Paleocene faunal evolution at Polecat Bench in the northern Bighorn Basin of Wyoming, USA
2016-03-29PhilipGINGERICH
Philip D. GINGERICH
(Museum of Paleontology,University of MichiganAnn Arbor, Michigan 48109-1079, USA gingeric@umich.edu)
Paleocene faunal evolution at Polecat Bench in the northern Bighorn Basin of Wyoming, USA
Philip D. GINGERICH
(Museum of Paleontology,University of MichiganAnn Arbor, Michigan 48109-1079, USA gingeric@umich.edu)
Polecat Bench and McCullough Peaks in the northern Bighorn Basin of Wyoming have yielded major discoveries leading to understanding both mammalian biostratigraphy of the North American Paleocene, and the pattern and cause of continental faunal turnover at the Paleocene-Eocene thermal maximum. In 1910, Princeton and American Museum paleontologists showed that mammals of the Paleocene were distinctly different from those of the Eocene. This soon led to acceptance of the Paleocene as an epoch different from the Eocene. Eighty years later, Michigan and Carnegie Institution paleontologists showed that the continental Paleocene-Eocene boundary was marked by abrupt faunal turnover and dwarfing associated with a major carbon isotope excursion. This led to recognition of the Paleocene-Eocene thermal maximum or ‘PETM’as a greenhouse warming event that was truly global. A young scholar involved in paleontology in 1950, Minchen Chow, was introduced to fi eld work on Paleocene mammals in Wyoming. There he developed the experience and expertise that led to parallel exploration of the Paleocene in China. The challenge now is to understand how the parallel Paleocene histories of the North American and Asian continents fi t together.
Polecat Bench, McCullough Peaks, Paleocene, Eocene, biochronology, dwar fi ng, carbon isotope excursion, PETM
1 Introduction
Major advances in paleontology were made in the last century, and the Paleocene history I review here spanned most of the twentieth century, from 1910 to the end. It is a story of exploration, persistence, and good fortune involving four leaders (Fig. 1) and many active followers.
The northern Bighorn Basin is important to the history of Paleocene research for fourreasons: 1) this is the area where the Paleocene was fi rst shown effectively to be different from the Eocene; 2) this is the area where a succession of Paleocene mammalian faunas enabled some of the North American Paleocene land-mammal ages to be subdivided into biochrons based on superposition and evolutionary succession; 3) this is the area where the Paleocene-Eocene transition was fi rst shown to be a sharp boundary, with dwar fi ng of some mammalian lineages at the boundary; 4) this is the area where the carbon isotope excursion marking the Paleocene-Eocene Thermal Maximum (PETM) was fi rst found on land, showing that (a) the greenhouse warming event marking the end of the Paleocene and beginning of the Eocene was global, and (b) linking continental and marine Paleocene-Eocene time scales. Each of these is described in turn.

Fig. 1 Leaders in early research on the Paleocene mammals of western North America and China:William J. Sinclair, Walter Granger, Glenn L. Jepsen, and Minchen Chow A. William Sinclair (1877-1935) of Princeton University in Princeton, New Jersey; photograph courtesy of the Department of Geosciences, Princeton University. B. Walter Granger (1872-1941) of the American Museum of Natural History; photograph courtesy of the Vertebrate Paleontology archives, American Museum of Natural History in New York, New York. C. Glenn Jepsen (1903-1974) of Princeton University; photograph courtesy of the Department of Geosciences, Princeton University. D. Minchen Chow (1918-1996) of Lehigh and Princeton universities, and the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences in Beijing; photograph courtesy of the Chow family, from Mulzet Studio, Bethlehem, Pennsylvania, circa 1950
The Bighorn Basin and the Clarks Fork Basin are distinct river basins separated by the ‘Clarks Fork Divide,’ which includes Polecat Bench (Fig. 2). The Clarks Fork River is a tributary of the Yellowstone River farther north in Montana, while the Shoshone River south of the divide is a tributary of the Bighorn River (which also eventually joins the Yellowstone River in Montana). Both drainage basins are part of a larger Bighorn structural basin, and I will refer to both as the northern Bighorn Basin.
3.5.3 间苗补苗 为培养充足优质壮苗,根据苗木生长状况要及时间苗和补苗。间苗的原则是“留好去坏,留稀去稠”。间苗对象为病苗、受伤苗、弱苗和非常稠密的苗木。补苗要选在阴雨天或傍晚进行。补苗后应及时浇水,使苗木根系与土壤密结,必要时可适当遮阴。
Early explorers involved in Paleocene research in the Bighorn Basin came from American institutions, but the in fl uence was broader. A young Chinese scholar involved in the research returned to China with experience that led to parallel investigation of mammalian evolution in Asia. This review complements one published earlier (Gingerich, 2010). Here I focus on North America and Asia, but the European Paleocene has its own rich record (Russell, 1964), as does the Paleocene of South America (Clyde et al., 2014; Woodburne et al., 2014) and that of Africa(Gheerbrant, 1992; Gheerbrant and Rage, 2006).

Fig. 2 Map of Paleocene and Early Eocene strata in the northern Bighorn Basin of northwestern Wyoming, U.S.A., showing the distribution of University of Michigan fossil localities, which include sites visited by earlier American Museum of Natural History and Princeton University expeditions Open symbols are fossil localities yielding Paleocene mammals of the Puercan (Mantua Quarry square), Torrejonian (Rock Bench Quarry square), Tiffanian (squares), and Clarkforkian (triangles) land-mammal ages. Solid symbols are fossil localities yielding Wasatchian mammals of the earliest Eocene Wasatchian zones Wa-M and Wa-0 (black diamonds), and Wasatchian zones Wa-1 through Wa-6. Note that the basin axis runs through the area diagonally, from northwest to southeast, and beds on both sides dip toward the basin axis. Dips are shallow north and east of the axis, and steeper south and west of the axis. The drab carbonaceous-shale-rich Fort Union Formation includes Puercan through Clarkforkian fossil mammal localities (Early to Late Paleocene). The red-banded fl uvial Willwood Formation includes Clarkforkian and Wasatchian fossil mammal localities (Late Paleocene and Early Eocene). Qt marks sands and gravels of Quaternary terraces. Terraces near the towns of Powell and Ralston are now irrigated farmland. Individual fossil localities mentioned in the text are labeled (MP-1, SC-2, etc.)
2 A Paleocene epoch different from the Eocene
The geological time scale has gone through a long history, starting with terms that re fl ected time in some sense, but principally the composition, fabric, layering, and coherence of rocks on the earth’s surface: Primary (crystalline), Secondary (layered), Tertiary (consolidated fl uvial sediments), and Quaternary (alluvium). Charles Lyell (1833) subdivided Tertiary time(his ‘Tertiary epoch’) into Eocene, Miocene, and Pliocene intervals. John Phillips (1840) grouped these into the ‘Kainozoic’ or Cenozoic, which we now recognize as an era (Eocene, Miocene, and Pliocene are now ‘epochs’ of the Paleogene and Neogene periods within the Cenozoic Era).
因了一个“怹”,因了满目深情跃然的“怹”,那篇貌似寻常的怀念师长的文章不再寻常,而那个“怹”——常驻于心上的他,一个与“您”同样厚重的字眼,成为牢牢攫住我心的罗盘。
The paleobotanist Wilhelm Schimper (1874) inserted a ‘Période paléocène’ at the beginning of the ‘Époque Tertiaire’ to re fl ect a grade of evolution in European fossil fl oras. Almost immediately, the vertebrate paleontologist Paul Gervais (1877) proposed a new‘Orthrocène’ to re fl ect overlap of European Cretaceous and Eocene vertebrate taxa. Neither proposal was readily accepted. In North America, Edward Drinker Cope (1888) proposed a‘Puerco’ series for strata bearing vertebrate fossils intermediate between and distinct from those of the underlying Cretaceous and overlying Eocene faunas, but this too failed to gain wide acceptance.
生源地为城市的大学生内隐自杀意念得分显著高于生源地为农村的大学生,与之前一些研究者通过外显问卷测量得到的结果相一致[37],原因可能在于与生源地为农村的大学生相比,家住城市的大学生抗挫折能力相对不够,情绪易受外界影响而产生自杀意念.本研究发现,不同家庭类型大学生的自杀意念存在显著差异,普通家庭类型大学生内隐自杀意念得分显著高于多代家庭类型的大学生,原因可能在于多代家庭类型大学生家庭成员较多,可以获得情感上的沟通、交流与支持,因而更加积极乐观.
Bowen G J, Clyde W C, Koch P L et al., 2002. Mammalian dispersal at the Paleocene/Eocene boundary. Science, 295:2062-2065
On July 29 a nice collection of Clarkforkian mammals was made in Sand Coulee, north of SC-188 (Fig. 2), showing again that automobiles made it possible to range widely when collecting. This included the type specimen of ClarkforkianPlesiadapis cookei, named for James W. Cooke. Rock Bench Quarry (Fig. 2) of Torrejonian age was discovered on the east side of Polecat Bench on August 6, when a number of jaws were collected including the type ofPlesiolestes problematicus. Then on August 14 the Mantua Quarry (Fig. 3) of Puercan age was found at a lower stratigraphic level a little south of Rock Bench Quarry. The fi rst fossil discovered at Mantua Quarry was the holotype ofLoxolophus nordicus. The following day several blocks of matrix were removed from Mantua Quarry, and Sinclair, Jepsen, and crew headed back to Princeton.
由于高校教学模式与高中差异较大,学生与教师在课外接触时间少,教育主客体之间的沟通交流时间也少,加之大学生多为“90后”,甚至“00后”,他们的性格特点:敏感、叛逆、羞怯,高校思想政治教育主客体之间的代沟加深,致使思想政治教育的主客体之间的信任度缺乏。依托新媒体平台,有助于转化教育主体的身份,使得教育主体可以以朋友身份与教育客体进行有效沟通,主客体双方的地位平等,学生更容易敞开心扉,消除对思想政治教育主体的畏惧情绪,降低教育客体心理防范意识,从而更加有效引导教育客体在新媒体平台中主动向教育主体敞开心扉,增强了教育主客体之间的信任度,提高了思想政治教育的效果。
A focus on collecting Wa-0 mammals for several summers yielded a representative fauna including: 1) the earliest North American representatives of Artiodactyla, Perissodactyla, and Primates, as Granger (1914) fi rst emphasized; 2) a faunal composition intermediate between the condylarth-dominated faunas of the Clarkforkian and modern-order dominated faunas of the Wasatchian; and 3) conspicuous dwar fi ng of some taxa, such asEctocionin the Wa-0 interval (Fig. 5; Gingerich, 1989, 2003). Further study in the central and southern Bighorn Basin has con fi rmed the distinctiveness of the Wa-0 fauna (Rose et al., 2012; Secord et al., 2012).
高内在动机通过增强认知的灵活性和复杂性提升一般创造力。较强的自我实现需要、浓厚的兴趣、强烈的使命感和责任感可以激发内在动机,一旦内在动机得到激活,人的一般创造力就会大幅度提升。树立远大的理想、信念,培植强烈的社会责任感和使命感,同时保有对某一事物的持久兴趣可以激发较高的内在动机。当人感觉到对工作能够自主支配和有能力时,内在动机最容易产生[8]。给予个体更多的工作自主支配权以及提高个体能力能够促进提高内在动机。最后,由于外在动机对内在动机存在挤出效应[9],因而,在可能的情境下,可以尽量减少对个体的经济报酬、物质奖励等的外在动机激励。
Sinclair and Granger returned to the central Bighorn Basin in 1911, collecting farther south on Fivemile, Tenmile, and Fifteenmile creeks from June 25 through July 29. According to Sinclair’s fi eld book for 1911, they moved north to Cody and the northern Bighorn Basin at the beginning of August. On August 5 they camped at ‘Olsen’s ranch’ near an area of McCullough Peaks badlands. On August 6 Sinclair and Granger collected on the west slopes of McCullough Peaks, fi nding “Very few fossils, none diagnostic.” August 7 was spent climbing the western peak of McCullough Peaks. On the way Sinclair drew a stratigraphic section in a deep draw he estimated to be 1-1/2 miles northeast of the Olsen ranch. The section shows Fort Union Formation sandstones and clays that dip at 15° to the northwest, beneath a terrace of more horizontal volcanic river gravels and overlying limestone blocks. Sinclair also sketched a dry wash, possibly what is now Rough Gulch, along which Fisher (1906) drew the contact of Lower Eocene on Fort Union Formation.
交换模式的计划生育政策原则上应当与市场经济体制相适应,按照交换原理来讲,生育的决定权应由每个家庭自主决定,不过可以在宏观上以原则的方式制定交换模式的计划生育政策来平衡出生率和出生数,但仍会受到其它因素的制约,如经济发展水平、社会保障程度、养育成本、妇女的生育率等。 市场模式的计划生育政策就是要按照市场交换原理,实现实行者和被实行者的利益均衡,以此完善计划生育政策——降低政策成本,实现政策效益。
The fourth phase of biostratigraphic study was focused on Polecat Bench and faunal succession through the Tiffanian land-mammal age (Secord, 2004, 2008). This built on Jepsen’s (1930) initial studies, supplemented with fossils from many new localities, and it led to a revised biochronology and zonation of the Middle and Late Paleocene on Polecat Bench (Fig. 6; Secord et al., 2006). The principal addition to geochronology was dating of sanidine in an ash (Belt ash) at (59.00 ± 0.30) Ma. This ash is in magnetochron C26R, thePlesiadapis churchillilineage zone, and Tiffanian land-mammal biozone Ti-4a. Biostratigraphic innovations and revisions in Secord et al. (2006; Fig. 6) include 1) better correlation of Jepsen’s Cedar Point, Divide, Croc Tooth, Schaff, and Princeton quarries to the geomagnetic polarity time scale, with the first three quarries lying in magnetochron C26R and the latter two quarries lying in magnetochron C25R; 2) better constraint on stratigraphic intervals; 3) better de fi nition of zones and zone recognition by conversion of lineage zones to more discrete interval zones whenever possible (e.g., subdivision of Ti-4 into Ti-4a and Ti-4b based on recognition of the fi rst appearance ofPhenacolemur, subdivision of Ti-5 into Ti-5a and Ti-5b based on recognition of the fi rst appearance ofProbathyopsis, and recognition of Cf-3 based onCopecionrather than the dominance ofPhenacodusandEctocion); and finally 4) better interpolation and correlation of all zones to the geological time scale. Secord (2008) represents the present state of understanding of Paleocene biochronology in the northern Bighorn Basin.
“So-called Ft. Union is mainly blue shales, with a few pink bands... At this horizon we found:Coryphodon,Phenacodus, creodont (?genus), turtle, small broken tooth, and associated jaw fragments. On another bench, somewhat lower down stratigraphically in the same draw are found:Esthonyx-like tooth, phenacodont? tooth, gastropods, bird bones, and a marsupial-like form.”
Surprisingly, there are no entries for these specimens in Sinclair and Granger’s 1911 field catalog. Specimens from the two levels were lumped together when they were listed by Sinclair and Granger (1912; with identi fi cations provided by William D. Matthew of the American Museum of Natural History).
On August 11, Sinclair and Granger moved to a camp north of the Shoshone River and two miles east of the village of Ralston (Fig. 2). From here, on August 12 and 13, they visited the ‘bluff north of Ralston’ (Polecat Bench) and the head of Big Sand Coulee. A larger fauna was obtained, but it again includedPhenacodusandEsthonyxand none of the typical ‘Wasatch’genera. Sinclair and Granger were clear in stating that strata bearing the new fauna lay below the typical Wasatch beds. They were uncertain whether the age difference was signi fi cant, and hence proposed further examination of the stratigraphy (Sinclair and Granger, 1912). This was, nevertheless, a turning point in recognition that Paleocene mammalian faunas are different from those of the Eocene. The 1911 fi eld season ended with Sinclair and Granger working in badlands north and east of McCullough Peaks for a month from August 15 through September 16.
Walter Granger returned to the northern Bighorn Basin in 1912, assisted by William Stein. The two men worked from camps in the Clarks Fork Basin from September 4 through October 10. This provided a much clearer understanding of the faunas and stratigraphy involved, leading Granger (1914) to recognize and name a new ‘Clark Fork’ mammalian fauna from the southwestern base of McCullough Peaks, from Polecat Bench, and from the Clarks Fork Basin itself. He described the Clark Fork fauna as lacking perissodactyls, artiodactyls, rodents, and primates, and as having a predominance ofPhenacodusandEctocioncondylarths, with the latter two genera constituting three-fourths of the collection obtained.Champsosauruswas cited as a distinctly Cretaceous and Paleocene form, and Granger noted that the fauna may possibly represent the “top of the Paleocene series.” When supplemented by later work, this became the basis for a Clarkforkian land-mammal age (Wood et al., 1941).
自古以来,传播在信息传递和交流中创造与维系着社会的发展。传播技术的改变,推动着人类社会的发展,改变着人类缔造的文化世界,改变着个体与社会的精神生活方式。从面对面的口头传播,到印刷的书写传播,再到高科技的电子传播,承载不同传播技术的各种媒体,赋予传播对象不同的权力,对社会伦理变革与个体的道德生活产生着不同的影响。随着信息技术的迅猛发展,新媒体日益凸显出强大的伦理变革功能,成为社会生活领域中全新的权力来源,新媒体赋权塑造了全新的社会生活状态,并影响着传统的伦理世界,具有深刻的伦理意义。
Henry Fairfield Osborn publishedThe Age of Mammals in Europe, Asia, and North Americain 1910. According to Osborn, ‘basal Eocene’ was the fi rst of his ‘seven great faunal phases,’ with no mention of a Paleocene (Osborn, 1910). Sinclair (1912) mentioned the ‘Puerco Paleocene of North America’ but did not elaborate. Then in 1913 William Berryman Scott ofPrinceton, Sinclair’s senior professor, wrote inA History of Land Mammals in the Western Hemisphere(Scott, 1913):
“So far as North America is concerned, the best available evidence points to the conclusion that we should regard the Fort Union, Puerco and Torrejon as the most ancient of the Cenozoic formations, though retaining so many features of Mesozoic life that a separate division of the Tertiary, the Paleocene epoch, is made for them.” (Scott, 1913:99).
Sinclair and Granger (1914) used Paleocene without qualification when summarizing their research on Paleocene deposits of the San Juan Basin, New Mexico.
The transition to acceptance of a Paleocene epoch was completed when Matthew (1914) characterized the Paleocene vertebrate fauna as follows:
“The mammals are dominantly placentals of archaic orders... Approximately 10 per cent of the fauna is multituberculate. The remainder belong to groups of placentals which became extinct during the Eocene. The later Tertiary and modern orders of mammals are not present except the Carnivora and certain groups doubtfully referred to Insectivora and Edentates. There are no perissodactyls, artiodactyls, rodents, or primates, these orders appearing suddenly at the beginning of the true Eocene.” (Matthew, 1914:382).
Acceptance of a Paleocene epoch distinct from the Eocene was due in part to continued, if inconsistent, use of Paleocene by Europeans (e.g., Depéret, 1906; Dollo, 1909), but more importantly it was due to Sinclair and Granger’s demonstration of a thick succession of strata in the northern Bighorn Basin with a fauna lacking Perissodactyls, Artiodactyls, Rodents, or Primates. Matthew’s (1914) noting “no perissodactyls, artiodactyls, rodents, or primates”copies verbatim the conclusion of Sinclair and Granger’s 1911 and 1912 field work in the northern Bighorn Basin. Perissodacyls, artiodactyls, and true primates were absent from Granger’s Clark Fork collection, but further investigation would show that rodents are present and important in this interval.
现代观赏石的审美理念突破了古典观赏石的局限性,其审美标准被界定为:形、质、色、纹、韵及命名、配座、传承八大要素,极大地扩展了观赏石的现代审美范畴,适应了人们对观赏石文化的多元化需求,在新的观赏石标准里,色彩成了重要的审美元素。
3 Succession of Paleocene mammalian faunas
3.1 Princeton fi eld research
The next advance in Paleocene biostratigraphy of the Bighorn Basin came in 1928 when William Sinclair’s student, Glenn Jepsen (Fig. 1C), decided to explore Paleocene strata of the northern Bighorn Basin, which became his doctoral dissertation research (Jepsen, 1930). Sinclair led an expedition to the Dorsey Creek badlands of the central Bighorn Basin in 1927, with Jepsen as a participant. The motivation for Jepsen’s subsequent investigation of the Paleocene was never stated explicitly, but Sinclair may have been interested to see work he had started on Paleocene faunas continue. Jepsen himself may have been intrigued by the discovery of Paleocene primates (Proprimates) in the Fort Union Formation of Montana (Gidley, 1923), and by a 1927 news report of a human tooth found in the ‘Garden of Eden,’ in the Fort Union Formation of the Clarks Fork Basin at Bear Creek, Montana (Simpson, 1928). The ‘human’tooth proved to be a molar of the condylarthPhenacodus.
Jepsen’s 1928 field research started on June 27 when he, Lemoyne Cook, and Joseph F. Page set up camp in the Clarks Fork Basin. Their fi rst camp was at locality SC-2 (Fig. 2),which is richly fossiliferous, yielding typical early Wasatchian fossils. Thus it was a good place to train new collectors. Jepsen’s crew camped at SC-2 until July 13, but they had an automobile and so could range north and east from this camp to explore Clarkforkian Late Paleocene as well as Wasatchian Early Eocene strata.
On July 13 Jepsen moved to a second camp near locality SC-80 (Fig. 2) on the west side of Polecat Bench. From this camp the crew worked along Polecat Bench and into the head of Big Sand Coulee. During this time the holotype ofProbathyopsis successorwas found at a locality in the head of Big Sand Coulee, in red-banded strata interpreted to be Wasatchian because of their red coloration (but now known to be Clarkforkian in age). Then on July 27 Jepsen moved to a third camp near an abandoned coal mine just east of localities SC-262, 263, and 264 (Fig. 2) on the southeast side of Polecat Bench. During the week spent at this third camp dinosaur remains were found in the Cretaceous to the north and east, a specimen ofPlesiadapsis rexwas found near the camp, and the holotype ofTitanoides gidleyiwas found a mile southwest of the camp. Jepsen’s expedition of 1928 proved the feasibility of fi nding fossils stratigraphically below the typical ‘Clark Fork’ documented by Granger (1914).
推荐理由:全书以中国历史时序排列,分为史前、夏商西周、春秋战国、秦汉、魏晋南北朝、隋唐五代、辽、宋、西夏、金、元、明、清(1644~1840)、清(1840~1911)、中华民国共15卷(25册),1700万字。全书各卷均以综述与治乱兴衰、经济、国家控制、社会结构、精神文化、社会生活等六编构成,以专题形式叙史,后附主要参考史料、历史纪元表。
As a result of this success, in 1929 Sinclair returned to the Clarks Fork Basin with Jepsen and the undergraduate students James W. Cooke, Sven Dorf, and Joseph F. Page. They set up camp on June 29 near locality SC-198 (Fig. 2) on the west edge of Polecat Bench, at a site Jepsen had visited during his last day in the fi eld the year before. The fi rst two weeks of the 1929 season were spent prospecting near this fi rst camp, mostly along the west edge of Polecat Bench. On July 11 the fi rst specimens were collected at what later became the very productive Princeton Quarry of late Tiffanian age. However, the actual concentration of jaws and teeth at Princeton Quarry was not recognized until July 22, when two days were spent quarrying here.Phenacolemur pageifrom Princeton Quarry was named for Joseph F. Page.
William Sinclair of Princeton University (Fig. 1A) and Walter Granger of the American Museum of Natural History (Fig. 1B) initiated fi eld work in continental strata of the Bighorn Basin in 1910. Their objective was to understand how earlier observations on Eocene faunas of the Wind River Basin might relate to those in the Bighorn Basin. The latter was known to be large, richly fossiliferous, and poorly explored. Eocene strata in both basins include alternations of bright red and drab blue clays. Sinclair and Granger (1912) were among the fi rst to show that iron concentrations differed in the two lithologies, and they were possibly the fi rst to recognize that the color banding re fl ected cycles of climate change.
联合国维和任务基本上都在战略距离较远的国外开展,一般来说条件比较恶劣,但这些年来一直处于和平环境下的中国军队实战经验不够丰富,远离本土作战的能力较为欠缺,在纷繁复杂的国际维和环境中发挥作用的能力受到限制。此外,空中和海上补给能力不足也阻碍了中国远距离迅速部署部队的能力。
In six weeks of fi eld work Sinclair and Jepsen succeeded in fi nding what became three very productive Puercan, Torrejonian, and Tiffanian-age fossil quarries, intercalated between Cretaceous strata below and Clarkforkian latest Paleocene strata above. The 1928 and 1929 collections formed the basis of Jepsen’s doctoral dissertation (Jepsen, 1930) and initiated a major publication on multituberculates (Jepsen, 1940). Jepsen continued fi eld research in the northern Bighorn Basin for the rest of his professional career.

Fig. 3 Mantua Quarry (Puercan land-mammal age, Early Paleocene) on the east side of Polecat Bench (Fig. 2), discovered by Glenn Jepsen’s Princeton University team at the end of the 1929 fi eld season This locality is a dif fi cult but productive fossil locality yielding the Puercan or ‘Mantuan’-age mammalian fossils described by Jepsen (1930) and later Van Valen (1978). A. regional view of Mantua Quarry in westdipping strata at Polecat Dome on the east side of Polecat Bench; B. close-up view Mantua Quarry. Note the scree slope below the quarry of sandstone blasted from the cliff face above the quarry. Fossils were found in the base of the thickest sandstone, just above the lignite marking the conformable Cretaceous-Paleocene contact. Photographs by the author
3.2 Analysis of Princeton Collections
The Princeton University collections of Paleocene mammals from the northern Bighorn Basin made by Glenn Jepsen during the course of his career were at once both important and understudied. I mentioned the publication on multituberculates (Jepsen, 1940), Jepsen’s last substantial publication on Paleocene mammals, but he continued collecting until 1974. Elwyn Simons (1960) published a monographic treatment of Paleocene Pantodonta based onthe Princeton collection, and Michael Archer (1967) wrote a senior thesis at Princeton on the Paleocene Dinocerata.
Roger Wood (1967) published an analysis of American Museum and Princeton collections in which he concluded:
“Therefore, the Clark Fork ‘fauna’ consists of four specimens referable to four mammalian genera, each from a different locality. None of the genera (and not more than one of the species) are restricted to a lithically distinguishable Clark Fork horizon. Such evidence scarcely warrants recognition of the Clark Fork as a provincial age, faunal zone, or member of the Polecat Bench Formation.” (Wood, 1967:28).
The Wood study annoyed Jepsen, and part of the time I spent working for Jepsen during the summers of 1967 and 1968 was spent trying to fi nd Clarkforkian-age sites to refute Wood’s conclusions. Later our University of Michigan research focused on this question as well (see below).
Jepsen and Woodburne (1969) described a specimen of a ‘hyracothere’ or horse that they believed came from Paleocene ‘horse pasture’ strata at locality SC-144 on the west side of Polecat Bench (Fig. 2), a locality now known to be Tiffanian in age. The specimen representsHyracotherium, probablyH. grangeri, but it was not recognized as a Paleocene horse when it was collected, and the locality assigned to the specimen (or specimen assigned to the locality) later in the laboratory is almost certainly incorrect. The Jepsen-Woodburne study followed publication of another claimed PaleoceneHyracotherium(Morris, 1968), which also proved to be Early Eocene rather than Paleocene in age (Novacek et al., 1991).
My senior thesis at Princeton was a study of Polecat Bench stratigraphy and a succession of pollen fl oras (Gingerich, 1968). The Polecat Bench stratigraphic section later formed the backbone of my Ph.D. dissertation on the biostratigraphy and evolution of the proprimate family Plesiadapidae (Gingerich, 1974, 1975, 1976). Kenneth Rose published a major review of Paleocene proprimates of the family Carpolestidae based largely on Princeton specimens (Rose, 1975, 1977). Combining the Rose study of carpolestids with my results for plesiadapids yielded the robust pattern of diversi fi cation and change through time shown in Fig. 4. Given the size of the Princeton samples available to us in 1974-1976, it is no surprise that our initial hypotheses in relation to stratigraphy have stood the test of time as additional specimens were added.
A large number of studies of Paleocene mammals based on the Princeton collection from the vicinity of Polecat Bench appeared after Jepsen retired. These included Rose (1973, 1975, 1979, 1981), Szalay (1973), Gingerich (1974, 1975, 1976, 1977, 1979, 1980, 1983a), Van Valen (1978), Krause (1980, 1982, 1987a, b), Rose and Krause (1982), Thewissen and Gingerich (1987), Gunnell (1986, 1989), Thewissen (1989, 1990), Zhou (1995), and Bloch (2001). The northern Bighorn Basin is centrally located in North America, and has the thickest and most complete Middle and Late Paleocene stratigraphic sections. However, important studies of Paleocene mammals have been carried out in several other basins ranging geographically from Alberta, Canada, in the north, to Texas, U.S.A., in the south. The additional studies are reviewed in Archibald et al. (1987) and in Lofgren et al. (2004).
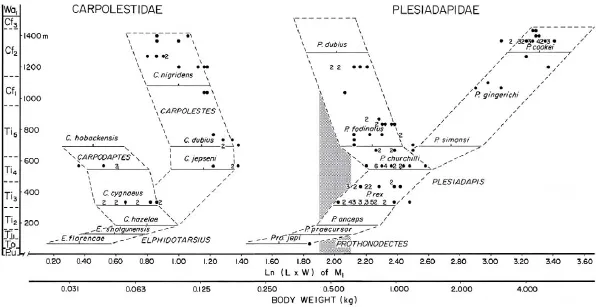
Fig. 4 Stratigraphic plot of evolutionary change in the size of fi rst lower molars of Middle and Late Paleocene Carpolestidae and Plesiadapidae from strata bordering Polecat Bench in the northern Bighorn Basin, Wyoming Initial hypotheses of size change in carpolestids and plesiadapids are shown as horizontal bars connected by dashed lines, based on tooth sizes for species from Princeton University localities reported by Rose (1975) and Gingerich (1976). University of Michigan specimens collected subsequently from the same strata generally fall within the ranges found initially, illustrating the testability of stratophenetic hypotheses of phylogenetic relationships and evolutionary change. Body weight inferred from tooth size is plotted on the abscissa, and time inferred from superposition and faunal succession is plotted on the ordinate. Carpolestids are smaller than 500 g in body weight (shaded vertical bar), whereas plesiadapids are generally larger than 500 g. Abbreviations of land-mammal ages: Cf. Clarkforkian; Pu. Puercan; Ti. Tiffanian; To. Torrejonian; Wa. Wasatchian. Figure reprinted from Gingerich (1987)
3.3 University of Michigan fi eld research
University of Michigan fi eld research in the northern Bighorn Basin was initiated in 1975 with two broad objectives. The fi rst was to study evolutionary change, bed by bed, in a broad range of Paleocene-Eocene mammals (following the examples of Gingerich, 1974, 1975, 1976, and Rose, 1975). The second objective was to clarify the transition from Paleocene to Eocene mammals, both in terms of the land mammal ages involved and in terms of the appearance of modern groups such as Artiodactyla, Perissodactyla, and Primates. Taxonomic revisions were required for many groups, which were facilitated by the collections available from earlier American Museum and Princeton University expeditions. However, our new and more ambitious objectives required additional collections from more tightly constrained and better documented stratigraphic intervals. This required a continuous program of fi eld exploration, which proceeded in four successive phases.
The fi rst biostratigraphic phase involving new fi eld work was an extension of Granger’s (1914) Clark Fork mammalian fauna. Horace Wood et al. (1941) listed the type locality for the Clarkforkian land-mammal age as the “divide between Bighorn and Clarks Fork basins” and cited Jepsen’sPlesiadapis cookeias the index fossil. The Clarkforkian was said to include fi rstappearances ofCoryphodonandEsthonyx, and last appearance ofCarpolestesandPlesiadapis.
Roger Wood (1967) had written, concerning the index fossilPlesiadapis cookei:
“Precise locality data are associated with only one of the supposedly Clarkforkian representatives ofPlesiadapis cookei. But this specimen, the type, was found in the region east of the mouth of Little Sand Coulee where disagreements exist concerning the location of the contact between Paleocene and Eocene sediments. The specimen’s uncertain stratigraphic derivation precludes its being clearly assigned either a Paleocene or an Eocene age. The two other specimens ofP. cookeiwere also found in the same general area, and the same uncertainty applies to them. Thus, on the basis of published evidence it is not possible to includeP. cookeide fi nitely in the Clark Fork fauna.” (Wood, 1967:23).
However, my subsequent dissertation research (Gingerich, 1974, 1976) showed thatPlesiadapis cookeioccupied a distinct zone crossing the Clarks Fork Basin just to the south of the Fort Union-Willwood formational boundary (Fig. 2).
With this as background, we collected fossils back and forth across the formational boundary from 1975 through 1978, and these new collections became the basis for full documentation and systematic analysis of what became a large Clarkforkian land-mammal fauna (Rose, 1979, 1981). In the course of this fi eld work, we were able to identify a ‘boundary sandstone’ unit extending across the Clarks Fork Basin separating Clarkforkian Late Paleocene mammals below from Wasatchian Early Eocene mammals above. Much of the Clarkforkian sequence of strata in the Clarks Fork Basin contains red beds, which misled Wood (1967; and also Jepsen) into thinking that the beds were Eocene. A paleomagnetic study along the south side of Polecat Bench and north side of Clarks Fork Divide enabled correlation of the Paleocene section on Polecat Bench to the geomagnetic polarity timescale (Butler et al., 1981).
Once the Clarkforkian was thoroughly analyzed, our second phase of biostratigraphic study involved the Clarkforkian-Wasatchian boundary. The ‘boundary sandstone’ separating the two land-mammal ages was found to contain a Wasatchian mammalian fauna distinct from those of the underlying Clarkforkian and overlying Wasatchian. For convenience I had numbered Rose’s (1981) three Clarkforkian faunal zones Cf-1, Cf-2, and Cf-3, and I had divided Granger’s ‘Sand Coulee’ beds into two Wasatchian faunal zones numbered Wa-1 and Wa-2 (with additional zones below and above these; Gingerich, 1983b). The only way to insert a stratigraphically intermediate Wasatchian zone in the sequence was to number it Wa-0.
Sinclair and Granger spent the summer of 1910 in the central Bighorn Basin, collecting near Otto and St. Joe, on Dorsey Creek and Elk Creek. They worked from July 10 through September 27, and were able to find three productive ‘Wasatch’ intervals that later became lower, middle, and upper Graybullian faunas. Sinclair and Granger found a productive ‘Wind River Lost Cabin’ interval above these, and inferred the presence of intervening ‘Wind River Lysite’ beds (Sinclair and Granger, 1911).
As fi eld investigation of the Paleocene-Eocene transition continued, it became clear that there was a new and distinctive mammalian fauna with Wasatchian af fi nities in a 3-5 meter interval between zones Cf-3 and Wa-0. This was unusual in yielding an Early Eocene genus,Meniscotherium, that is otherwise unknown in the Bighorn Basin. Consequently the zone between Cf-3 and Wa-0 was called the Bighorn BasinMeniscotheriumzone, and abbreviated Wa-M (Gingerich and Smith, 2006).

Fig. 5 Change in the tooth size and estimated body weight of a lineage of the condylarth genusEctocionthrough Late Paleocene and Early Eocene time Sample size is 452 individual specimens. Each fi lled circle represents fi rst lower molar size in a different individual. Specimens included come from 78 successive stratigraphic levels spanning the Clarkforkian through middle Wasatchian land-mammal ages in the Clarks Fork Basin. Shaded interval tracks mean values, with a 1:N:1 sample size weighting of successive means and a shaded width of ±2 phenotypic standard deviations. Successive means separatingEctocion parvusfrom earlier and laterEctocion osbornianusare conspicuously greater than those separating successive means withinE. osbornianus(inset box A). Rates of evolutionary change between successive samples are calculated on the assumption of a 2-year generation time (inset box B). Figure modi fi ed from Gingerich (2006)
The second phase of biostratigraphic study continues today with attempts to put Paleocene-Eocene faunal change in paleoenvironmental context. Following recognition of Wa-0 as a distinct faunal zone in 1989, both soil nodules and mammalian teeth from this interval were found to have unusually light isotopic values (Koch et al., 1992). This carbon isotope excursion (CIE) was seen to correlate with a similar CIE found at or near the Paleocene-Eocene boundary in Ocean Drilling Program site 690 at 65° south latitude in the Weddell Sea off Antarctica (Stott et al., 1990; Kennett and Stott, 1991). Correlation of the two CIEs was important for three reasons: 1) it linked the continental Paleocene-Eocene boundary to the marine Paleocene-Eocene boundary in a way not possible before (the marine boundary was often placed a million years later in time than the continental Paleocene-Eoceneboundary); 2) the CIE at site 690 was associated with an oxygen isotope excursion indicating oceanic warming of 4-5°C; and 3) similar carbon isotopic excursions at the Paleocene-Eocene boundary in the northern and southern hemispheres, on land and in the sea, suggested that climate change at this time was global (Koch et al., 1992). Further studies at Polecat Bench include those by Clyde and Gingerich, 1998; Bowen et al., 2001, 2014; Magioncalda et al., 2004; Abdul Aziz et al., 2008; and Clyde et al., 2013.
The third phase of biostratigraphic study in the northern Bighorn Basin was a major effort to document Wasatchian faunas in and around the McCullough Peaks (Clyde, 1997, 2001). As background, Clyde et al. (1994) were able to extend correlation of Wasatchian biochrons to the geomagnetic polarity timescale. Research on the systematics of Paleocene-Eocene mammals continued in parallel with other studies, with reviews of taxa such as Perissodactyla (Gingerich, 1991), Mesonychia (Zhou, 1995), and small mammals preserved in freshwater limestones (Bloch, 2001).
August 8 and 9 were spent collecting fossils in the ‘so-called’ Fort Union Formation on the southwest side of McCullough Peaks (locality MP-1 in Fig. 2). Sinclair and Granger (1912) published a photograph of the site, which is on the divide between two dry creeks that are now called Rough Gulch and Penny Gulch. Sinclair described the site and the fossils recovered in his 1911 fi eld notebook:
4 Parallel research in China
Minchen Chow studied in the United States from 1947 until 1950, when he received his Ph.D. degree from Lehigh University in Bethlehem, Pennsylvania (Miao et al., 2010). Chow’s doctoral dissertation was on Paleozoic freshwater bivalves of Pennsylvania. Lehigh has a long history of education of Chinese students that began in 1879. More than two thousand Chinese graduate and undergraduate students have been educated at Lehigh, and historically the relationship has been especially close in the fields of geology and mining. This can be attributed to Bethlehem’s early development as a center of heavy industry and trade. Bethlehemwas once the second-largest steel producing center in the United States (second to Pittsburgh). Steel requires iron and coal, which depend in turn on expertise in geology and mining.

Fig. 6 Paleocene biozones and faunal evolution through the Early, Middle, and Late Paleocene bordering and near Polecat Bench in the northern Bighorn Basin, Wyoming The North American Torrejonian (Torr) land-mammal age (LMA) represents the latter part of the Early Paleocene Danian stage/age, the Tiffanian represents much of the Middle Paleocene Selandian and Late Paleocene Thanetian stage/age, and the Clarkforkian (Clark) represents the latter part of the Thanetian stage/age. The chart shows two zones of the Torrejonian (To-2 and To-3), eight zones of the Tiffanian (Ti-1 through Ti-6), and three zones of the Clarkforkian (Cf-1 through Cf-3). Each zone is also named. Some are interval zones (iz), while others are lineage zones (lz) representing successive stages in the evolution ofPlesiadapis. Representative localities are listed for each zone. Magnetic polarity chrons are shown near the right side of the chart. Biozones are interpolated to fi t a numerical calibration in millions of years (Ma), based on radiometric dating of ash beds and magnetochrons. Inset figure to the right of the chart shows decreasing mammalian species richness (diversity) through the Early and Middle Paleocene when deep ocean temperatures (and by inference ocean and land surface temperatures) were declining, and increasing species richness through the Middle and Late Paleocene when deep ocean temperatures were rising. All mammalian faunal diversity estimates are rarified to a common sample size of 105 specimens. Three major immigrations of allochthonous taxa are recorded at the beginning of Tiffanian biochron Ti-5, at the beginning of the Clarkforkian, and at the beginning of the Wasatchian, with the latter being divisible into distinct Wa-M and Wa-0 immigrations. The first two immigrations include taxa that are distinctly Asian in af fi nities, and the third includes taxa of both European and Asian af fi nities. All three immigrations into North America happened during the interval of Middle to Late Paleocene warming and increasing species diversity. Research and illustrations are from Secord (2008)
While at Lehigh, Chow took summer courses at the American Museum of Natural History in New York. There in 1949 he met the esteemed vertebrate paleontologists of the time: George Gaylord Simpson and Ned Colbert. Chow almost certainly reviewed the Paleocene mammals of Mongolia, which Simpson had helped to describe. Miao et al. (2010) reported that this is where Chow decided he wanted to study vertebrate paleontology. A young Bobb Schaeffer of the American Museum was studying Triassic fi shes excavated on the Princeton University campus. Schaeffer recommended that Chow approach Glenn Jepsen at Princeton for training, and he no doubt also recommended Chow to Jepsen. Chow moved to Princeton later in 1949. He then spent part of a long summer in 1950 working with graduate student William Morris in the Early Eocene of the Washakie Basin in southern Wyoming, and part of the summer of 1950 prospecting with Jepsen’s crew in the Paleocene of the northern Bighorn Basin.
卓时Premier系列Precious月相功能腕表的镶钻表耳重现海瑞温斯顿纽约第五大道旗舰店的拱门造型,令人过目难忘,表圈与表耳共镶嵌57颗圆形明亮式切工钻石。 时、分指示及月相功能通过优质瑞士自动上链机械机心驱动,其动力储存长达68小时。腕表配备蓝色或棕色鳄鱼皮表带,搭配镶嵌17颗圆形明亮式切工钻石的白18K金或玫瑰18K金材质针扣。
Chow received his Ph.D. from Lehigh in December of 1950, and returned to China at the beginning of 1951. He joined C. C. Young at the newly established Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) in Beijing in 1952. Chow was a co-founder of the journalVertebrata PalAsiatica, fi rst published in 1957, and he edited the journal from 1980 to 1991. Chow directed the IVPP from 1979 to 1984, and he directed the Beijing Natural History Museum from 1982 until his death in 1996. The time spent in the Bighorn Basin made a lasting impression on Minchen Chow, and in 2001, at his request, his ashes were interred near Princeton Quarry in the Clarks Fork Basin.
The time Minchen Chow spent working on Paleocene mammals at Princeton and in the field in Wyoming had a lasting influence on Chinese paleontology too. The only Paleocene mammals known from Asia came from Gashato (Matthew and Granger, 1925; Matthew et al., 1928, 1929; Szalay and McKenna, 1971) and from Nemegt (Trofimov, 1952; Flerov, 1952; Gromova, 1952), both in Mongolia. The first Paleocene mammal to be found in China was found in 1959 by a geologist working in the Turfan Basin of far western Sinkiang (Xinjiang) Autonomous Region. Chow (1960) recognized the teeth of this to represent a dinoceratan or uintathere, and named a new speciesProdinoceras turfanensisthat was slightly smaller than its congenerProdinoceras martyrnamed by Matthew et al. (1929) from Gashato. Chow almost certainly studied the type ofP. martyrwhen he was a student at the American Museum.
In 1960, geological mapping in the Nanhsiung or Nanxiong Basin of Kwangtung or Guangdong Province in South China produced vertebrate remains, and this fortunate discovery inspired a focused search for more. By 1963 Young and Chow were able to report the first Paleocene mammals from the Nanxiong Basin in Guangdong. These were compared to the Torrejonian-age generaPantolambda(Pantodonta) andDissacus(Mesonychia), which Chow also knew from his studies at Princeton and the American Museum. Young and Chow (1963) wrote:
“Probably more interesting and significant is the finding of some primitive mammals in the upper part ... of the series. So far as those have been preliminarily identi fi ed, at least a creodont and a pantodont ... are present. The pantodont, which is represented by skulls and skeletons, is apantolambdid-like form of small size with rather primitive dentition. The creodont, represented by skull fragments and upper cheek teeth, resembles the Torrejonian mesonychidDissacus. The fi nding of these forms indicates that we have most likely located in North Kwangtung a promising earlier Paleocene mammalian horizon in Asia.” (Young and Chow, 1963:1411).
They are also my friends.They are from England.What should I give them?Why?
Research on the Paleocene mammals of China was interrupted in its infancy in 1966 by initiation of the Chinese Cultural Revolution. This revolution lasted through the early 1970s.Vertebrata PalAsiaticaceased publication at volume 10 in 1966, and publication did not resume until volume 11 appeared in 1973. The first issue of volume 11 included “New mammalian genera and species from the Paleocene of Nanhsiung, N. Kwangtung” by Minchen Chow, Yuping Chang, Banyue Wang, and Suyin Ting (Chow et al., 1973).
The search for South China vertebrate fossils fi rst found in the Nanxiong Basin expanded to the Qianshan Basin of Anhui Province in 1970 and 1971 (Hou, 1974), the Xuancheng Basin in Anhui Province (Tang and Yan, 1976), the Chaling Basin in Hunan Province (Gao, 1975), and the Chijiang Basin of Jiangxi Province (Chow, 1977; Ting and Zhang, 1979). Paleocene mammals were also found at Nomogen in Nei Mongol (Inner Mongolia) (Chow et al., 1976). These discoveries all came so rapidly that by 1977 Chow could write (Chow, 1977):
“The recovery of Paleocene mammalian fossils ... is of considerable interest ... to paleomammalogists all over the world. These Paleocene beds and faunas are the oldest and one of the most complete sequences of continental mammal-bearing Paleocene yet known anywhere in the world outside of North America. They are very important to a correct understanding of the evolution and distribution of mammals at the beginning of the Age of Mammalia.” (Chow, 1977:666).
Chow’s (1977) summary published in English did interest paleomammalogists all over the world, because he continued and concluded:
“The most characteristic feature of the Paleocene mammalian faunas of Asia is the greater proportion of the autochthonous and endemic forms in almost every faunal assemblage. ... almost none of the genera so far known from the Paleocene of Asia can be assigned with certainty to any known North American genera, less so to those of Europe. ... These features clearly show that Asia was then zoogeographically quite isolated from both Europe and North America.“The most certain and radical change in the mammalian faunas of Asia occurred at the beginning of the Eocene, when Beringia became an effective corridor for interchange of land animals between Asia and North America. ... For lack of information, it is still quite dif fi cult to make any plausible inference of sound speculation about the paleozoogeographical relationship between Asia and Europe (mainly western Europe) or between Asia and the subcontinent of India.” (Chow, 1977:677).
What a remarkable institutional accomplishment it was for teams from the Institute of Vertebrate Paleontology and Paleoanthropology to discover and document that the Paleocene of the largest northern continent was fi lled with animals different from those of Europe and North America. What a challenge to all of us to explain a history that rapidly became much more complicated and interesting.
The 1977 report was only a beginning. Research on Chinese faunas has continued, with many new discoveries and many new publications (e.g., Chow, 1979; Li and Ting, 1983;Russell and Zhai, 1987; Tong et al., 1995; Meng and McKenna, 1998; Meng et al., 1998; Beard, 1998; Ting, 1998; Wang et al., 1998, 2007, 2010; Bowen et al., 2002, 2005).
5 Conclusions
Chance favors the prepared mind, which is another way of saying that we build on the accomplishments of those who precede us. William Sinclair and Walter Granger’s exploration of the McCullough Peaks and Clarks Fork Divide showed that there was a substantial interval of Cenozoic time preceding the Eocene that lacks mammals of modern orders and is worthy of recognition as an epoch of equal rank. Glenn Jepsen found all of the North American Paleocene land-mammal ages in one sequence on Polecat Bench, with the Cretaceous below and Eocene above. But when the Polecat Bench record seemed to be the history of life in the Paleocene, a student he trained, Minchen Chow, led teams that discovered Asia’s history was different. We are fortunate to have a rich fossil record of Paleocene mammals to work with on all of the northern continents, but there is still more work to do to understand the full history on each continent, and it continues to be an ongoing challenge to understand how the individual histories fi t together.
Acknowledgments I am indebted, as we all are, to William Sinclair, Walter Granger, Glenn Jepsen, and Minchen Chow for their efforts to develop the parallel histories of life in North America and in Asia. I will always be grateful to Minchen Chow for the invitation to spend several weeks at the Institute of Vertebrate Paleontology and Paleoanthropology in 1979, when I was permitted to mold and distribute casts of many of the new specimens studied and described by IVPP colleagues in the 1970s. I thank Yuanqing Wang, Chendong Wang, Yi Zhang, Qian Li, and Guosheng Yu for organizing the 2015 International Symposium on Terrestrial Paleocene Biota and Paleobiogeography in Qianshan, with fi eld trips to Qianshan Basin Paleocene sites and Tianzhushan Global Geopark, and I thank Yuanqing Wang for inviting me to participate. Jin Meng, Desui Miao, and Yuanqing Wang provided helpful reviews of the manuscript.
Abdul Aziz H, Hilgen F J, Luijk Gerson M van et al., 2008. Astronomical climate control on paleosol stacking patterns in the Upper Paleocene-Lower Eocene Willwood Formation (Bighorn Basin, Wyoming). Geology, 36: 531-534
Archer M, 1967. A revision of the taxonomy and phylogeny of the early Tertiary unitatheres. A. B. thesis. Princeton:Princeton University. 1-128
Archibald J D, Clemens W A, Gingerich P D et al., 1987. First North American land mammal ages of the Cenozoic era. In: Woodburne M O ed. Cenozoic Mammals of North America: Geochronology and Biostratigraphy. Berkeley:University of California Press. 24-76
二是坚持示范引路抓点带面打开工作新局面。镇街人大工作存在的困难和不足,既有历史原因,也有基层工作实情。鉴于目前现状,示范引路抓点带面是现实路径,应根据不同区位,选取试点单位先行开展。
Beard K C, 1998. East of Eden: Asia as an important center of taxonomic origination in mammalian evolution. In: Beard K C, Dawson M R eds. Dawn of the Age of Mammals in Asia. Bull Carnegie Mus Nat Hist, 34: 5-39
Bloch J I, 2001. Mammalian paleontology of freshwater limestones from the Paleocene-Eocene of the Clarks Fork Basin, Wyoming. Ph.D. thesis. Ann Arbor: University of Michigan. 1-358
Bowen G J, Koch P L, Gingerich P D et al., 2001. Re fi ned isotope stratigraphy across the continental Paleocene-Eocene boundary on Polecat Bench in the northern Bighorn Basin. In: Gingerich P D ed. Paleocene-Eocene Stratigraphy and Biotic Change in the Bighorn and Clarks Fork Basins, Wyoming. Univ Mich Pap Paleont, 33: 73-88
2.1 Princeton and American Museum fi eld research
2.2 Acceptance of a Paleocene epoch
Bowen G J, Koch P L, Meng J et al., 2005. Age and correlation of fossiliferous Late Paleocene-Early Eocene strata of the Erlian Basin, Inner Mongolia, China. Am Mus Novit, 3474: 1-26
Bowen G J, Maibauer B J, Kraus M J et al., 2014. Two massive, rapid releases of carbon during the onset of the Paleocene-Eocene thermal maximum. Nat Geosci, 8: 44-47
Butler R F, Gingerich P D, Lindsay E H, 1981. Magnetic polarity stratigraphy and biostratigraphy of Paleocene and Lower Eocene continental deposits, Clarks Fork Basin, Wyoming. J Geol, 89: 299-316
Chow M C, 1960.Prodinocerasand a summary of mammalian fossils of Sinkiang. Vert PalAsiat, 4: 99-102
Chow M C, 1977. Palaeocene vertebrate horizons and mammalian faunas of South China. Sci Sin, 20: 665-678
Chow M C, 1979. Vertebrate paleontology in China (1949-1979). Vert PalAsiat, 17: 263-276
Chow M C, Chang Y P, Wang B Y et al., 1973. New mammalian genera and species from the Paleocene of Nanhsiung, N. Kwangtung. Vert PalAsiat, 11: 31-35
Chow M C, Qi T, Li Y, 1976. Paleocene stratigraphy and faunal characters of mammalian fossils of Nomogen Commune, Si-Zi-Wangqi, Nei Mongol. Vert PalAsiat, 14: 228-233
Clyde W C, 1997. Stratigraphy and mammalian paleontology of the McCullough Peaks, northern Bighorn Basin, Wyoming:implications for biochronology, basin development, and community reorganization across the Paleocene-Eocene boundary. Ph.D. thesis. Ann Arbor: University of Michigan. 1-270
Clyde W C, 2001. Mammalian biostratigraphy of the McCullough Peaks area in the northern Bighorn Basin. In: Gingerich P D ed. Paleocene-Eocene Stratigraphy and Biotic Change in the Bighorn and Clarks Fork Basins, Wyoming. Univ Mich Pap Paleont, 33: 109-126
Clyde W C, Gingerich P D, 1998. Mammalian community response to the latest Paleocene thermal maximum: an isotaphonomic study in the northern Bighorn Basin, Wyoming. Geology, 26: 1011-1014
Clyde W C, Stamatakos J, Gingerich P D, 1994. Chronology of the Wasatchian land-mammal age (Early Eocene):magnetostratigraphic results from the McCullough Peaks section, northern Bighorn Basin, Wyoming. J Geol, 102:367-377
Clyde W C, Gingerich P D, Wing S L et al., 2013. Bighorn Basin Coring Project (BBCP): a continental perspective on early Paleogene hyperthermals. Sci Dril, 16: 21-31
Clyde W C, Wilf P, Iglesias A et al., 2014. New age constraints for the Salamanca Formation and lower Río Chico Group in the western San Jorge Basin, Patagonia, Argentina: implications for Cretaceous-Paleogene extinction recovery and land mammal age correlations. Geol Soc Am Bull, 126: 289-306
Cope E D, 1888. Synopsis of the vertebrate fauna of the Puerco series. Trans Am Philos Soc, 16: 298-361
Depéret C, 1906. Relations stratigraphiques des faunes de Cernay et de Meudon au mont de Berru. Bull Soc Géol Fr, Sér 4, 6:442-443
Dollo L, 1909. The fossil vertebrates of Belgium. Ann New York Acad Sci, 19: 99-119
Fisher C A, 1906. Geology and water resources of the Bighorn Basin, Wyoming. US Geol Surv Prof Pap, 53: 1-72
Flerov K K, 1952. Pantodonts (Pantodonta) collected by the Mongolian paleontological expedition of the Academy of Sciences of the U.S.S.R. Tr Paleont Inst Akad Nauk SSSR, 41: 43-50
Gao H, 1975. Paleocene mammal-bearing beds of Chaling Basin, Hunan. Vert PalAsiat, 13: 89-95
Gervais P, 1877. Enumération de quelques ossements d'animaux vertébrés recueillis aux environs de Reims par M. Lemoine. J Zool, Paris, 6: 74-79
Gheerbrant E, 1992. Les mammifères Paléocènes du Bassin d'Ouarzazate (Maroc) I. Introduction générale et Palaeoryctidae. Palaeontogr, Abt A, 224: 67-132
Gheerbrant E, Rage J-C, 2006. Paleobiogeography of Africa: how distinct from Gondwana and Laurasia? Palaeogeogr Palaeoclimat Palaeoecol, 241: 224-246
Gidley J W, 1923. Paleocene primates of the Fort Union, with discussion of relationships of Eocene primates. Proc US Natl Mus, 63: 1-38
Gingerich P D, 1968. Pollen stratigraphy of the Polecat Bench Formation, Paleocene, Park County, Wyoming. A. B. thesis. Princeton: Princeton University. 1-81
Gingerich P D. 1974. Cranial anatomy and evolution of early Tertiary Plesiadapidae (Mammalia, Primates). Ph.D. thesis. New Haven: Yale University. 1-370
Gingerich P D, 1975. New North American Plesiadapidae (Mammalia, Primates) and a biostratigraphic zonation of theMiddle and Upper Paleocene. Contrib Mus Paleont, Univ Michigan, 24: 135-148
Gingerich P D, 1976. Cranial anatomy and evolution of early Tertiary Plesiadapidae (Mammalia, Primates). Univ Mich Pap Paleont, 15: 1-140
Gingerich P D, 1977.Aletodon gunnelli, a new Clarkforkian hyopsodontid (Mammalia, Condylarthra) from the Early Eocene of Wyoming. Contrib Mus Paleont, Univ Michigan, 24: 237-244
Gingerich P D, 1979.Lambertocyon eximius, a new arctocyonid (Mammalia, Condylarthra) from the Late Paleocene of western North America. J Paleont, 53: 524-529
Gingerich P D, 1980.Tytthaena parrisi, oldest known oxyaenid (Mammalia, Creodonta) from the Late Paleocene of western North America. J Paleont, 54: 570-576
Gingerich P D, 1983a. New Adapisoricidae, Pentacodontidae, and Hyopsodontidae (Mammalia, Insectivora and Condylarthra) from the Late Paleocene of Wyoming and Colorado. Contrib Mus Paleont, Univ Michigan, 26: 227-255
Gingerich P D, 1983b. Paleocene-Eocene faunal zones and a preliminary analysis of Laramide structural deformation in the Clarks Fork Basin, Wyoming. Wyo Geol Assoc Guideb, 34: 185-195
Gingerich P D, 1987. Evolution and the fossil record: patterns, rates, and processes. Can J Zool, 65: 1053-1060
Gingerich P D, 1989. New earliest Wasatchian mammalian fauna from the Eocene of northwestern Wyoming: composition and diversity in a rarely sampled high- fl oodplain assemblage. Univ Mich Pap Paleont, 28: 1-97
Gingerich P D, 1991. Systematics and evolution of Early Eocene Perissodactyla (Mammalia) in the Clarks Fork Basin, Wyoming. Contrib Mus Paleont, Univ Michigan, 28: 181-213
Gingerich P D, 2003. Mammalian responses to climate change at the Paleocene-Eocene boundary: Polecat Bench record in the northern Bighorn Basin, Wyoming. In: Wing S L, Gingerich P D, Schmitz B et al. eds. Causes and Consequences of Globally Warm Climates in the Early Paleogene. Geol Soc Am, Spec Pap, 369: 463-478
Gingerich P D, 2006. Environment and evolution through the Paleocene-Eocene thermal maximum. Trends Ecol Evol, 21:246-253
Gingerich P D, 2010. Mammalian faunal succession through the Paleocene-Eocene thermal maximum (PETM) in western North America. Vert PalAsiat, 48: 308-327
Gingerich P D, Smith T, 2006. Paleocene-Eocene land mammals from three new latest Clarkforkian and earliest Wasatchian wash sites at Polecat Bench in the northern Bighorn Basin, Wyoming. Contrib Mus Paleont, Univ Michigan, 31:245-303
Granger W, 1914. On the names of Lower Eocene faunal horizons of Wyoming and New Mexico. Bull Am Mus Nat Hist, 33: 201-207
Gromova V I, 1952. On primitive carnivores of the Paleogene of Mongolia and Kazakhstan (in Russian). Tr Paleont Inst Akad Nauk SSSR, 41: 51-77
Gunnell G F, 1986. Evolutionary history of Microsyopoidea (Mammalia, Primates?) and the relationship of Plesiadapiformes to Primates. Ph.D. thesis. Ann Arbor: University of Michigan. 1-630
Gunnell G F, 1989. Evolutionary history of Microsyopoidea (Mammalia, ?Primates) and the relationship between Plesiadapiformes and Primates. Univ Mich Pap Paleont, 27: 1-157
Hou L H, 1974. Paleocene lizards from Anhui, China. Vert PalAsiat, 12: 193-200
Jepsen G L, 1930. Stratigraphy and paleontology of the Paleocene of northeastern Park County, Wyoming. Proc Am PhilosSoc, 69: 463-528
Jepsen G L, 1940. Paleocene faunas of the Polecat Bench Formation, Park County, Wyoming. Part I. Proc Am Philos Soc, 83: 217-340
Jepsen G L, Woodburne M O, 1969. Paleocene hyracothere from Polecat Bench Formation, Wyoming. Science, 164: 543-547
Kennett J P, Stott L D, 1991. Abrupt deep-sea warming, palaeoceanographic changes and benthic extinctions at the end of the Paleocene. Nature, 353: 225-229
Koch P L, Zachos J C, Gingerich P D, 1992. Correlation between isotope records in marine and continental carbon reservoirs near the Palaeocene-Eocene boundary. Nature, 358: 319-322
Krause D W, 1980. Multituberculates from the Clarkforkian Land-Mammal age, Late Paleocene-Early Eocene of western North America. J Paleont, 54: 1163-1183
Krause D W, 1982. Evolutionary history and paleobiology of early Cenozoic Multituberculata (Mammalia), with emphasis on the family Ptilodontidae. Ph.D. thesis. Ann Arbor: University of Michigan. 1-575
Krause D W, 1987a.Baiotomeus, a new ptilodontid multituberculate (Mammalia) from the Middle Paleocene of western North America. J Paleont, 61: 595-603
Krause D W, 1987b. Systematic revision of the genusProchetodon(Ptilodontidae, Multituberculata) from the Late Paleocene and Early Eocene of western North America. Contrib Mus Paleont, Univ Michigan, 27: 221-236
Li C K, Ting S Y, 1983. The Paleogene mammals of China. Bull Carnegie Mus Nat Hist, 21: 1-98
Lofgren D L, Lillegraven J A, Clemens W A et al., 2004. Paleocene biochronology: the Puercan through Clarkforkian landmammal ages. In: Woodburne M O ed. Late Cretaceous and Cenozoic Mammals of North America. New York:Columbia University Press. 43-105
Lyell C, 1833. Principles of Geology, Volume III. London: John Murray. 1-398
Magioncalda R, Dupuis C, Smith T et al., 2004. Paleocene-Eocene carbon isotope excursions in organic carbon and pedogenic carbonate: direct comparison in a continental stratigraphic section. Geology, 32: 553-556
Matthew W D, 1914. Evidence of the Paleocene vertebrate fauna on the Cretaceous-Tertiary problem. Bull Geol Soc Am, 25: 381-402
Matthew W D, Granger W, 1925. Fauna and correlation of the Gashato Formation of Mongolia. Am Mus Novit, 189:1-12
Matthew W D, Granger W, Simpson G G, 1928. Paleocene multituberculates from Mongolia. Am Mus Novit, 331: 1-4
Matthew W D, Granger W, Simpson G G, 1929. Additions to the fauna of the Gashato Formation of Mongolia. Am Mus Novit, 376: 1-12
Meng J, McKenna M C, 1998. Faunal turnovers of Palaeogene mammals from the Mongolian Plateau. Nature, 394: 364-367
Meng J, Zhai R J, Wyss A R, 1998. The Late Paleocene Bayan Ulan fauna of Inner Mongolia, China. Bull Carnegie Mus Nat Hist, 34: 148-185
Miao D S, Li C K, Wang Y Q, 2010. Minchen Chow’s academic marathon from the Bighorn Basin to the Nanxiong Basin. Vert PalAsiat, 48: 281-284
Morris W J, 1968. A new early Tertiary perissodactyl,Hyracotherium seekinsi, from Baja California. Nat Hist Mus Los Angeles Co, Contrib Sci, 151: 1-11
Novacek M J, Ferrusquía-Villafranca I, Flynn J J et al., 1991. Wasatchian (Early Eocene) mammals and other vertebrates from Baja California, Mexico: the Lomas las Tetas de Cabra fauna. Bull Am Mus Nat Hist, 208: 1-88
Osborn H F, 1910. The Age of Mammals in Europe, Asia, and North America. New York: MacMillan Company. 1-635
Phillips J, 1840. Palaeozoic series. In: Long G ed. The Penny Cyclopaedia of the Society for the Diffusion of Useful Knowledge. London: Charles Knight. 153-154
Rose K D, 1973. The mandibular dentition of Plagiomene (Dermoptera, Plagiomenidae). Breviora, Mus Comp Zool, 411:1-17
Rose K D, 1975. The Carpolestidae: early Tertiary primates from North America. Bull Mus Comp Zool, 147: 1-74
Rose K D, 1977. Evolution of carpolestid primates and chronology of North American Middle and Late Paleocene. J Paleont, 51: 536-542
Rose K D, 1979. The Clarkforkian land-mammal ‘age’ and mammalian faunal composition across the Paleocene-Eocene boundary. Ph.D. thesis. Ann Arbor: University of Michigan. 1-628
Rose K D, 1981. The Clarkforkian land-mammal age and mammalian faunal composition across the Paleocene-Eocene boundary. Univ Mich Pap Paleont, 26: 1-197
Rose K D, Krause D W, 1982. Cyriacotheriidae, a new family of early Tertiary pantodonts from western North America. Proc Am Philos Soc, 126: 26-50
Rose K D, Chew A E, Dunn R H et al., 2012. Earliest Eocene mammalian fauna from the Paleocene-Eocene Thermal Maximum at Sand Creek Divide, southern Bighorn Basin, Wyoming. Univ Mich Pap Paleont, 36: 1-122
Russell D E, 1964. Les mammifères Paléocènes d'Europe. Mém Mus Natl Hist Nat, Paris, Sér C, Sci Terre, 13: 1-324
Russell D E, Zhai R J, 1987. The Paleogene of Asia: mammals and stratigraphy. Mém Mus Natl Hist Nat, Paris, Sér C, Sci Terre, 52: 1-488
Schimper W P, 1874. Traité de Paléontologie Végétale, Volume III. Paris: J. B. Baillière. 1-896
Scott W B, 1913. A History of Land Mammals in the Western Hemisphere. New York: Macmillan Company. 1-693
Secord R, 2004. Late Paleocene biostratigraphy, isotope stratigraphy, and mammalian systematics of the northern Bighorn Basin, Wyoming. Ph.D. thesis. Ann Arbor: University of Michigan. 1-532
Secord R, 2008. The Tiffanian Land-Mammal Age (Middle and Late Paleocene) in the northern Bighorn Basin, Wyoming. Univ Mich Pap Paleont, 35: 1-192
Secord R, Gingerich P D, Smith M E et al., 2006. Geochronology and mammalian biostratigraphy of Middle and Upper Paleocene continental strata, Bighorn Basin, Wyoming. Am J Sci, 306: 211-245
Secord R, Bloch J I, Chester S G B et al., 2012. Evolution of the earliest horses driven by climate change in the Paleocene-Eocene thermal maximum. Science, 335: 959-962
Simons E L, 1960. The Paleocene Pantodonta. Trans Am Philos Soc, 50: 1-81
Simpson G G, 1928. A new mammalian fauna from the Fort Union of southern Montana. Am Mus Novit, 297: 1-15
Sinclair W J, 1912. Contributions to geologic theory and method by American workers in vertebrate paleontology. Bull Geol Soc Am, 23: 262-266
Sinclair W J, Granger W, 1911. Eocene and Oligocene of the Wind River and Bighorn basins. Bull Am Mus Nat Hist, 30:83-117
Sinclair W J, Granger W, 1912. Notes on the Tertiary deposits of the Bighorn Basin. Bull Am Mus Nat Hist, 31: 57-67
Sinclair W J, Granger W, 1914. Paleocene deposits of the San Juan Basin, New Mexico. Bull Am Mus Nat Hist, 33: 297-316
Stott L D, Kennett J P, Shackleton N J et al., 1990. The evolution of Antarctic surface waters during the Paleogene:inferences from the stable isotopic composition of planktonic foraminifera, ODP leg 113. Proc Ocean Drill Prog, Sci Res, 113: 849-863
Szalay F S, 1973. New Paleocene primates and a diagnosis of the new suborder Paromomyiformes. Folia Primatol, 19:73-87
Szalay F S, McKenna M C, 1971. Beginning of the age of mammals in Asia: the Late Paleocene Gashato fauna, Mongolia. Bull Am Mus Nat Hist, 144: 269-318
Tang Y J, Yan D F, 1976. Notes on some mammalian fossils from the Paleocene of Qianshan and Xuancheng, Anhui. Vert PalAsiat, 14: 91-99
Thewissen J G M, 1989. Evolution of Paleocene and Eocene Phenacodontidae (Mammalia, Condylarthra). Ph.D. thesis. Ann Arbor: University of Michigan. 1-279
Thewissen J G M, 1990. Evolution of Paleocene and Eocene Phenacodontidae (Mammalia, Condylarthra). Univ Mich Pap Paleont, 29: 1-107
Thewissen J G M, Gingerich P D, 1987. Systematics and evolution ofProbathyopsis(Mammalia, Dinocerata) from Late Paleocene amd Early Eocene of western North America. Contrib Mus Paleont, Univ Michigan, 27: 195-219
Ting S Y, 1998. Paleocene and Early Eocene land mammal ages of Asia. Bull Carnegie Mus Nat Hist, 34: 124-147
Ting S Y, Zhang Y P, 1979. Insectivora and Anagalida from the Chijiang Basin of Jiangxi Province. In: IVPP, NIGPAS eds. Mesozoic and Cenozoic Red Beds of South China. Beijing: Science Press. 354-358
Tong Y S, Zheng S H, Qiu Z D, 1995. Cenozoic mammal ages of China. Vert PalAsiat, 33: 290-314
Tro fi mov B A, 1952. On the genusPseudictops- a peculiar insectivore from the lower Tertiary deposits of Mongolia. Tr Paleont Inst Akad Nauk SSSR, 41: 7-12
Van Valen L M, 1978. The beginning of the age of mammals. Evol Theory, 4: 45-80
Wang Y Q, Hu Y M, Chow M C et al., 1998. Chinese Paleocene mammal faunas and their correlation. Bull Carnegie Mus Nat Hist, 34: 148-185
Wang Y Q, Meng J, Ni X J et al., 2007. Major events of Paleogene mammal radiation in China. Geol J, 42: 415-430
Wang Y Q, Meng J, Beard C K et al., 2010. Early Paleogene stratigraphic sequences, mammalian evolution and its response to environmental changes in Erlian Basin, Inner Mongolia, China. Sci China Earth Sci, 53: 1918-1926
Wood H E, Chaney R W, Clark J et al., 1941. Nomenclature and correlation of the North American continental Tertiary. Bull Geol Soc Am, 52: 1-48
Wood R C, 1967. A review of the Clark Fork vertebrate fauna. Breviora, Mus Comp Zool, 257: 1-30
Woodburne M O, Goin F J, Bond M et al., 2014. Paleogene land mammal faunas of South America: a response to global climatic changes and indigenous fl oral diversity. J Mammal Evol, 21: 1-73
Young C C, Chow M C, 1963. Cretaceous and Paleocene vertebrate horizons of North Kwangtung. Sci Sin, 12: 1411
Zhou X Y, 1995. Evolution of Paleocene-Eocene Mesonychidae (Mammalia, Mesonychia). Ph.D. thesis. Ann Arbor:University of Michigan. 1-402
美国怀俄明州大角盆地北部Polecat台地古新世动物群演化
Philip D. GINGERICH
(美国密歇根大学古生物博物馆 安阿伯 48109-1079)
怀俄明州大角盆地北部的Polecat台地和麦卡洛峰的重要发现对了解北美古新世哺乳动物生物地层以及古新世-始新世极热事件中陆生动物群更替模式和原因具有重要意义。1910年,普林斯顿大学和美国自然历史博物馆的古生物学者指出,古新世的哺乳动物与始新世的明显不同,这使得古新世是一个不同于始新世的时期的观点很快被接受。80年之后,密歇根大学和卡耐基研究院的古生物学者指出,陆相古新统-始新统界线可以用动物群的快速更替和小型化作为标志,并伴随着一次重要的碳同位素漂移,这使得古新世-始新世极热事件(PETM)作为真正的全球性温室升温事件得到了公认。1950年,投身古生物事业的年轻学者周明镇被介绍到怀俄明参加古新世哺乳动物野外工作。在那里,他积累了专业知识和经验,并促成了对中国古新统进行的类似考察。现在的挑战是如何将北美和亚洲大陆相似的古新世历史拼合在一起。
Polecat台地,麦卡洛峰,古新世,始新世,生物年代学,小型化,碳同位素漂移,古新世-始新世极热事件
P534.612-613
A 文章标号:1000-3118(2016)03-0212-23
2015-12-19
Gingerich P D, 2016. Paleocene faunal evolution at Polecat Bench in the northern Bighorn Basin of Wyoming, USA. Vertebrata PalAsiatica, 54(3): 212-234
