大青鲨渔业生物学研究进展
2016-03-06李伟文田思泉戴小杰陈新军
李伟文,田思泉,2,3,4,戴小杰,2,3,4,陈新军,2,3,4
(1.上海海洋大学海洋科学学院 201306;2.上海海洋大学国家远洋渔业工程技术研究中心 201306;3.上海海洋大学大洋渔业可持续开发教育部重点实验室 201306;4.上海海洋大学农业部大洋渔业资源环境科学观测实验站 201306)
·综述·
大青鲨渔业生物学研究进展
李伟文1,田思泉1,2,3,4,戴小杰1,2,3,4,陈新军1,2,3,4
(1.上海海洋大学海洋科学学院 201306;2.上海海洋大学国家远洋渔业工程技术研究中心 201306;3.上海海洋大学大洋渔业可持续开发教育部重点实验室 201306;4.上海海洋大学农业部大洋渔业资源环境科学观测实验站 201306)
大青鲨(Prionace glauca)广泛分布于南、北温带和热带的三大洋区,是延绳钓渔业和刺网渔业的主要兼捕对象,对维持海洋生态系统的稳定性和多样性起着重要的作用。随着捕捞强度的加大,大青鲨的资源出现衰退的现象。但由于基础数据的不确定性,大青鲨资源评估的结果并未能得到很好的应用。本文以文献计量统计分析为基础,对大青鲨渔业生物学的研究历程进行阐述,了解了不同时期大青鲨渔业生物学的研究重点。综合诸多文献资料,对大青鲨的年龄与生长、基础繁殖生物学、种群结构的鉴定、资源丰度指数和资源评估等方面的研究方法进行综述和分析。最后根据上述的综合和分析结果,提出了今后相关研究的注意事项以及相关的方法,以进一步提高大青鲨的资源评估的可信度,提高对大青鲨资源现状的了解,以期为大青鲨的资源养护提供科学依据。
大青鲨;渔业生物学;资源丰度指数;评估模型
大青鲨(Prionace glauca)隶属于真鲨目(Carcharhiniform),真鲨科(Carcharhinidae),大青鲨属,广泛分布于温带和热带水域的近岸及洋区,最长可达300 cm[1-3]。大青鲨是延绳钓渔业和刺网渔业的主要兼捕对象,每年大青鲨的兼捕量可达6 500 000~11 000 000 ind[4-6]。大青鲨生命周期长,生长慢、成熟晚、繁殖力低,导致了其较低的内禀增长率和种群恢复能力。大青鲨位于海洋食物链的顶端,是海洋生态系统的重要组成部分,对维持海洋生态系统的稳定起着极其重要的作用[7]。大青鲨的鲨鱼翅是传统的海产珍品,鱼肉可以制作鱼丸,随着全球鱼翅贸易的扩大,大青鲨的捕捞量出现接近或可能超过最大持续产量的情况,并被国际保护联盟(IUCN)确定为近危物种[7]。
为了大青鲨渔业资源的可持续利用,维护海洋生态系统的稳定,避免大青鲨种群面临资源衰退甚至崩溃的危险,大西洋金枪鱼资源保护委员会(International Commission for the Conservation of Atlantic Tunas,ICCAT)已经要求缔约方和非缔约方上报该鱼种的渔业数据,并准备对该种群进行科学的评估[8-9]。国际社会对于该渔业资源的养护和管理提上日程,各区域性渔业管理组织逐渐开始进行大青鲨的资源评估研究,并提出相应的管理措施[10-12]。
在渔业日益发展的今天,各个国家和地区逐渐意识到了保护鲨鱼以实现鲨鱼可持续利用的重要性,为此,探讨鲨鱼的科学评估势在必行。但是大青鲨的资源评估由于基础数据的不确定性,导致了其结果并未能得到很好的应用。而大青鲨的生长、基础繁殖生物学及其种群结构的鉴定是进行合理评估的基础,有效的资源丰度指数是种群评估的重要指标,合理的模型选择是进行科学评估的有效工具。不同的学者对大青鲨的生长、繁殖生物学、种群结构、资源的丰度指数及其评估模型进行了一定的研究,本文将各个研究结果进行概述,旨在提高对大青鲨资源现状的了解,以期对大青鲨的资源养护提供科学依据。
1 大青鲨文献计量统计
以“web of science”数据库为基础,该数据库并未收集所有期刊的文献,所以在统计上主要是探讨鲨鱼资源学的发展动态,以期为后续的研究提供研究方向。以“shark”和“resource”为主题词进行文献检索,针对检索结果做文献计量统计。共检索到文献389篇,其中期刊类论文347篇,会议性论文52篇,其余文献类共计23篇。
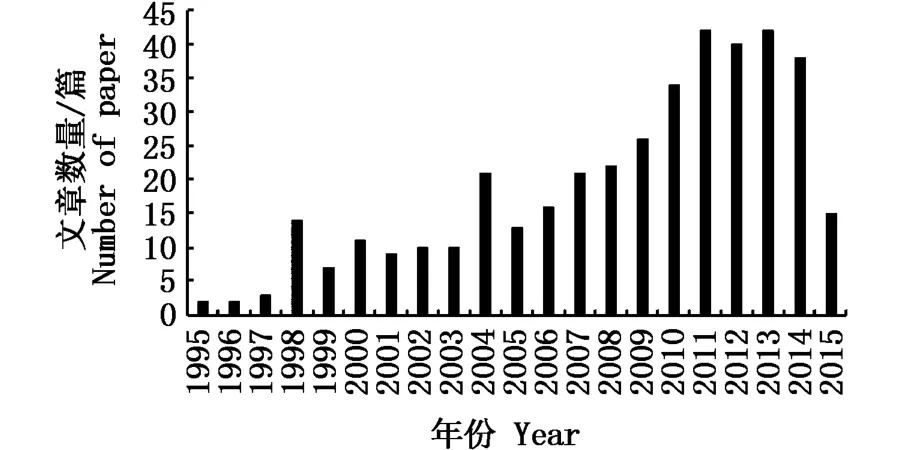
图1 大青鲨渔业生物学文章的年份分布Fig.1 Articles on fishery biology of blue shark in various years
关于鲨鱼资源类的研究论文在web of science上最早见于1995年,且当年仅有2篇相关的论文发表。随后,相关的研究呈现逐渐增长趋势,2010以后,文献数量出现了较大的增长,这与鲨鱼的资源量的下降和相关渔业组织对鲨鱼的关注度增高相关(图1)。在相关发表的文献中,美国、澳大利亚和加拿大是研究鲨鱼最多的国家,三个国家相关的研究论文数量达256篇。
在统计的389篇文献中,以web of science上45次以上引用文章为高引文献,共有54篇文章为高引文献。结合文献统计中十年的发展趋势,在1995~1999年,学科中研究较为热门的主要是集中在鲨鱼整体的分布现状、鲨鱼可持续发展的探讨以及鲨鱼在生态系统中的作用等,主要针对鲨鱼群体的研究,对单一鲨鱼品种的研究较少。2000~2009年的10年间,鲨鱼的兼捕和保护、摄食生态、单一品种鲨鱼的评估、繁殖生物学、分子生态学、资源丰度探讨等方向逐渐深入,并逐渐形成体系。从2010年至今,基于生态系统的管理模式引起了人们的关注,资源评估模型应用和计算手段的提升,成了现阶段鲨鱼资源评估的焦点,也是实现鲨鱼可持续发展的必经之路。
2 大青鲨的年龄与生长
大青鲨的生长速度与大青鲨的资源蕴藏量存在紧密的关系,所以,研究大青鲨的年龄和生长是研究大青鲨资源状况的重要内容之一,科学地掌握大青鲨的年龄和生长的信息有助于预测大青鲨资源的变动,可以为大青鲨的渔业管理措施制定提供科学的理论指导[13]。
2.1 生长方程
在鲨鱼的年龄与生长的研究方面上,不同的学者采用了不同的生长模型对鲨鱼的生长状况进行描述。表1列出了8种最常见的描述鲨鱼生长的模型,共描述了15种鲨鱼[14],2004年以前,关于大青鲨生长状况的描述只局限于 Von Bertalanffy模型,同时,Von Bertalanffy模型也是应用最广泛的一种模型。近年来,学者们采用了多种生长方程模型对同一种鲨鱼的生长进行描述,利用AIC(Akaike Information Criterion)信息量准则筛选最适的模型,减少了模型带来的误差。2004年,LESSA等[15]首次利用了Von Bertalanffy模型,Richards模型和Schnute模型三种模型描述了大青鲨的生长,经AIC信息量准则筛选后证明Von Bertalanffy模型最能科学的描述大青鲨的生长。
2.2 生长参数的估算
AASEN[24]最早开始利用长度频度分析的方法分析估算了大青鲨的相关生长参数,接着STEVENS[25]利用了脊椎骨对大青鲨的年龄进行鉴定,并估算相关的生长参数。1979年,TANAKA等[26]首次利用了Von Bertalanffy生长模型分析了北太平洋大青鲨雌性的生长参数,1994年,NAKANO[3]对北太平洋的大青鲨做了类似的分析,其结果与TANAKA得出的结果相似。SKOMAL等[27]、高春霞等[28]对大西洋大青鲨的相关生长参数进行估算(表2),两者的结果并没有显著的差异,但是与NANKANO[3]对北太平洋大青鲨分析后所得的生长参数具有显著的差异,可能是北太平洋大青鲨种群与北大西洋大青鲨种群分属两个具有显著分化的种群,导致了其在生长上出现了显著的差异。
3 大青鲨的繁殖生物学
大青鲨群体的繁殖活动是其生命活动的最主要的组成部分,是其独特的繁殖特性与群体对特定水域的生活环境长期适应的结果[13]。大青鲨的繁殖力直接关系着补充群体的数量,独特的胎生方式提高了其后代的成活率,保证了补充群体的数量。合理地掌握大青鲨的繁殖机理,有助于提供合理的捕捞规格及其数量,制定相应的科学管理措施。为此,国内外学者对大青鲨的繁殖生物学做了一系列的研究,以期为合理地进行大青鲨的资源评估提供基础依据。
3.1 繁殖习性
在北大西洋,大青鲨的繁殖种群从北美东岸扩张到欧洲西海岸[29]。大青鲨交配的时间发生在5月末到11月初之间,受精后大约9~12个月后产仔[7]。在西北大西洋,大青鲨的种群主要是由未成熟的雄性和雌性以及成熟的雄性群体组成[30-31]。虽然在西北大西洋的大青鲨种群中有一小部分的成熟雌性个体,但是一些未成熟的雌性个体的输卵管中有一些新鲜精子的存在,并在它们的身体上留有新鲜的咬痕,表明了一些未成熟的雌性个体已经开始交配[31-32]。
大青鲨的繁殖方式为胎生,在子宫内发育的鲨鱼仔由卵黄胎盘与子宫内膜连接,解剖雌体体腔时,可以发现怀孕个体的子宫一般出现膨大。戴小杰等[33]于2003年7月~11月,对东太平洋热带公海海域大青鲨的繁殖生物学特征进行了研究,吴峰等[34]于2007年12月~2008年3月,对热带中东大西洋海域的大青鲨的繁殖生物学特征也进行了相关研究。在大青鲨种群的雌雄性比问题上,研究结果一致表明,在子宫中,雌雄胎儿数性比为1∶1[33-34];且雄性大青鲨精巢和总性腺重量均随鱼体全长的增加而增加,雌性大青鲨卵巢重量随鱼体全长变化不大。

表1 鲨鱼中常用的生长模型Tab.1 Grow th models commonly used for sharks

表2 大青鲨Von Bertalanffy生长参数Tab.2 Von Bertalanffy grow th parameters for blue shark
3.2 初次性成熟长度
虽然大青鲨分布广泛,但是其性成熟状况从未被直接观察,所以对大青鲨的繁殖习性了解较少[35]。目前关于大青鲨的繁殖特性主要来源于实体解剖、标志跟踪调查[29]和基于对其它大鲨鱼种类的直接观察后进行推断[36]。不同的方法用于掌握大青鲨的交配习性以及初次性成熟对应的长度,这些议题对于深入了解种群结构具有重要作用。
表3统计了不同学者针对不用海域对大青鲨初次性成熟的体长及其对应的年龄。目前共研究了4个太平洋海域,2个印度洋海域以及7个大西洋海域,根据研究结果对比,三个海域大青鲨对应的初次性成熟的体长及其对应的年龄并不存在显著的差异。
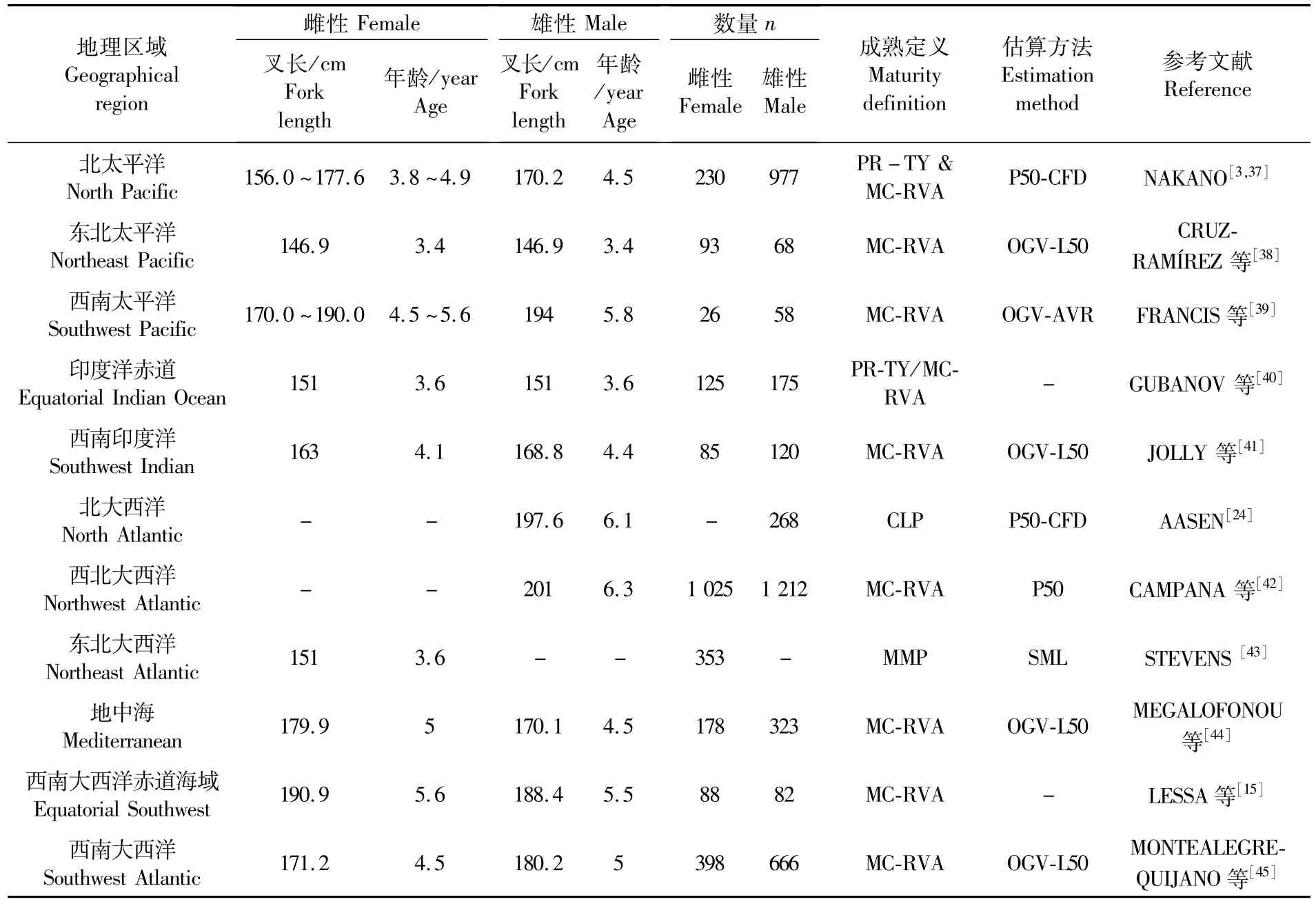
表3 全球估算大青鲨初次性成熟的长度及其对应的年龄Tab.3 Global estimates of size at maturity of blue shark P.glauca and their respective calculated ages
3.3 繁殖力分析
大青鲨是繁殖能力最强的大型鲨鱼之一,在四种大洋性真鲨的繁殖力分析中,大青鲨的繁殖力最高,高于镰状真鲨(Carcharhinus falciformis)、长鳍真鲨(Carcharhinus longimanus)和路氏双髻鲨(Sphyrna lewini)[45]。NAKANO[3]对北太平洋669 ind怀仔鱼雌体进行观察后发现,每胎产仔鱼数量为1~62 ind,平均每胎25.6 ind;CARRERAFERNÁNDEI等[46]在研究墨西哥加利福利亚海岸大陆架水域妊娠期的大青鲨中发现,不同发育阶段的胎儿数范围为27~33 ind;戴小杰等[33]观察到东太平洋海域大青鲨的每胎产仔鱼数量为13~62 ind;吴峰等[34]的调查结果,热带大西洋海域大青鲨怀胎儿数量为3~50 ind,平均33 ind。以上研究表明各个水域大青鲨的繁殖力具有一定的差异性,同时也说明了大青鲨具有很强的繁殖能力。高繁殖力表明大青鲨在大洋性生态系统中具有较强的适应能力,同时也是其广布各大洋、成为优势种的原因之一。
4 大青鲨种群结构现状分析
大青鲨种群结构的鉴定直接关系到大青鲨资源数量变动及生活习性的深入研究,只有在充分了解大青鲨种群结构的基础上,才能对大青鲨资源的合理利用和管理提出科学的依据。
4.1 标志法分析大青鲨种群结构
对太平洋大青鲨进行标志放流研究的主要国家是美国、新西兰和日本3个国家(表4)。通过对这些标志的结果进行分析,绘制太平洋大青鲨标志放流结果分布图(图2)。结果显示南北太平洋大青鲨群体的运动并没有跨过赤道,表明了南北太平洋大青鲨群体分属两个不同的亚种群。但值得注意的是,其中有1 ind大青鲨从南太平洋跨界洄游到印度洋,暗示了太平洋与印度洋大青鲨群体间可能存在群体交流。
在大西洋,大青鲨的种群结构并不清晰。但现在假设至少存在3个种群,北大西洋种群、南大西洋种群和地中海种群[48]。主要是北大西洋和南大西洋种群的分界线位于5°N,而地中海种群则以直布罗陀海峡作为分界线。种群的这些假设来源于标志回捕的数据分析,根据北大西洋种群的跨洋区穿越的数据分析,只有极少数的标志回捕记录会穿越赤道[49-50]。进一步分析,对整个大西洋的大青鲨样本进行微卫星标志分析,表明北大西洋和南大西洋分属两个不同的遗传种群[48]。
4.2 大青鲨种群结构的分子标记分析
近年来,相关的渔业管理组织开始对太平洋大青鲨进行种群评估。为了能够获得有效的种群评估,种群的结构现状是评估的基础。大青鲨是分布最为广泛、种群数量最多的一种大洋性鲨鱼[51],尽管大洋交界处会有不同地理种群的个体混合,但标志重捕研究表明,大青鲨的运动范围局限在其本身所在的海域,各海域间大青鲨的基因交流很少[52]。
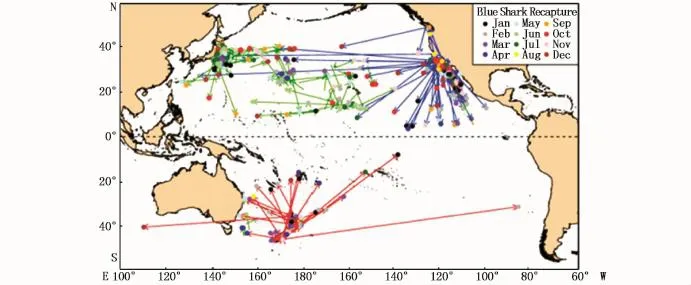
图2 太平洋大青鲨标志放流结果分布[47]Fig.2 Movement routes of blue shark in the Pacific[47]

表4 太平洋大青鲨标志的主要国家[47]Tab.4 Blue shark tagging conducted countries in the Pacific[47]
郑真真等[53]利用线粒体DNA的控制区和COⅠ(细胞色素氧化酶亚基Ⅰ)基因的部分序列分子标记来研究全球大青鲨种群遗传结构和遗传多样性,样本来自中东太平洋、中西太平洋、中东大西洋、西南大西洋和印度洋5个海域,研究结果显示五个群体间未形成显著的遗传结构,且三大洋群体间基因交流频繁,不存在显著的遗传分化,遗传多样性较高;认为三大洋的大青鲨为一个随机交配的群体,各个采样点间基因交流频繁,并且遗传多样性水平都呈下降趋势。TAGUCHI等[54]基于线粒体细胞色素b基因,对来自太平洋和太平洋与印度洋跨界的404 ind大青鲨样本做了种群结构鉴定,结果显示10个采样位点间的采样群体存在广泛的基因交流,不存在显著的遗传分化,为单一种群,其结果与郑真真等[53]的研究结构高度相似。
5 大青鲨资源丰度分析
大青鲨的资源丰度指数是反映资源状况的一个指标,从某种程度上反映了资源的整体变化趋势,在渔业过程中,通常采用单位捕捞努力量(catch per unit of effort,CPUE)表示,是大青鲨资源评估过程中一个重要的参数,直接影响着大青鲨资源评估的结果。选择合适的资源丰度指数,对于准确评估大青鲨的资源状况具有重要的意义[13]。
日本是大西洋公海延绳钓主捕金枪鱼重要的国家,其延绳钓渔业在大西洋作业的历史最长,最具有代表性。根据日本渔捞日志报告记录,其延绳钓渔业在热带大西洋(20°N~20°S海域)捕获的大青鲨1993~1999年的CPUE在每千钩3.95~1.24 ind之间,其观察员获得的大青鲨1995年的CPUE为5.96,1997年为1.73,而1999年仅仅为0.87,渔捞日志数据和观察员的数据均呈下降趋势[55-56]。
戴小杰等[8]在1994~2001年期间,对热带大洋性中上层鲨鱼资源进行了调查。海上调查共4个航次,在第1~3航次的CPUE达到每千钩5~7 ind,而在第4航次下降为每千钩0.8 ind,下降84%~88%。从大青鲨资源密度指标来看,无论是以重量计算或以尾数计算的CPUE都呈下降趋势。反映渔业资源状况的另一个指标就是渔获物的年龄组成,从年龄组成的角度看,随着捕捞强度的增加,渔获物的年龄组成必然下降,反映在鱼体的平均长度或者重量也呈下降趋势。调查结果显示,目前兼捕大西洋中上层鲨鱼的捕捞努力量在增加,资源呈下降趋势。
6 大青鲨资源评估分析
大青鲨的资源评估是基于科学观察员调查数据、海上渔捞日志以及港口调查等数据,利用相关的渔业资源评估模型,科学估算大青鲨及其种群的相关参数,以回溯大青鲨的渔业捕捞历史,评估大青鲨捕捞活动,鲨鱼管理对大青鲨资源利用的影响,并对大青鲨的渔业资源发展趋势进行预测和风险评估[13]。因此,渔业资源评估是渔业资源科学管理的基础。
在20世纪60年代和20世纪70年代,大多数鲨鱼的种群评估主要使用了logistic Schaefer模型[57],主要是因为数据的缺乏和计算能力的不足。这种模式往往证明不了鲨鱼种群动态模型,因为它被认为是生产过剩和资源生物量之间的关系,在最大值之间对称分布,介于零资源生物量和承载能力之间[58]。PELLA等[59]提出的补充形状参数,允许生产关系向左或向右倾斜。尽管这个模型具有灵活性和稳定性,由于数量成反比关系的参数估计和模型的性能,使得这个模型可能操作起来比Schaefer模型更不准确[60]。
在最近的大西洋大青鲨的种群评估中,大西洋金枪鱼养护与管理委员会认为大西洋金枪鱼的渔业数据得到了很大的改善。2008年,在对大西洋北部种群和南部种群的评估中开发了一种新的评估模型,称为生态风险评估(Ecological risk assessment,ERA)[61-62]。种群评估的结果显示北部和南部大西洋种群均没有遭受过度捕捞,也没有正在遭受过度捕捞。
生态风险评估推断,大青鲨是中上层延绳钓渔业中的中度易受损物种。然而,依据严格的条件假设,评估的结果显示大青鲨为高度的易受损物种,原因在于其相关定量和定性信息的缺乏。这就要求通过不断的努力以解决种群结构和种群统计学的不确定性,开发没有偏移的相对丰度指数和完整的历史捕捞系列数据,以改善将来大西洋大青鲨的资源评估工作。
根据CORTÉS[63]的研究结果,大青鲨的繁殖力是记录中最高的,令人惊讶的是其早期的生长速度很快,以致于其经过第1年的生长就可以达到幼仔大小的两倍[27,64]。尽管贝叶斯估计方法在减少数据产生误差的统计上具有一定的优势,但值得注意的是先验分布的选择可以改变种群评估后的状态,特别是当数据信息不足的条件下对评估结果的影响较大。目前的研究结果显示,经过统计学分析后,也包含了预测后的r,南大西洋大青鲨的内禀增长率的先验概率分布增加了r[10,63]。通过两种不同的模型分析r和φ的后验分布,结果与先前的结果相似,显示了数据的不确定性[64-65]。于是,2013年,RICE等[12]开始使用了基于贝叶斯统计的剩余产量模型对北太平洋大青鲨做了比较系统的评估,结果显示了北太平洋大青鲨处于一个相对稳定的状态,并没有出现过度捕捞,但是其结果的不确定性较大,建议必须谨慎地利用北太平洋大青鲨的资源。
7 不足与展望
目前,大青鲨资源评估的种群基础是标志方法。在太平洋海域,根据标志的结果可以将大青鲨划分成南北太平洋两个不同的种群,但是有些学者提出了太平洋大青鲨为单一种群的观点。可见,大青鲨种群现状并不清晰。根据海上观察员的观察结果显示,在赤道海域具有一定的大青鲨资源丰度。因此可以假设在赤道海域还存在着一个或者多个种群,它们可以在赤道海域彼此进行基因的交流,充当南北群体的交换媒介。未来可以在赤道海域附近做相关的标志性实验,进一步掌握赤道海域大青鲨种群结构现状。再者,新兴的分子标志技术在研究种群的遗传结构上具有一定的优越性,可以收集赤道海域样本,结合南北太平洋高纬度海域大青鲨样本,对整个太平洋的大青鲨种群结构做出科学的研究。
种群的结构是研究鱼类基础生物学的前提,不同的地理种群由于环境因素和饵料等因素的不同,其生长状况也不尽相同,繁殖能力也会存在显著差异。为此,在今后的研究中,可以根据样本收集的海域不同,对样本进行种群海域的划分,进行有差别的研究,这样更能全面地了解大青鲨的生长状况及繁殖现状。此外,由于捕捞强度的不同,导致捕捞群体的个体变小,因此,长时间序列的比较研究能较为充分地认识大青鲨的资源现状,为资源评估提供理论基础。大青鲨为维持自身在生态系统中的生存,可以导致其初次性成熟提前,影响其繁殖力。同样地可以针对历史时期的繁殖力统计,寻找其繁殖力是否发生显著改变,这将对大青鲨的资源评估产生重要的影响。
不同种群的生物学基础不尽相同,差异显著。大青鲨的生物学是进行资源评估的基础,所以,种群的结构及其相应的生物学研究是进一步提高大青鲨资源评估的有效手段。随着计算机计算能力的提高以及评估模型的发展,越来越多的模型被尝试着应用到资源评估中来。但是有效的评价手段是准确选择模型的基础,不同的模型应在合适的范围内使用。鉴于大青鲨历史数据的缺乏以及数据质量的不足,有目的的研究在现有数据缺乏的条件下,开发有效进行大青鲨资源评估的模型,对于提高大青鲨资源评估具有深远的意义。
[1] 孟庆闻,苏锦祥,缪学祖.鱼类分类学[M].北京:中国农业出版社,1995.
MENG QW,SU JX,LIAO X Z.Fishery Taxonomy[M].Beijing:Chinese Agricultural Press,1995.
[2] 朱元鼎,孟庆闻.中国动物志 (圆口纲 软骨鱼纲)[M].北京:科学出版社,2001.
ZHU Y D,MENG Q W.China fauna(Class Cyclostomata,Chondrichthyes)[M].Beijing:Science Press,2001.
[3] NAKANO H.Age,reproduction and migration of blue shark in the North Pacific Ocean[J].Bulletin of the National Research Institute of Far Seas Fisheries,1994(31):141-256.
[4] CLARKE S C,MCALLISTER M K,MILNERGULLAND E J,et al.Global estimates of shark catches using trade records from commercial markets[J].Ecology Letters,2006,9(10):1115-1126.
[5] CASTRO J I,WOODLEY C M,BRUDEK R L.A preliminary evaluation of the status of shark species[M].Roma:Food and Agriculture Organization of United Nations,1999.
[6] BONFIL R.Overview of world elasmobranch fisheries[M].Roma:Food and Agriculture Organization of United Nations,1994.
[7] KOHLER N E,TURNER P A,HOEY J J,et al.Tag and recapture data for three pelagic shark species:Blue shark(Prionace glauca),shortfin mako(Isurus oxyrinchus),and porbeagle(Iamna nasus)in the North Atlantic Ocean[J].The International Commission for the Conservation of Atlantic Tunas,2002,54(4):1231-1260.
[8] 戴小杰,许柳雄.大西洋中上层鲨鱼资源状况的初步探讨[J].水产学报,2003,27(4):328-333.
DAIX J,XU L X.Preliminary research on stock of pelagic sharks in the Atlantic Ocean[J].Journal of Fisheries of China,2003,27(4):328-333.
[9] 戴小杰,项忆军.热带大西洋公海金枪鱼延绳钓渔获物上钩率分析[J].水产学报,2000,24(1):81-85.
DAIX J,XIANG Y J.Analysis on catch rate of tuna longline fishing in the high sea of tropical Atlantic Ocean[J].Journal of Fisheries of China,2000,24(1):81-85.
[10] AIRES-DA-SILVA A M,GALLUCCIV F.Demographic and risk analyses applied to management and conservation of the blue shark(Prionace glauca)in the North Atlantic Ocean[J].Marine and Freshwater Research,2007,58(6):570-580.
[11] KLEIBER P,CLARKE S,BIGELOW K,et al.North Pacific blue shark stock assessment[M].Hawaii:National Oceanic and Atmospheric Administration,2009.
[12] RICE J,HARLEY S,MAUNDER M,et al.Stock assessment of blue shark in the North Pacific Ocean using stock synthesis[C].Scientific Committee Tenth Regular Session Western and Central Pacific Ocean Committee,Marshall Islands,2014.
[13] 詹秉义.渔业资源评估[M].北京:中国农业出版社,1995.
ZHAN B Y.Fishery stock assessment[M].Beijing:Chinese Agricultural Press,1995.
[14] 戴小杰,高春霞.鲨鱼类年龄和生长特性的研究进展[J].水产学报,2012,36(10):1624-1632.
DAIX J,GAO C X.Review on studies of age and growth of sharks[J].Journal of Fisheries of China,2012,36(10):1624-1632.
[15] LESSA R,SANTANA F M,HAZIN F H.Age and growth of the blue sharkPrionace glauca(linnaeus,1758)off northeastern Brazil[J].Fisheries Research,2004,66(1):19-30.
[16] VON BERTALANFFY L.A quantitative theory of organic growth(Inquires on growth laws.II)[J].Human Biology,1938,10(2):181-213.
[17] FABENS A J.Properties and fitting of the von Bertalanffy growth curve[J].Growth,1965,29(3):265.
[18] RICKER W E.Computation and interpretation of biological statistics of fish populations[M].Roma:Food and Agriculture Organization of United Nations,1987.
[19] MOLLET H F,EZCURRA JM,O’SULLIVAN JB.Captive biology of the pelagic stingray,Dasyatisviolacea(bonaparte,1832)[J].Marine and Freshwater Research,2002,53(2):531-541.
[20] RICKER W E.11 growth rates and models[J].Fish Physiology,1979(8):677-743.
[21] ROBERTSON T B.Chemical basis of growth and senescence[M].Roma:Food and Agriculture Organization of United Nations,1923.
[22] BARRETO R R,LESSA R P,HAZIN F H,et al.Age and growth of the blacknose shark,Carcharhinus acronotus(Poey,1860)off the northeastern Brazilian Coast[J].Fisheries Research,2011,110(1):170-176.
[23] SCHNUTE J.A versatile growth model with statistically stable parameters[J].Canadian Journal of Fisheries and Aquatic Sciences,1981,38(9):1128-1140.
[24] AASEN O.Length and growth of the porbeagle(Lamna nasus,Bonnaterre)in the North West Atlantic[J].Reports on Norwegian Fishery and Marine Investigations,1963:13(6):20-37.
[25] STEVENS J D.Vertebral rings as a means of age determination in the blue shark(Prionace glaucaL.)[J].Journal of the Marine Biological Association of the United Kingdom,1975,55(3):657-665.
[26] TANAKA S,MIZUE K.Studies on sharks,15:Age and growth of Japanese dogfishMustelus manazo bleekerin the East China Sea[J].Nippon Suisan Gakkaishi,1979,45(1):43-50.
[27] SKOMAL G B,NATANSON L J.Age and growth of the blue shark(Prionace glauca)in the North Atlantic Ocean[J].Fishery Bulletin-national Oceanic and Atmospheric Administration,2003,101(3):627-639.
[28] 高春霞,戴小杰,吴 峰,等.北大西洋大青鲨年龄和生长初步研究[J].上海海洋大学学报,2013,22(1):120-127.
GAO C X,DAI X J,WU F,et al.A preliminary study on age and growth of the blue shark(Prionace glauca)in the North Atlantic Ocean[J].Journal of Shanghai Ocean University,2013,22(1):120-127.
[29] CASEY JG,KOHLER N E.Long distance movements of Atlantic sharks from the NMFS cooperative shark tagging program[J].Underwater Naturalist,1991,19(3):87-91.
[30] CASEY J G.Transatlantic migrations of the blue shark:a case history of cooperative shark tagging[M].Fort Lauderdale:International Game Fish Association,1985.
[31] NAKANO H,STEVENS JD.The biology and ecology of the blue shark,Prionace glauca[M].New Jersey:Blackwell Publishing,2008.
[32] PRATT H L.Reproduction in the blue shark,Prionace glauca[J].Fisheries Bulletin,2001(77):445-470.
[33] 戴小杰,许柳雄.东太平洋热带海域大青鲨繁殖生物学特征[J].水产学报,2006,29(4):565-569.
DAI X J,XU L X.Reproductive biology characteristics ofPrionace glaucain the tropical Eastern Pacific Ocean[J].Journal of Fisheries of China,2006,29(4):565-569.
[34] 吴 峰,戴小杰,姜润林.热带中东大西洋海域大青鲨繁殖生物学研究[J].海洋湖沼通报,2012,3(6):29-36.
WU F,DAIX J,JIANG R L.Reproductive biology of blue shark in the tropical Eastern Central Atlantic Ocean[J].Transactions of Oceanology and Limnology,2012,3(6):29-36.
[35] LITVINOV F F,LAPTIKHOVSKY V V.Methods of investigations of shark heterodonty and dental formulae's variability with the blue shark,Prionaceglauca taken as an example[J].ICES CM,2005(27):15.
[36] TRICAST C,LEFEUVRE E M.Mating in the reef white-tip sharkTriaenodon obesus[J].Marine Biology,1985,84(3):233-237.
[37] NAKANO H,SHIMAZAKI K,MAKIHARA M.Distribution and biological characteristics of the blue shark(Prionace glauca)in the Central North Pacific[J].Bulletin of the Faculty of Fisheries-Hokkaido University,1985:36(3),99-113.
[38] CRUZ-RAMÍREZ A,SORIANO-VELÁSQUEZ S,SANTANA-HERNÁNDEZ H,et al.Aspectos reproductivos del tiburón azulPrionace glaucacapturado por la flota palangrera de mediana altura del Puerto de Manzanillo,Colima[J].Ciencia Pesquera,2012,20(1):39-48.
[39] FRANCIS M P,DUFFY C.Length at maturity in three pelagic sharks(Lamna nasus,Isurus Oxyrinchus,andPrionace glauca)from New Zealand[J].Fishery Bulletin,2005,103(3):489-500.
[40] GUBANOV Y P,GRIGOR'YEV V N.Observations on the distribution and biology of the blue sharkPrionace glauca(carcharhinidae)of the Indian Ocean[J].Voprosy Ikhtiologii,1975,15(1):37-43.
[41] JOLLY K A,SILVA C D,ATTWOOD C G.Age,growth and reproductive biology of the blue sharkPrionace glaucain South African waters[J].African Journal of Marine Science,2013,35(1):99-109.
[42] CAMPANA S E,MARKS L,JOYCE W,et al.Effects of recreational and commercial fishing on blue sharks(Prionace glauca)in Atlantic Canada,with inferences on the North Atlantic population[J].Canadian Journal of Fisheries and Aquatic Sciences,2006,63(3):670-682.
[43] STEVENS J D.The occurrence and significance of tooth cuts on the blue shark(Prionace glaucaL.)from British waters[J].Journal of the Marine Biological Association of the United Kingdom,1974,54(2):373-378.
[44] MEGALOFONOU P,DAMALAS D,DE METRIO G.Biological characteristics of blue shark,Prionace glauca,in the Mediterranean Sea[J].Journal of the Marine Biological Association of the United Kingdom,2009,89(6):1233-1242.
[45] MONTEALEGRE-QUIJANO S,CARDOSO A T C,SILVA R Z,et al.Sexual development,size at maturity,size at maternity and fecundity of the blue shark Prionace glauca(linnaeus,1758)in theSouthwest Atlantic[J].Fisheries Research,2014,160(3):18-32.
[46] CARRERA-FERNÁNDEZ M,GALVÁN-MAGAÑA F,CEBALLOS-VÁZQUEZ P.Reproductive biology of the blue shark Prionace glauca(Chondrichthyes:Carcharhinidae)off Baja California Sur,México[J].Aqua,2010,16(3):101-110.
[47] SIPPEL T,WRAITH J,KOHIN S,et al.A summary of blue shark(Prionace glauca)and shortfin mako shark(Isurus oxyrinchus)tagging data available from the North and Southwest Pacific Ocean[C].California:Working Document Submitted to the ISC Shark Working Group Workshop,La Jolla,2011.
[48] TAVARES R,ORTIZ M,AROCHA F.Population structure,distribution and relative abundance of the blue shark(Prionace glauca)in the Caribbean Sea and adjacent waters of the North Atlantic[J].Fisheries Research,2012,129(2):137-152.
[49] KOHLER N E,TURNER PA.Stock structure of the blue shark(Prionace glauca)in the North Atlantic Ocean based on tagging data[M].New Jersey:Blackwell Publishing,2008.
[50] FITZMAURICE P,GREEN P,KEIRSE G,et al.Stock discrimination of the blue shark,based on Irish tagging data[J].Inter-American Tropical Tuna Commission Bulletin,2005,58(3):1171-1178.
[51] KUBODERA T,WATANABE H,ICHIIT.Feeding habits of the blue shark,Prionace glauca,and salmon shark,Lamna ditropis,in the transition region of the Western North Pacific[J].Reviews in Fish Biology and Fisheries,2007,17(2):111-124.
[52] MANDELMAN JW,COOPER PW,WERNER T B,et al.Shark bycatch and depredation in the US Atlantic pelagic longline fishery[J].Reviews in Fish Biology and Fisheries,2008,18(4):427-442.
[53] 郑真真,许强华,戴小杰,等.基于线粒体控制区部分序列的大青鲨种群遗传结构研究[J].大连海洋大学学报,2014,9(5):463-468.
ZHENG Z Z,XU Q H,DAIX J,et al.Population genetic structure of blue sharkPrionace glaucabased on partial control region sequence in mitochondrial DNA[J].Journal of Dalian Ocean University,2014,9(5):463-468.
[54] TAGUCHI M,KING J R,WETKLO M,et al.Population genetic structure and demographic history of pacific blue sharks(Prionace glauca)inferred from mitochondrial DNA analysis[J].Marine and Freshwater Research,2015,66(3):267-275.
[55] NAKANO H.Updated standardizedCPUEfor pelagic sharks caught by the Japanese longline fishery in the Atlantic Ocean[J].Inter-American Tropical Tuna Commission Bulletin,1999,51(6):1796-1803.
[56] MATSUNAGA H.StandardizedCPUEfor shortfin mako caught by the Japanese tuna longline fishery in the Atlantic Ocean,1994-2005[J].Inter-American Tropical Tuna Commission Bulletin,2008,62(5):1581-1586.
[57] CARVALHO F,AHRENS R,MURIE D,et al.Incorporating specific change points in catchability in fisheries stock assessment models:an alternative approach applied to the blue shark(Prionace glauca)stock in the South Atlantic Ocean[J].Fisheries Research,2014,154(4):135-146.
[58] MAUNDER M N.Is it time to discard the Schaefer Model from the stock assessment scientist’s toolbox?[J].Fisheries Research,2003,61(1):145-149.
[59] PELLA J J,TOMLINSON P K.A generalized stock production model[J].Inter-American Tropical Tuna Commission Bulletin,1969,13(3):416-497.
[60] PRAGER M H.Comparison of logistic and generalized Surplus-production Models applied to swordfish,Xiphias gladius,in the North Atlantic Ocean[J].Fisheries Research,2002,58(1):41-57.
[61] ANON N.Report of the 2008 shark stock assessment meeting[R].Madrid:Inter-American Tropical Tuna Commission,2009:1343-1491.
[62] CORTÉSE,AROCHA F,BEERKIRCHER L,etal.Ecological risk assessment of pelagic sharks caught in Atlantic pelagic longline fisheries[J].Aquatic Living Resources,2010,23(1):25-34.
[63] CORTÉS E.Incorporating uncertainty into demographic modeling:application to shark populations and their conservation[J].Conservation Biology,2002,16(4):1048-1062.
[64] MEJUTO J,GARCÍA-CORTÉSB.Reproductive and distribution parameters of the blue sharkPrionace glauca,on the basis of on-board observations at sea in the Atlantic,Indian and Pacific Oceans[J].The International Commission for the Conservation of Atlantic Tunas,2005,58(3):974-1000.
[65] MCALLISTER M K,KIRKWOOD G P.Using bayesian decision analysis to help achieve a precautionary approach for managing developingfisheries[J].Canadian Journal of Fisheries and Aquatic Sciences,1998,55(12):2642-2661.
Research progress of blue shark Prionace glauca fishery biology
LI Wei-wen1,TIAN Si-quan1,2,3,4,DAI Xiao-jie1,2,3,4,CHEN Xin-jun1,2,3,4
(1.Collge of Marine Sciences,Shanghai Ocean University,Shanghai 201306,China;2.National Distant Water Fisheries Engineering Research Center,Shanghai Ocean University,Shanghai201306,China;3.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Minister of Education,Shanghai Ocean University,Shanghai201306,China;4.Scientific Observing and Experimental Station of Oceanic Fishery Resources,Ministry of Agriculture,Shanghai Ocean University,Shanghai201306,China)
Blue shark(Prionace glauca)is widely distributed in the south and north tropical and temperate waters around the world and commonly caught by longline fisheries and gill-net fisheries.Although it is relatively productive,there is global concern about the impact of depletion and possible loss of apex predator fishes in marine ecosystems.In addition,it is of particular relevance in relation to sharks as their biological characteristics make them particularly vulnerable to over-exploitation.So the Regional Fisheries Management Organizations(RFMOs)started to assess the status of blue shark in three oceans.As the fishing intensity increases blue shark resource has been declining.The stock assessment of blue shark could not be recognized by public due to its uncertainty of basic data.Based on document quantitative analysis,this study briefly introduced the fishery biological research history of blue shark,and the focus themes of different periods were acknowledged.Multiplies references on age and growth,reproductive biology,population structure,abundance index and stock assessment were introduced and analyzed.Finally suggestions were put forward for future study to improve the credibility of stock assessment and to improve the understanding of blue shark resources situation,providing a scientific basis for the maintenance of blue shark fisheries.
blue shark(Prionace glauca);fishery biology;abundance index;stock assessment models
S 931
A
1004-2490(2016)05-0540-11
2015-09-18
区域国际组织观察员计划(D8002-14-8014A-2);远洋渔业协同创新中心(B2-5004-13-0101)
李伟文(1988-),男,福建漳州人,博士研究生,研究方向为渔业资源评估及保护生物学。
E-mail:liweiwen0207@163.com
戴小杰,教授。E-mail:xjdai@shou.edu.cn
