川西亚高山不同年龄紫果云杉径向生长对气候因子的响应
2016-03-10赵志江康东伟李俊清
赵志江,康东伟,李俊清,*
1 福建中咨工程咨询有限公司,福州 350003
2 青岛农业大学园林与林学院,青岛 266109
3 北京林业大学省部共建森林培育与保护教育部重点实验室, 北京 100083
川西亚高山不同年龄紫果云杉径向生长对气候因子的响应
赵志江1,2,康东伟3,李俊清3,*
1 福建中咨工程咨询有限公司,福州350003
2 青岛农业大学园林与林学院,青岛266109
3 北京林业大学省部共建森林培育与保护教育部重点实验室, 北京100083
摘要:运用树木年轮气候学的基本方法,建立王朗自然保护区紫果云杉在集中分布上限区域的年轮宽度年表,选取差值年表分析不同年龄云杉的径向生长同逐月气候因子的相关及响应关系,结果显示:幼龄组云杉年表的敏感度高于中龄组和老龄组云杉,幼龄组云杉对生长季前及生长季的气温状况显著正相关;中龄组云杉年表仅与当年4月份和7月份的月平均最低气温显著正相关;老龄组云杉的年轮宽度指数同上年生长季(上年8月份)的月平均气温和月平均最低温显著负相关,上年生长季高温的“滞后效应”在老龄组云杉体现的更为突出;幼龄组与中龄组云杉对当年6月份降水持续增加显示出明显的负相关关系,上年12月份的降水会对幼龄组和老龄组云杉径向生长不利。研究表明幼龄组云杉包含的气候信息要优于中龄组和老龄组云杉,在该区域进行相关研究时应根据研究需要选取不同年龄跨度的云杉年表。
关键词:紫果云杉;年龄;径向生长;气候因子
树木在自然环境下的生长主要依赖于生长期的气候条件,但是诸如微气候、上年生长、树龄等附加因子同样会对树木的生长造成一定的影响[1]。通常情况下,依据树木年轮气候学的技术操作流程,通过移除非气候“噪音”来放大树轮资料中的气候信号,这个过程称为“标准化”,其前提假设为:经过“标准化”处理之后,树木生长与气候的关系不会依赖于树木年龄而有所不同,即在给定的时间序列内不同年龄的树木对于气候的响应一致[1- 2]。但是,生理生态学的相关研究表明,与生长相关的环境信号有可能会依赖于年龄因素,例如,多数针叶树种的老树光合速率较低[3]。因此,年龄对于树木生长-气候响应的不确定性影响逐渐受到树木年轮气候学研究的重视[4]。
近些年,相关研究主要侧重于检验树木生长对于气候的响应关系在不同龄级或径级间是否一致,研究结果会因树种和地点的不同而有所差异。部分研究表明年龄不会明显影响树木年表的气候响应,例如,加拿大南部山区的幼龄与老龄落叶松(LarixlyaliiParl.)的年表对于气候响应基本一致[5- 6],欧洲中部阿尔卑斯山的五针松(Pinuscembra)不同年龄组年表所含的气候信号没有明显变化[7],浑善达克沙地不同径级油松的年表对气候因子的响应存在明显的一致性[8]。
然而,多数研究表明幼龄树与老龄树的径向生长与气候因子间的关系存在明显差异。例如,在中部斯堪的纳维亚山脉,老龄的(大于250a)欧洲赤松(PinussylvestrisL.)比中龄的对气候响应更为敏感[9];祁连山低于200a的祁连圆柏(SabinaprzewalskiiKom.)与其他4组高龄树相比,在气候响应方面存在明显的不一致[10];葡萄牙西北部的海岸松(Pinuspinaster)幼龄树的早材对于气候响应更为敏感,而老龄树的晚材更易受到更易受到气候的影响[11];我国长白山北坡低龄油松(Pinuskoraiensis)年表与高龄年表亦存在明显差异[12]。
川西亚高山位于我国青藏高原东缘,属于典型的气候敏感区域。紫果云杉(Piceapurpurea)作为该区域分布的典型优势树种,年龄因素是否会对其年表以及气候响应产生影响亟待揭示,其结果会直接影响今后利用紫果云杉树轮资料进行气候重建的可靠性。为避免出现因取样年龄不同造成掩盖部分气候信息的现象,本文旨在阐明不同龄级的紫果云杉年表对于气候因子的响应规律,以期为在本区域深入开展树轮生态学的针对性研究提供合理、科学的依据。
1材料与方法
1.1野外取样
本文研究地点位于四川省王朗国家级自然保护区,保护区面积为322.97km2,海拔跨度2300—4980m,属于丹巴-松潘半湿润季风气候,冬季寒冷干燥、日照强烈、降水较少,夏季温暖湿润、日照较少、降水集中。年均温为2.3℃,年降水量约为1100mm[13]。区内紫果云杉主要分布于阳坡海拔2600—3000m的范围内。在紫果云杉典型集中分布区的上限布设一条20m(坡向方向)×100m(水平方向)的代表性样带,样带中心海拔为3003m(32°54.2′ N,104°03.2′ E),用生长锥对样带内胸径大于10cm云杉在胸径处(离地高度为130cm)进行取样,胸径小于20cm的取1个样芯,胸径大于20cm的取1—2个样芯。由于该区域大径级(胸径大于40cm)紫果云杉分布较为分散,因此在样带外选取一定数量的大径级树进行补充取样,各采样点与样带处于同一海拔范围,以满足不同年龄样芯的需求。
1.2年龄分组及年表建立
将样芯处理完成并进行测量,根据Duncan[14]与Norton等[15]的方法判断接近和未到髓心的样芯的年龄,即树木在胸径处的年龄,然后按照100a为标准[2]将样芯分为3组:幼龄组(Young Age Classes,YAC,小于100a的样芯)、中龄组(Middle Age Classes,MAC,年龄介于100—200a的样芯)、老龄组(Old Age Classes,OAC,大于200a的样芯)[2]。分组完成后,利用COFECHA程序对各年龄组的云杉年轮序列进行质量检验并完成交叉定年[16],其中不包括心腐、破损、钻偏严重的样本,3个龄级交叉定年成功的样芯数量分别为:幼龄组取自33棵树的33个样芯,中龄组取自24棵树的27个样芯,老龄组取自18棵树的27个样芯。然后利用ARSTAN软件移除生长趋势来建立年表[17],所采用的去趋势方法为负指数函数拟合或任意斜率的线性回归拟合[2],所建立的3个龄级云杉的标准年表见图1。
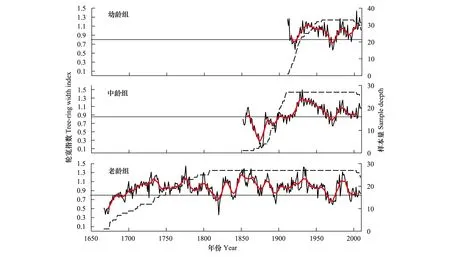
图1 王朗自然保护区不同年龄紫果云杉标准年表与样本数量Fig.1 Standard chronologies and sample numbers of P. purpurea for different age classes in the Wanglang Natural Reserve
1.3敏感度与胸径大小的关系
姜庆彪等发现轮宽样芯的敏感度同胸径大小之间存在一定的关系[8],这在一定程度上也会反映树木大小对于树木敏感性的影响。敏感度的计算公式如下:
(1)
式中,xi是第i个年轮宽度值或指数;xi+1是第i+1个年轮宽度值或指数;n是该样本年轮总数。
本文利用SimaPlot 10.0软件提供的线性回归方程计算了本研究中各交叉定年成功且到达髓心的样芯的敏感度同胸径处年龄的关系。
1.4不同年表同气候因子的关系分析
分析不同龄级云杉的差值年表同气候因子的关系,包括相关分析与响应函数分析,分析过程通过DendroCLIM 2002软件完成[18],气象数据通过山地小气候模型(MTCLIM)对采样点进行模拟,其中初始气象数据选取与采样点处于同一气候区且直线距离最近的松潘气象站(32°39′ N,103°34′ E,海拔2850m)的数据,时间跨度为1951—2009年,选取上年6月至当年9月共计16个月份的月平均气温(Tmean)、月平均最高气温(Tmax)、月平均最低气温(Tmin)与月降水(P)。
2结果与分析2.1不同龄级年表的质量与统计分析
不同龄级紫果云杉标准年表与差值年表的统计参数见表1。公共区间分析中的各参数均反映出差值年表的质量要优于标准年表。标准年表与差值年表的平均敏感度(MS)、标准差(SD)、样芯间平均相关系数(RBAR)、样本总体代表性(EPS)、信噪比(SNR)、第一特征根解释量(PC1)均体现如下趋势:即幼龄组与老龄组年表高于中龄组年表,尤以幼龄组年表最高。样芯敏感度与对应样树年龄的回归分析表明(图2),敏感度与胸径大小之间不存在线性相关关系;从散点分布上看,除部分年龄较小的样芯敏感度较大外,样芯的敏感度与年龄之间并不无明显规律。
表1王朗自然保护区不同龄级紫果云杉标准年表与差值年表的特征统计
Table 1Dendrochronological statistics for standard and residual chronologies of theP.purpureaat different age classes in the the Wanglang Nature Reserve
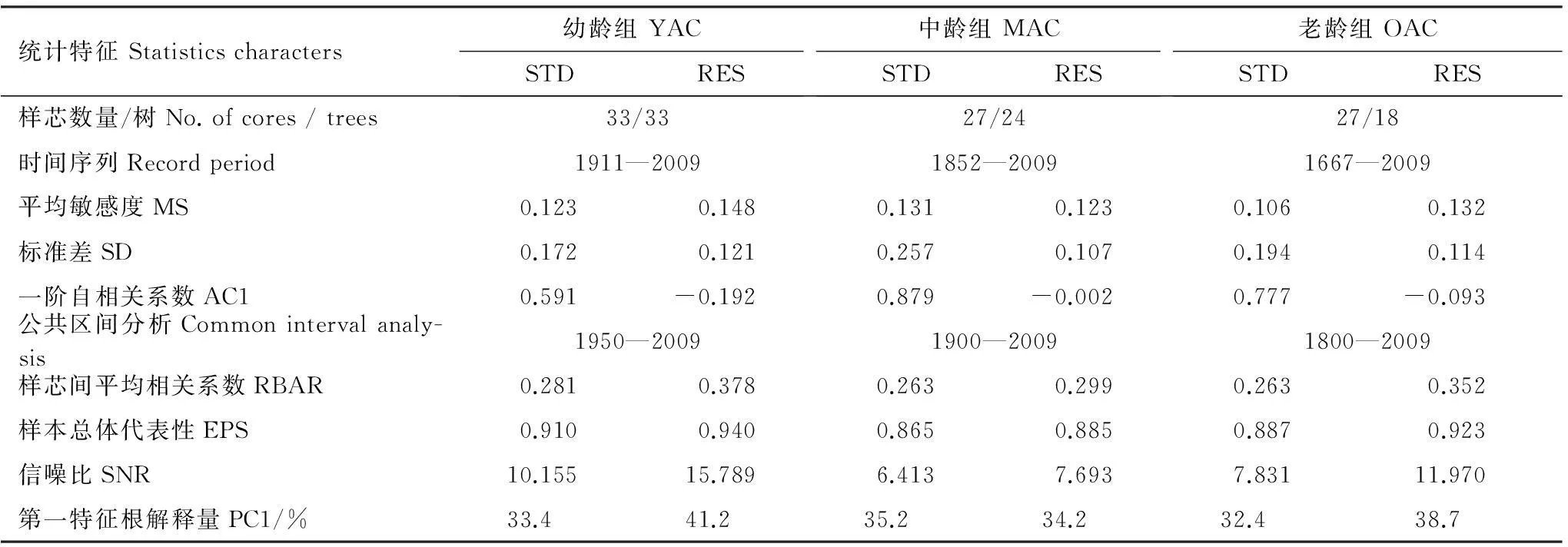
统计特征Statisticscharacters幼龄组YACSTDRES中龄组MACSTDRES老龄组OACSTDRES样芯数量/树No.ofcores/trees33/3327/2427/18时间序列Recordperiod1911—20091852—20091667—2009平均敏感度MS0.1230.1480.1310.1230.1060.132标准差SD0.1720.1210.2570.1070.1940.114一阶自相关系数AC10.591-0.1920.879-0.0020.777-0.093公共区间分析Commonintervalanaly-sis1950—20091900—20091800—2009样芯间平均相关系数RBAR0.2810.3780.2630.2990.2630.352样本总体代表性EPS0.9100.9400.8650.8850.8870.923信噪比SNR10.15515.7896.4137.6937.83111.970第一特征根解释量PC1/%33.441.235.234.232.438.7
YAC:幼龄组young age classes;MAC:中龄组middle age classes;OAC:老龄组old age classes;STD:标准年表standard chronologies;RES:差值年表residual chronologies;MS:平均敏感度 mean sensitivity;SD:标准差standard deviation;AC1:一阶自相关系数 autocorrelation order 1;RBAR:样芯间平均相关系数 mean correlation among radii;EPS:样本总体代表性 express population signal;SNR:信噪比 signal-to-noise ratio;PC1:第一特征根解释量 variance in first eigenvector
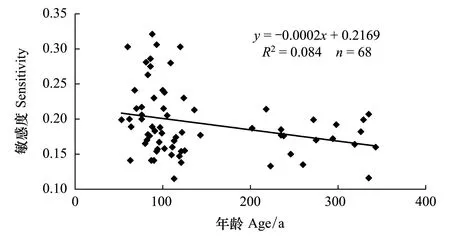
图2 王朗自然保护区紫果云杉样芯敏感度与年龄大小的相关关系 Fig.2 Correlation between sensitivity of P. purpurea samplings and ages in the Wanglang Nature Reseve
2.2不同龄级年表与气候因子的关系
由于各龄级云杉差值年表的信噪比等参数均优于标准年表,因此本文选取差值年表同各气候因子进行相关及响应分析[19],结果见图3。对于幼龄组云杉年表,相关分析的结果表明,年轮指数同下列气候因子显著正相关:上年11月份、当年2、6、9月份的月平均气温,上年11月份、当年2、9月份的月平均最高气温,当年2、3、4、6、7、8、9月份的月平均最低气温;年轮指数还同上年12月与当年6月的降水显著负相关。响应函数分析的结果表明,年轮指数与当年2月份的月平均最高气温、月平均最低气温以及当年4月份的月平均最低气温显著正相关。
对于中龄组云杉,同年轮指数具有明显相关关系的气候因子相对较少,仅在相关分析中有所体现。中龄云杉的径向生长同当年2月份、7月份的月平均最低气温显著正相关,同当年6月份的降水显著负相关。
对于老龄组云杉,相关分析显示,上年8月份的月平均气温、月平均最低气温以及当年9月份的月平均最低气温与年轮指数显著负相关;相关与响应函数分析的结果均表明上年12月份的降水对径向生长起明显的负作用。

图3 不同龄组紫果云杉差值年表与月气候因子的相关及响应关系(*P<0.05)Fig.3 Correlations and responses between P. purpurea residual chronology with different age classes and monthly climatic factors in the Wanglang Nature Reserve (* P<0.05)
3讨论
3.1树龄对年表参数的影响
通过3个龄级年表特征对比(表1),发现幼龄云杉年表的质量明显高于中龄与老龄云杉年表,中龄云杉的年表质量最差,说明幼龄云杉对于周边环境的变化更为敏感,样芯敏感性与年龄大小关系也印证了这一点(图2)。姜庆彪等在对浑善达克沙地不同径级油松(Pinustubulaeformis)研究时也发现,小径级油松具有更高的敏感度[8]。王婷分析了天山云杉(Piceaschrenkiana)随年龄变化的年表特征,结果显示中龄组的年表质量要优于幼龄组与老龄组[20]。一般认为大径级[21]或大龄级[22]树木对于气候响应更加的敏感,与本研究的结果有所差别,主要原因可能是不同地点不同树种随年龄(径级)变化的生理过程不一致[8]。在王朗自然保护区紫果云杉的主要分布区,幼龄或老龄云杉更适合进行气候响应分析研究。
3.2不同树龄年表对气候因子的响应差异
不同树龄年表对于气温的敏感性存在很大的差异性。幼龄组云杉对生长季前(上年11月份、当年2月份)及生长季(6月份、9月份)的气温状况均显示出明显的相关性,生长季前及生长季升温均有利于其径向生长,特别是当年2、3、4月份及6、7、8、9月份的月平均最低气温。生长季前的升温有利于幼龄云杉度过寒冷冬季,减少冻害的发生,同时通过增强细胞活性、促进营养物质运输等方式为树木生长做好充足准备[8]。生长季初(当年6月份)的升温能够提高云杉体内与生长相关的酶活性、提高光合速率、促进营养物质积累。生长季末(当年9月份)较高的气温有利于延长生长期时间,促进宽轮的产生。在这一区域最低气温有可能是影响幼龄云杉形成层活性的关键因素,由此限制木质部的管胞分裂与扩展[23]。生长季较低的气温与土壤温度会通过影响水分有效性限制形成层活性,进而不利于树木径向生长[24]。而中龄组云杉差值年表仅与当年4月份和7月份的月平均最低气温显著正相关,中龄组云杉标准年表同气候的关系还显示当年6月份较高的月平均气温和月平均最低气温有利于云杉径向生长,说明生长季前及生长季低温是限制中龄云杉径向生长的主要气温因子,即中龄组云杉生长对当年生长季前及生长季最低气温较为敏感。与幼龄组和中龄组云杉不同,老龄组云杉的差值年表年轮宽度指数同上年生长季(上年8月份)的月平均气温和月平均最低温显示出明显的负相关关系,另,标准年表的气候相关分析也显示上年7月份、8月份和当年9月份的月平均气温和月平均最高气温与轮宽指数显著负相关。可见生长旺季和生长季末的高温会过度消耗云杉体内储存的光合产物,不利于云杉径向生长。
不同龄级云杉生长对于降水的响应也不完全一致。幼龄组与中龄组云杉对当年6月份降水持续增加显示出明显的负相关关系,可能的原因是生长季充沛降雨造成云杉根部的无氧呼吸,从而消耗较多的有机物限制年轮宽度增加[25]。而幼龄组与老龄组云杉年表同冬季(上年12月份)的降水显著负相关,12月份过多的降水会加剧云杉的冻害或机械损伤,从而影响到下年的生长[26]。
目前,还不能从机理上深入揭示不同龄级云杉生长对于气候因子的响应差异,部分学者认为遗传基因[2]和林分动态[27]等造成的树木生理机能的不同是其主要原因。树龄不同,则生理机能不同,随之树木对气候的响应也会不同。例如,幼龄云杉在生长季末(当年9月份)会受到低温胁迫,过早的遭受冻害,从而降低活性影响生长;而对于老龄云杉,由于长期的适应过程,会对早期冻害产生一定的抗性不易受损,但是生长季末(当年9月份)的高温会增强呼吸速率,不利于有机物质的积累。幼龄与老龄云杉对于冬季(上年12月份)的降水更加敏感,而对于位于生长壮年的中龄云杉具有更强的抵御冬季严寒的能力。幼龄和中龄云杉的生长会受到当年生长季(当年6月份)充足降水的抑制,而对于老龄云杉而言,由于枝干粗壮、冠幅较大对于水分的需求量大,会降低根部无氧呼吸的强度。
4结论
在王朗自然保护区紫果云杉的集中分布上限区域,幼龄组云杉年表的统计参数要优于中龄组与老龄组云杉年表,与之相对应,幼龄组云杉对于环境的敏感性也高于其他两个龄组。在温度方面,幼龄组云杉生长同当年的气温状况表现出良好的相关性,老龄组云杉生长主要受到上年生长季高温的显著抑制,中龄组云杉生长主要得益于当年4月份和7月份较高的低温。在降水方面,当年6月份持续降水将会明显不利于幼龄组与中龄组云杉的生长,上年12月份的降水亦会对幼龄组和老龄组云杉产生抑制作用。鉴于此,本文认为今后在进行树轮气候学研究时,应根据研究需要恰当选取建立年表的样芯的年龄跨度,以免损失或放大气候信息。
致谢:感谢王朗自然保护区的大力支持。
参考文献(References):
[1]吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990: 44-145.
[2]Szeicz J M, MacDonald G M. Age-dependent tree-ring growth responses of subarctic white spruce to climate. Canadian Journal of Forest Research, 1994, 24(1): 120-132.
[3]Bond B J. Age-related changes in photosynthesis of woody plants. Trends in Plant Science, 2000, 5(8): 349-353.
[4]彭剑峰, 刘玉振, 王婷. 神农山白皮松不同龄组年轮-气候关系及PDSI 重建. 生态学报, 2014, 34(13): 3509-3518.
[5]Colenutt M E, Luckman B H. The dendrochronological characteristics of alpine larch. Canadian Journal of Forest Research, 1995, 25(5): 777-789.
[6]Colenutt M E, Luckman B H. Dendrochronological investigation ofLarixlyalliiat Larch Valley, Alberta. Canadian Journal of Forest Research, 1991, 21(8): 1222-1233.
[7] Esper J, Niederer R, Bebi P, Frank D. Climate signal age effects-Evidence from young and old trees in the Swiss Engadin. Forest Ecology and Management, 2008, 255(11): 3783-3789.
[8]姜庆彪, 高露双, 王晓明, 赵秀海. 浑善达克沙地油松树轮宽度与气候因子的关系. 应用与环境生物学报, 2012, 18(3): 405-410.
[9]Linderholm H W, Linderholm K. Age-dependent climate sensitivity ofPinussylvestrisL. in the central Scandinavian Mountains. Boreal Environment Research, 2004, 9(4): 307-317.
[10]Yu G R, Liu Y B, Wang X C, Ma K P. Age-dependent tree-ring growth responses to climate in Qilian juniper (SabinaprzewalskiiKom.). Trees, 2008, 22(2): 197-204.
[11]Vieira J, Campelo F, Nabais C. Age-dependent responses of tree-ring growth and intra-annual density fluctuations ofPinuspinasterto Mediterranean climate. Trees, 2009, 23(2): 257-265.
[12]王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡不同年龄红松年表及其对气候的响应. 生态学报, 2011, 31(21): 6378-6387.
[13] Taylor A H, Jang S W, Zhao L J, Liang C P, Miao C J, Huang J Y. Regeneration patterns and tree species coexistence in old-growthAbies-Piceaforests in southwestern China. Forest Ecology and Management, 2006, 223(1-3): 303-317.
[14]Duncan R P. An evaluation of errors in tree age estimates based on increment cores in Kahikatea (Dacrycarpusdacrydioides). New Zealand Natural Sciences, 1989, 16: 31-37.
[15]Norton D A, Palmer J G, Ogden J. Dendroecological studies in New Zealand 1. An evaluation of tree age estimates based on increment cores. New Zealand Journal of Botany,1987, 25(3): 373-383.
[16]Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983,43: 69-78.
[17]Cook E R. A Time Series Analysis Approach to Tree Ring Standardization (Dendrochronology, Forestry, Dendroclimatology, Autoregressive Process)[D]. Tucson: The University of Arizona, 1985.
[18]Biondi F, Waikul K. DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303-311.
[19]王晓明, 赵秀海, 高露双, 姜庆彪. 长白山北坡林线处岳桦年轮年表及其与气候的关系. 应用与环境生物学报, 2012, 18(1): 9-16.
[20]王婷. 天山中部不同海拔高度天山云杉林的生态学研究[D]. 武汉: 武汉大学, 2004.
[21]Chhin S, Hogg E H, Lieffers V J, Huang R M. Potential effects of climate change on the growth of lodge pole pine across diameter size classes and ecological regions. Forest Ecology and Management, 2008, 256(10): 1692-1703.
[22]Carrer M, Urbinati C. Age-dependent tree-ring growth responses to climate inLarixdeciduaandPinuscembra. Ecology, 2004, 85(3): 730-740.
[23]Deslauriers A, Morin H, Begin Y. Cellular phenology of annual ring formation ofAbiesbalsameain the Quebec boreal forest (Canada). Canadian Journal of Forest Research, 2003, 33(2): 190-200.
[24]Liang E Y, Wang Y F, Xu Y, Liu B, Shao X M. Growth variation inAbiesgeorgeivar.smithiialong altitudinal gradients in the Sygera Mountains, southeastern Tibetan Plateau. Trees, 2010, 24(2): 363-373.
[25]Lloyd A H, Graumlich L J. Holocene dynamics of treeline forests in the Sierra Nevada. Ecology, 1997, 78(4): 1199-1210.
[26]徐倩倩. 长白山东坡长白落叶松生长和气候因子关系的研究[D]. 北京: 北京林业大学, 2011.
[27] Szeicz J M, Macdonald G M. Recent white spruce dynamics at the subarctic alpine treeline of north-western Canada. Journal of Ecology, 1995, 83(5): 873-885.
Age-dependent radial growth responses ofPiceapurpureato climatic factors in the subalpine region of Western Sichuan Province, China
ZHAO Zhijiang1,2, KANG Dongwei3, LI Junqing3,*
1FujianCIECCEngineeringConsultingCo.Ltd,Fuzhou350003,China
2CollegeofLandscapeArchitectureandForestry,QingdaoAgriculturalUniversity,Qingdao266109,China
3TheKeyLaboratoryforSilvicultureandConservationofMinistryofEducation,BeijingForestryUniversity,Beijing100083,China
Abstract:Although tree growth is mainly dependent on natural climate conditions, factors such as microclimate, growth of the previous year, tree age, and others may also affect tree growth. In dendroclimatological studies, it is generally assumed that the relationships between tree growth and climate factors are age-independent after removing the biological growth trends related to tree age. However, in some multi-aged forest stands, climate-growth relationships may be biased, because the trees′ chronologies respond differently to climate factors depending on their age or size. To assess this age-dependent effect, in this study, we tested the consistency of climate-growth responses in tree-ring series from Picea purpurea trees of different age classes in the Wanglang Nature Reserve, Sichuan province. In the concentrated distribution area of the upper limit, spruces were grouped into three age classes: trees younger than 100 years (young age class, YAC), trees 100—200 years of age (middle age class, MAC), and trees older than 200 years (old age class, OAC). Residual chronologies of the three age classes were built to analyze the climate-growth relationships using correlation and response functions. Correlation analysis indicated that the radial growth of the YAC was significantly and positively correlated with mean monthly temperature of the previous November and current February, June, and September; mean monthly maximum temperature of the previous November and current February and September; and mean monthly minimum temperature of the current February, March, April, June, July, August, and September, and it was significantly and negatively affected by monthly precipitation of the previous December and current June. The ring width of the MAC was positively correlated with mean monthly minimum temperature of the current April and July, and was negatively correlated with monthly precipitation of the current June. The residual chronology of the OAC was significantly and negatively affected by mean monthly temperature of the previous August, mean monthly minimum temperature of the previous August and current September, and monthly precipitation of the previous December. Response analysis showed that radial growth of the YAC was significantly and positively correlated with mean monthly maximum temperature of the current February, and mean monthly minimum temperature of the current February and April. There was no significant correlation between radial growth of the MAC and climate factors. The ring width of the OAC was significantly and negatively affected by monthly precipitation of the previous December.Overall, our results revealed that with an increase in tree age, the sensitivity of spruce to climate factors was reduced. The responses of young spruce were significantly correlated with temperature before the growing season and in the growing season. The chronology of the middle-aged spruce showed a significant and positive correlation with monthly mean minimum temperatures in the current April and July. The ring-width index of old spruce was significantly negatively correlated with monthly mean temperature and monthly mean minimum temperature of the previous August. The “lag effect” of high temperatures in the previous growing season was prominent in the old spruce. Spruce within the young and middle-aged groups showed a significant negative correlation with current June precipitation. Adequate precipitation in December was not beneficial for the radial growth of young and old spruce. Our study may demonstrate that future studies should consider the adaptive chronology of age span for spruce in this area to avoid losing or magnifying the climate signals.
Key Words:Picea purpurea; age; radial growth; climatic factors
DOI:10.5846/stxb201409121815
*通讯作者Corresponding author.E-mail: lijq@bjfu.edu.cn
收稿日期:2014- 09- 12;
修订日期:2015- 06- 08
基金项目:国家林业公益性行业科研专项(201404422);青岛农业大学高层次人才科研基金项目(631403)
赵志江,康东伟,李俊清.川西亚高山不同年龄紫果云杉径向生长对气候因子的响应.生态学报,2016,36(1):173- 179.
Zhao Z J, Kang D W, Li J Q.Age-dependent radial growth responses ofPiceapurpureato climatic factors in the subalpine region of Western Sichuan Province, China.Acta Ecologica Sinica,2016,36(1):173- 179.
