和展望[J].科学通报,57(2-3):95-102.
潘根兴.2008.中国土壤有机碳库及其演变与应对气候变化[J].气候变化研究进展,4(5):282-289.
邱冬生,庄大方,胡云锋,等.2004.中国岩石风化作用所致的碳汇能力估算[J].地球科学—中国地质大学学报,29(2):177-182,190.
沈文清,刘允芬,马钦彦,等.2006.千烟洲人工针叶林碳素分布、碳贮量及碳汇功能研究[J].林业实用技术,(8):5-8.
孙刚.2009.安徽肖坑天然毛竹林生产力及养分特征的研究[D].合肥:安徽农业大学:15-16.
汪永文,王力,王丽丽,等.2010.马尾松混交林林下植被结构及生物量特征研究[J].安徽农业大学学报,37(2):312-316.
徐德应,刘世荣.1992.温室效应、全球变暖与林业[J].世界林业研究,5(1):25-32.
杨昆,管东生,周春华.2006.潭江流域森林碳储量及其动态变化[J].应用生态学报,17(9):1579-1582.
易利萍.2008.枫香人工林生态系统生物量和养分的积累与分布[D].长沙:中南林业科技大学:17-21.
张贵啅.2009.杉木混交林生长与生物量研究[J].安徽农学通报,15(21):131-132.
张乐勤.2011.安徽池州森林植被碳贮量调查及分析[J].植物学报,46(5):544-551.
张琪,李恋卿,潘根兴,等.2004.近20年来宜兴市城水稻土有机碳动态及驱动因素[J].第四纪研究,24(2):236-242.
Carbon Sequestration Potential in Granitic Watersheds under Different Land Uses in Subtropical China
ZUO Shuangmiao1,2,YANG Jinling1,2*,ZHANG Ganlin1,2,HUANG Laiming1,2,3
1.State Key Laboratory of Soil and Sustainable Agriculture,Institute of Soil Science,Chinese Academy of Sciences,Nanjing 210008,China; 2.University of Chinese Academy of Sciences,Beijing 100049,China; 3.Key Laboratory of Ecosystem Network Observation and Modeling,Institute of Geographic Sciences and Natural Resources Research,Chinese Academy of Sciences,Beijing 100101,China
Abstract:It is critical to understand the relative contributions of plant assimilation and rock weathering to carbon sinks at watershed scale for the quantitative assessment of the carbon sequestration potential of ecosystems.This study used three adjoining granite watersheds with different land use types (F-100% forest,FA1-82% forest/18% farmland and FA2-76 % forest/24% farmland) as pilot areas for a comprehensive monitoring and quantification of carbon cycling fluxes.The rainwater and surface runoff of watersheds were monitored from March,2010 to February,2012.Plant samples were collected and related chemical properties were determined.Carbon sequestration potential and its influencing factors in small-scaled watersheds were investigated systemically.The results showed that the carbon storage and average annual adsorption of CO2of six different forest types (Masson pine (Pinus massoniana Lamb.) and broad-leaved mixed forest,Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) and broad-leaved mixed forest,Masson pine forest,Chinese fir forest,Shrub forest and Bamboo (Bambusoideae) forest) in F watershed were different.Ignoring the contribution of fertilization,carbon density of vegetation,average annual absorption of CO2of vegetation and annual absorption of CO2of rock weathering in the three watersheds (F,FA1 and FA2) were 44.5,37.3 and 35.0 t·hm-2,10.5,11.8 and 12.4 t·hm-2·a-1and 54.7×10-3,99.8×10-3and 109.2×10-3t·hm-2·a-1,respectively,which all increased with higher proportion of farmland in the watersheds.Though the direct contribution of fertilization in farmlands was minor,fertilization could indirectly increase carbon sink potential by many mechanisms.The concentration of HCO3-in the three watersheds increased with higher farmland proportion,suggesting that HCO3-concentrations in FA1 and FA2 were affected by fertilization to some extent.Subtracting the contribution of fertilization,annual absorption of CO2of rock weathering in FA1 and FA2 watershed were 84.4×10-3and 86.9×10-3t·hm-2·a-1,which were still both higher than that in the forested watershed (F).This indicated that there was difference between carbon sequestration by weathering in farmland soil and forest soil.The results demonstrated that agricultural activities may affect carbon sequestration potential of watershed to some extent.Although the contribution of vegetation to sequestration of CO2was much higher than rock weathering over short time scales,harvest and utilization of vegetation can intensify the emission of carbon in ecosystem.By contrast,rock weathering is a net carbon sink over both short and long time scales,so its influence on global carbon cycling should not be ignored.
Key words:watershed; CO2fixation by plants; rock weathering; carbon sequestration by weathering; carbon sequestration potential
收稿日期:2015-11-05
*通信作者:杨金玲(1973年生),女,副研究员,博士,研究方向为土壤发生和土壤地球化学。E-mail:jlyang@issas.ac.cn
作者简介:左双苗(1986年生),女,博士研究生,研究方向为土壤地球化学。E-mail:smzuo@issas.ac.cn
基金项目:国家自然科学基金项目(41471176;41071141)
中图分类号:X14
文献标志码:A
文章编号:1674-5906(2016)01-0001-08
DOI:10.16258/j.cnki.1674-5906.2016.01.001
引用格式:左双苗,杨金玲,张甘霖,黄来明.不同土地利用方式下亚热带花岗岩小流域碳汇潜力及其影响因素[J].生态环境学报,2016,25(1):1-8.
