毕节自育特色烤烟品种烟叶衰老期的氮代谢特征
2016-03-01李晓睿武云杰侯冰清张景华张小全杨铁钊
戚 莹, 李晓睿, 武云杰, 侯冰清, 张景华,张小全, 翟 欣, 杨铁钊*
(1.河南农业大学 烟草学院, 河南 郑州 450002; 2.贵州省烟草公司毕节地区公司, 贵州 毕节 551700)
毕节自育特色烤烟品种烟叶衰老期的氮代谢特征
戚 莹1, 李晓睿1, 武云杰1, 侯冰清1, 张景华1,张小全1, 翟 欣2, 杨铁钊1*
(1.河南农业大学 烟草学院, 河南 郑州 450002; 2.贵州省烟草公司毕节地区公司, 贵州 毕节 551700)
为毕节特色优质烤烟生产上的合理施肥提供理论依据,通过设置不同施氮量处理,研究氮素营养对3个毕节自育特色烤烟品种氮代谢相关指标的影响。结果表明:随着施氮量的增加,烟叶成熟过程中质体色素、可溶性蛋白和总氮积累量增大,谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)、硝酸还原酶活性(NR)增强,氨气补偿点升高,烟叶氮代谢强度增强。品种间差异显著,毕纳1号的氮素营养物质降解速率较快,其GS活性和NR活性较低,GDH活性较高,氨气补偿点较高,耐肥性较强;黔西1号耐肥性较弱,韭菜坪2号居中。烟叶的氮素运筹是氮素吸收、同化和在再转移能力的表现,其中GS活性起主导作用。毕纳1号、韭菜坪2号和黔西1号的氮代谢相关指标均以6 kg/667m2施氮量处理氮代谢较为协调,以毕纳1号耐肥性最强,适合6~7.5 kg/667m2肥力条件,黔西1号耐肥性较弱,适合4.5~6 kg/667m2的肥力条件,韭菜坪2号居中,适合6 kg/667m2的肥力条件。
烤烟; 氮代谢; 品种; 施氮量; 毕节
品种是农业生产的基础,而优良的品种则是农业生产的重要条件。烤烟品种对产量的贡献率为20%~35%,对品质的贡献率高达50%,在烟草生产中具有重要作用[1-3]。烟草品种具有较广泛的适应性,但只有种植在适宜的生态条件下,才能获得最佳的产量和品质。毕节地区是我国第三大烤烟产区,其烤烟种植区域分布在海拔800~2 300 m,海拔跨度大,立体气候特征明显,土壤类型较多[4-6]。根据品种对生态、栽培的响应,选择适合当地的特色烤烟品种,充分彰显毕节烟叶清甜香风格,以提高烟叶品质的适配性[7-8]。氮素是影响烟叶产量和品质的重要营养元素,不同基因型烤烟具有不同的氮素代谢特性[9-10]。特别是在烟叶衰老期,氮素的运筹和转化对烟叶成熟落黄至关重要[11-12]。氮素供应过多,烟叶产量提高但成熟期推迟,特别是氮化合物含量过高,造成品质下降;氮素供应过少,则烟叶发育不良,对产量和品质均不利[13]。不同品种的氮素代谢(氮素吸收和转化)存在很大差异,并最终影响烟叶的产量和品质[14]。由于不同地区的土壤特性差异,关于毕节地区烟叶特色烤烟品种氮代谢方面的研究报道极少。为此,通过不同施氮量处理,研究3个毕节特色烤烟品种的氮代谢特性,以期为当地特色优质烤烟生产上的合理施肥提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 烤烟品种 毕纳1号、韭菜坪2号和黔西1号,毕节地区自育优质烤烟品种。
1.1.2 肥料 硝酸铵、普钙和硫酸钾,由毕节市烟草公司提供。
1.2 试验时间及试验地概况
试验于2014年在毕节七星关区进行。试验地前茬作物为烟草,土壤肥力均匀,供试土壤为黄棕壤,基础肥力为全氮1.24 g/kg,碱解氮29.98 mg/kg,速效磷20.21 mg/kg,速效钾126.15 mg/kg,有机质33.87 g/kg,pH6.78。
1.3 试验设计
采用随机区组设计,施氮量设置低氮、中氮和高氮3个处理,分别为施用纯氮4.5 kg/667m2、6 kg/667m2和7.5 kg/667m2,磷钾肥均为P2O56 kg/667m2,K2O 15 kg/667m2。每个处理3次重复,共27个小区。每个小区66.7 m2,株行距为55 cm×110 cm。按照大田优质烟叶生产管理方法进行管理。选取各品种整齐一致的单株,以自下向上第11片叶为试验对象,在叶龄为30 d、40 d、50 d、60 d和调制前1 d(自幼叶长1 cm,宽0.5 cm时作为第1天叶龄)取样,各指标测定3次重复。
1.4 指标测定
谷氨酰胺合成酶(GS)活性参照文献[16]的方法测定,总GS活性计算以1 mg/min粗蛋白催化产生的γ-谷氨酰异羟肟酸μmol数表示。
谷氨酸脱氢酶(GDH)活性参照文献[17]的方法测定,总GDH活性计算以1 mg/min粗蛋白催化NADH减少的μmol数表示。在340 nm处测定30 s内吸光值的变化。
χs=Г×KH×Kd
1.5 数据处理
采用MicrosoftExcel 2013和SPSS 22.0对数据进行分析。
2 结果与分析
2.1 施氮量对烟叶质体色素含量的影响
质体色素是烟叶香气成分的主要香气前体物,其含量对氮素营养十分敏感,并影响烟叶的外观和内在品质[19]。由表1可知,叶绿素含量从叶龄为30 d或40 d时开始逐渐下降,烟叶衰老过程中,前期降幅较小,后期随着烟叶的成熟降幅增大。不同品种在同一处理下毕纳1号最低,黔西1号最高,但品种间差异不显著。随着施氮量的增加叶绿素含量显著提高,特别是高氮处理下叶绿素的积累峰值推迟10 d。类胡萝卜素的含量变化与叶绿素一致,但含量和降幅均小于叶绿素。品种间的最大积累量差异不明显,从峰值到调制前1 d的降幅以毕纳1号最大,黔西1号和韭菜坪2号较小。施氮量增大时,类胡萝卜素积累量增加,在高氮条件下峰值推迟到叶龄为40 d,但与叶绿素不同,在中氮处理下类胡萝卜素积累量最高。说明,类胡萝卜素存在品种与栽培的互作。
表1 不同施氮量烟叶衰老期叶绿素及类胡萝卜素的含量
Table 1 Leaf chlorophyll and carotenoid content of different tobacco varieties at leaf decrepitude period under different nitrogen application rate

处理Treatments品种Variety叶绿素含量/(mg/g)Chlorophyll30∗d40∗d50∗d60∗d1∗∗d类胡萝卜素含量/(mg/g)Carotenoid30∗d40∗d50∗d60∗d1∗∗d低氮毕纳1号3.21e2.54d1.90e1.51e1.20c0.80ab0.67c0.49d0.43c0.36c Lownitrogen韭菜坪2号3.32de2.53d2.02e1.58e1.16c0.80ab0.74b0.70b0.56b0.42bc黔西1号3.42d2.80c2.20e1.82d1.36bc0.76b0.75b0.60c0.53b0.47bc中氮毕纳1号3.74c3.08c2.30de1.82d1.24c0.92a0.72b0.57c0.43c0.32c Middlenitrogen韭菜坪2号3.55cd3.18c2.38d1.96d1.29c0.92a0.85a0.74ab0.61ab0.53b黔西1号3.83c3.57b3.12a2.57c1.91a0.84ab0.77b0.69b0.57b0.46bc高氮毕纳1号4.17b4.65a4.20b3.22b1.49b0.83ab0.87a0.81a0.71a0.63a Highnitrogen韭菜坪2号4.35ab4.84a4.46ab3.37ab1.75ab0.70b0.82a0.77a0.72a0.60a黔西1号4.58a4.75a4.56a3.46a1.95a0.77ab0.82a0.76a0.68a0.63a
注:不同字母表示在5%水平上差异显著,表头中有*的数字为叶龄,** 的数字为调制前的叶龄(下同)。
Note: Different letters mean significance of difference at 5% level. The figures with * and ** represent leaf age and leaf age before curing respectively.The same below.
2.2 施氮量对烟叶可溶性蛋白和总氮含量的影响
可溶性蛋白主要成分为1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco),特定叶龄的总氮和可溶性蛋白含量可以很好地反映作物氮素营养状况[20]。由表2可知,可溶性蛋白含量,在烟叶成熟过程中,可溶性蛋白均在叶龄50 d达峰值之后一直呈下降趋势,同一施氮量品种间差异显著,以黔西1号最高;不同施氮量处理,可溶性蛋白含量随施氮量的增加而升高,并在调制前含量最高。总氮含量,从叶龄30 d或40 d达峰值后下降,均以高氮处理峰值最高,而且中氮和高氮处理在叶龄40 d时达最大值。随着施氮量的增加,总氮最大积累量升高,且调制前的总氮含量也较高,从最大值到调制前的降解比例减少,烟叶衰老速度减慢。
表2 不同施氮量烟叶衰老期的可溶性蛋白及总氮含量
Table 2 Leaf soluble protein and total nitrogen content of different tobacco varieties at leaf decrepitude period under different nitrogen application rate

处理Treatments品种Variety可溶性蛋白含量/(mg/g)Solubleprotein30∗d40∗d50∗d60∗d1∗∗d总氮含量/%Totalnitrogen30∗d40∗d50∗d60∗d1∗∗d低氮毕纳1号4.29cd4.38e3.50e3.00e3.11d2.47c2.38d1.78d1.55de0.82d Lownitrogen韭菜坪2号4.42c4.59e4.42d3.85d3.12d2.74bc2.46d2.04c1.21e0.91d黔西1号4.41c5.90d5.65c4.22c2.63e2.91b2.81c2.22c1.73d0.91d中氮毕纳1号5.17d6.67c6.00b4.34c3.47c2.51c2.96c2.19c1.90cd1.75c Middlenitrogen韭菜坪2号5.42c6.16d5.64c4.95b3.12d2.68bc2.82c2.69b2.24c1.94b黔西1号5.77c7.00b5.59cc5.24a4.08b2.99b3.19bc2.44b2.40bc1.97b高氮毕纳1号6.20b7.51a5.90b5.40a4.07b3.12ab3.36b2.97a2.28c2.04b Highnitrogen韭菜坪2号6.32ab7.52a5.79bc5.08ab4.26a3.34a3.88a3.01a2.95a2.25a黔西1号6.41a7.57a6.49a5.12ab4.29a3.32a3.78a3.00a2.53b2.05b

Table 3 Leaf concentration of different tobacco varieties at leaf decrepitude period under different nitrogen application rate mmol/L

2.4 施氮量对烟叶衰老期谷氨酰胺合成酶、谷氨酸脱氢酶和硝酸还原酶活性的影响
2.4.2 谷氨酸脱氢酶(GDH) GDH具有合成氨和脱氨功能,是氮代谢的调节酶,烟叶衰老期的主要作用是脱氨[24-25]。从表4还看出,同一施氮处理下,GDH活性均在叶龄40 d时达峰值,低氮和中氮处理叶龄40 d以前GDH活性增幅较小;叶龄40~50 d后开始小幅下降,叶龄50 d至调制前1 d活性大幅下降。随着施氮量的升高,GDH活性大幅升高,在叶龄40 d后处理间差异显著。低氮和中氮处理,在不同品种间以毕纳1号GDH活性较高,韭菜坪2号和黔西1号差异不显著;高氮处理,毕纳1号显著高于韭菜坪2号,黔西1号最低。说明,施氮量增大,GDH活性对铵的转运作用增强,在后期氮素的运筹调节中起主要作用,品种间也存在差异。
表4 不同施氮量烟叶衰老期谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)和硝酸还原酶(NR)的活性
Table 4 Leaf GS, GDH and NR activity of different tobacco varieties at leaf decrepitude period under different nitrogen application rate
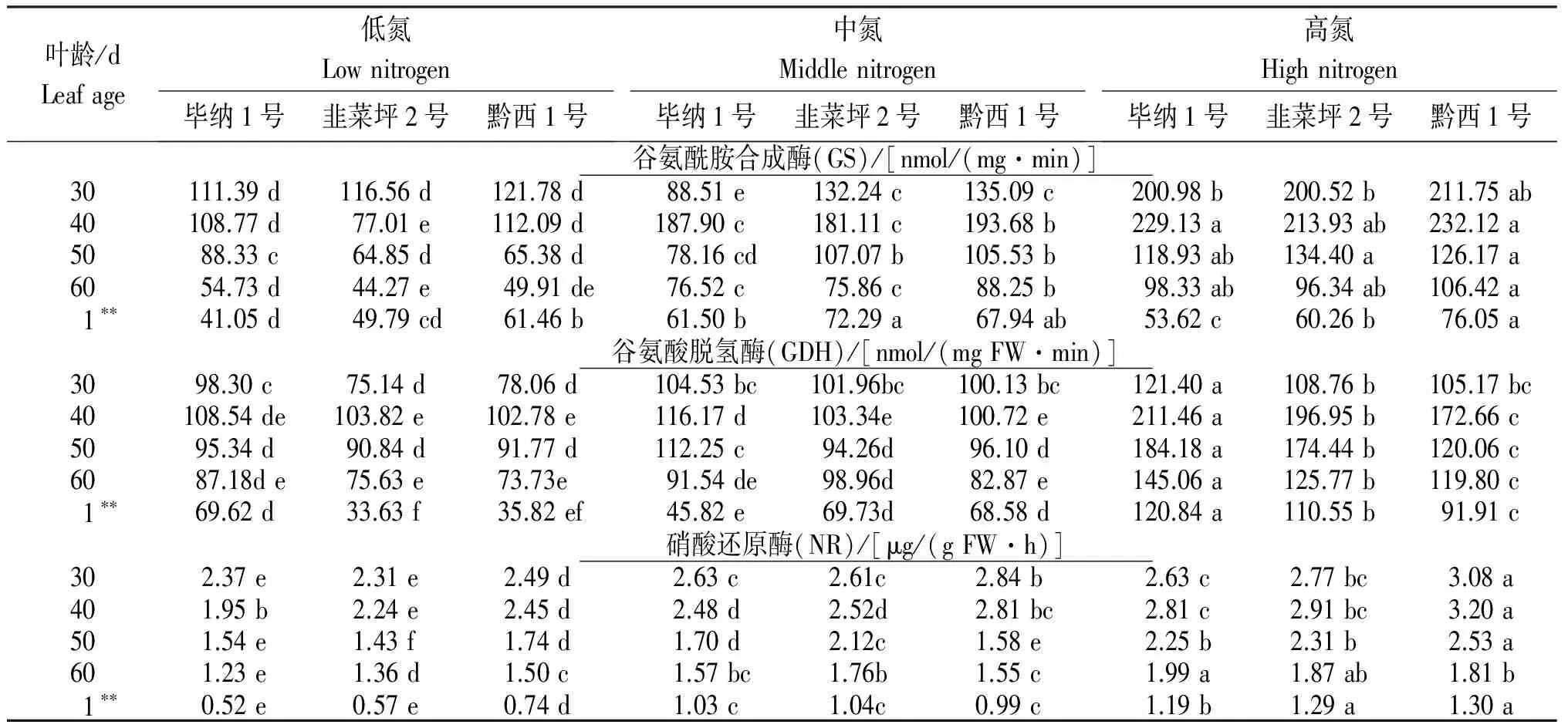
叶龄/dLeafage低氮Lownitrogen毕纳1号韭菜坪2号黔西1号中氮Middlenitrogen毕纳1号韭菜坪2号黔西1号高氮Highnitrogen毕纳1号韭菜坪2号黔西1号谷氨酰胺合成酶(GS)/[nmol/(mg·min)]30111.39d116.56d121.78d88.51e132.24c135.09c200.98b200.52b211.75ab40108.77d77.01e112.09d187.90c181.11c193.68b229.13a213.93ab232.12a5088.33c64.85d65.38d78.16cd107.07b105.53b118.93ab134.40a126.17a6054.73d44.27e49.91de76.52c75.86c88.25b98.33ab96.34ab106.42a1∗∗41.05d49.79cd61.46b61.50b72.29a67.94ab53.62c60.26b76.05a谷氨酸脱氢酶(GDH)/[nmol/(mgFW·min)]3098.30c75.14d78.06d104.53bc101.96bc100.13bc121.40a108.76b105.17bc40108.54de103.82e102.78e116.17d103.34e100.72e211.46a196.95b172.66c5095.34d90.84d91.77d112.25c94.26d96.10d184.18a174.44b120.06c6087.18de75.63e73.73e91.54de98.96d82.87e145.06a125.77b119.80c1∗∗69.62d33.63f35.82ef45.82e69.73d68.58d120.84a110.55b91.91c硝酸还原酶(NR)/[μg/(gFW·h)]302.37e2.31e2.49d2.63c2.61c2.84b2.63c2.77bc3.08a401.95b2.24e2.45d2.48d2.52d2.81bc2.81c2.91bc3.20a501.54e1.43f1.74d1.70d2.12c1.58e2.25b2.31b2.53a601.23e1.36d1.50c1.57bc1.76b1.55c1.99a1.87ab1.81b1∗∗0.52e0.57e0.74d1.03c1.04c0.99c1.19b1.29a1.30a
2.4.3 硝酸还原酶(NR) NR是植物吸收和同化土壤中氮素的关键酶[26]。由表4可知,同一施氮处理下,随着叶片的衰老,NR活性逐渐下降,并随施氮量的增加在叶龄30 d或40 d达峰值后下降,降幅较为平稳,只在叶龄60 d至调制前1 d呈大幅下降。品种间以黔西1号NR活性最高,毕纳1号和韭菜坪2号差异不大。随着施氮量的增大,NR活性增强,高氮处理下峰值推迟至叶龄40 d,至调制前1 d NR活性仍较高。说明,随着施氮量的增加,烟叶对氮素的吸收能力明显提高,烟叶衰老推迟。
2.5 施氮量对烟叶衰老期质外体铵浓度、pH和氨气补偿点的影响
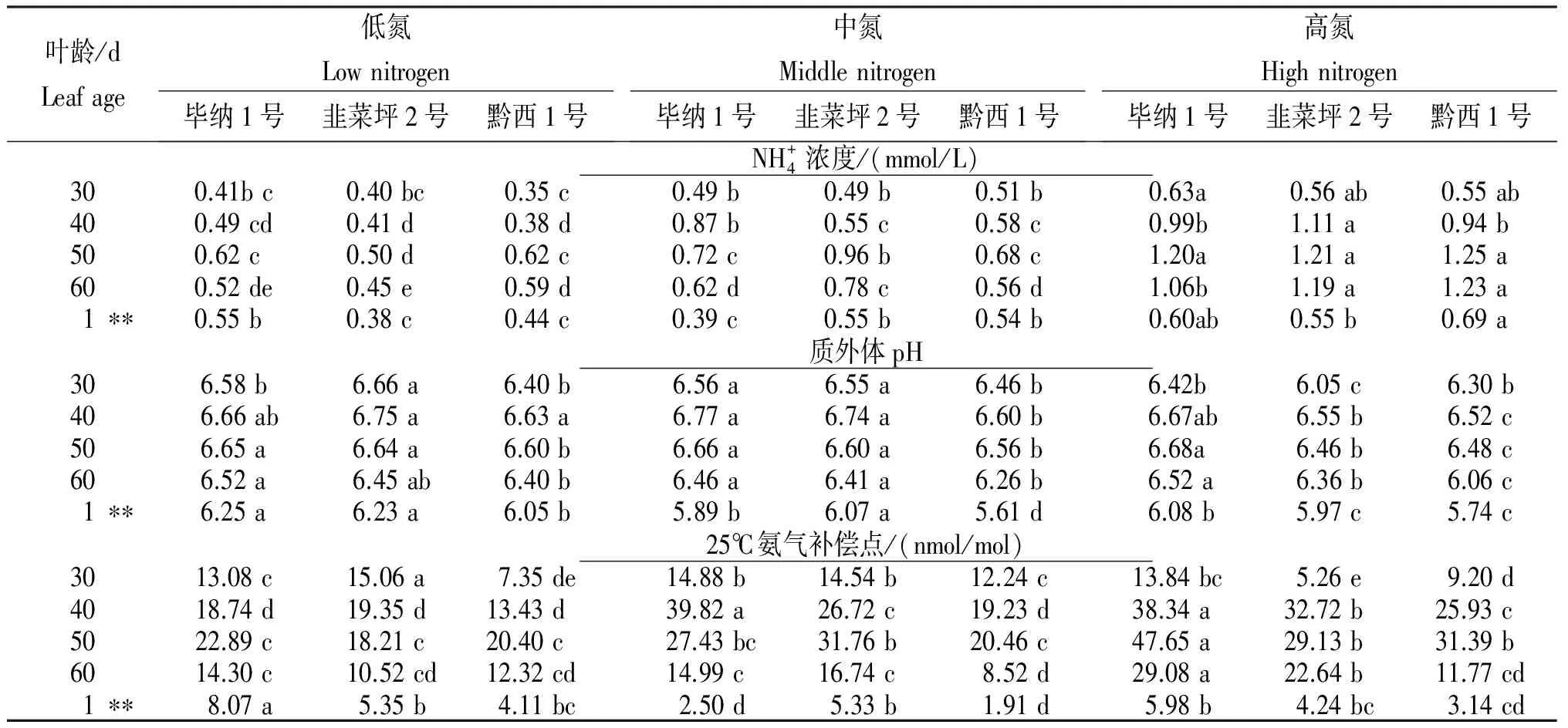
叶龄/dLeafage低氮Lownitrogen毕纳1号韭菜坪2号黔西1号中氮Middlenitrogen毕纳1号韭菜坪2号黔西1号高氮Highnitrogen毕纳1号韭菜坪2号黔西1号NH+4浓度/(mmol/L)300.41bc0.40bc0.35c0.49b0.49b0.51b0.63a0.56ab0.55ab400.49cd0.41d0.38d0.87b0.55c0.58c0.99b1.11a0.94b500.62c0.50d0.62c0.72c0.96b0.68c1.20a1.21a1.25a600.52de0.45e0.59d0.62d0.78c0.56d1.06b1.19a1.23a1∗∗0.55b0.38c0.44c0.39c0.55b0.54b0.60ab0.55b0.69a质外体pH306.58b6.66a6.40b6.56a6.55a6.46b6.42b6.05c6.30b406.66ab6.75a6.63a6.77a6.74a6.60b6.67ab6.55b6.52c506.65a6.64a6.60b6.66a6.60a6.56b6.68a6.46b6.48c606.52a6.45ab6.40b6.46a6.41a6.26b6.52a6.36b6.06c1∗∗6.25a6.23a6.05b5.89b6.07a5.61d6.08b5.97c5.74c25℃氨气补偿点/(nmol/mol)3013.08c15.06a7.35de14.88b14.54b12.24c13.84bc5.26e9.20d4018.74d19.35d13.43d39.82a26.72c19.23d38.34a32.72b25.93c5022.89c18.21c20.40c27.43bc31.76b20.46c47.65a29.13b31.39b6014.30c10.52cd12.32cd14.99c16.74c8.52d29.08a22.64b11.77cd1∗∗8.07a5.35b4.11bc2.50d5.33b1.91d5.98b4.24bc3.14cd
3 结论与讨论
3) 3个品种间氮代谢存在差异,毕纳1号的氮素营养物质降解相对较快,GS和NR活性较低,GDH活性较高,在高氮条件下也呈相同的变化趋势,具有相对较强的耐肥性,应保证相对较高的肥力条件。黔西1号则耐肥性相对较弱,特别是NR活性显著高于其他2个品种,GS活性也较高。因此,应在中氮处理下适当减少施肥量种植。
[1] 唐远驹.烟叶风格特色定位[J].中国烟草科学,2008,29(3):1-5.
[2] 王彦亭.我国烟草育种工作发展思路[J].中国烟草科学,2001,23(4):1-5.
[3] 贾兴华.烟草新品种及丰产栽培技术[M].北京:中国劳动社会保障出版社,2001.
[4] 代昌明,腊贵晓,翟 欣,等.毕节地区植烟土壤肥力状况评价[J].江西农业学报,2011,23(8):9-11,16.
[5] 牛路路,陈 雪,翟 欣,等.毕节不同海拔条件下烤烟化学成分分析[J].山西农业科学,2013,41(5):446-448.
[6] 符云鹏,王小翠,陈 雪,等.毕节烟区土壤pH值分布状况及与土壤养分的关系[J].土壤,2013,45(1):46-51.
[7] 王小翠,喻奇伟,符云鹏,等.毕节烟区烤烟化学成分、感官质量及其相关性研究[J].河南农业科学,2012,41(6):58-61,64.
[8] 石俊雄,陈 雪,雷 璐,等.生态因子对贵州烟叶主要化学成分的影响[J].中国烟草科学,2008,29(2):18-22.
[9] 陈顺辉,李文卿,江荣风,等.施氮量对烤烟产量和品质的影响[J].中国烟草学报,2003(Z1):36-40.
[10] 武云杰,张小全,段旺军,等.不同氮素利用效率基因型烤烟叶片衰老期间氮素代谢差异研究[J].中国烟草学报,2012,18(5):23-28.
[11] 武云杰,李 飞,杨铁钊,等.氮素营养水平对衰老期烟叶氮代谢的影响及品种间差异[J].中国烟草学报,2014(4):41-47.
[12] 武云杰,张小全,段旺军,等.烤烟叶片衰老期氨气挥发特征及其生理调控研究[J].西北植物学报,2012,32(10):2082-2088.
[13] 陈爱国,王树声,申国明,等.烤烟叶片成熟期间碳氮代谢主要物质流分析[J].中国烟草学报,2010,16(4):30-34.
[14] 武云杰,杨铁钊,张小全,等.不同烤烟品种烟叶衰老期氨气挥发及其与氮素代谢的相关性[J].中国农业科学,2013,46(19):4027-4034.
[15] 邹 琦.植物生理学实验指导[M].北京:中国农业出版社,2000.
[16] O’Neal D,Joy K W.Glutamine synthetase of pea leaves.I.Purification, stabilization, and pH optima[J].Archives of Biochemistry and Biophysics,1973,159:113-122.
[17] Turano F J,Dashner R,Upadhyaya A,et al.Purification of mitochondrial glutamate dehydrogenase from dark-grown soybean seedlings[J].Plant Physiology,1996,112:1357-1364.
[18] 段旺军,杨铁钊,刘化冰,等.烟叶氨气补偿点的品种间差异及其与氮素代谢的关系研究[J].植物营养与肥料学报,2011,17(2):419-424.
[19] Week W W.Chemistry of tobacco constituents influencing flavor and aroma[J].RecentAdvance in Tobacco Science,1985,11:175-200.
[20] Biswas A K,Choudhuri M A Mechanism of monocarpic senescence in rice[J].Plant Physiol,1980,65:340-345.
[21] Joy K W.Ammonia,glutamine and asparagine:a carbon-nitrogen interface[J].Can.J.Bot.,1988,66:2103-2109.
[22] Masclaux C,Valadier M H,Brugi re N,et al.Characterization of the sink/source transition in tobacco (Nicotiana tabacum L.)shoots in relation to nitrogen management and leaf senescence[J].Planta,2000,211:510-518.
[23] Brugiere N,Dubois F,Limami A,et al.Glutamine synthetase in the phloem plays a major role in controlling proline production[J].Plant Cell,1999,11:1995-2011.
[24] Miflin B J, Habash D Z.The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crops[J].Journal of Experimental Botany,2002,53(370):979-987.
[25] Skopelitis D S,Paranychianakis N V,Kouvarakis A,et al.The isoenzyme 7 of tobacco NAD(H)-dependent glutamate dehydrogenase exhibits high deaminating and low aminating activities in vivo[J].Plant Physiology,2007,145:1726-1734.
[26] 刘 丽,甘志军,王宪泽.植物氮代谢硝酸还原酶水平调控机制的研究进展[J].西北植物学报,2004,24(7):1355-1361.
[27] Dubois F,Brugiere N,Sangwan R S,et al.Localization of tobacco cytosolic glutamine synthetase and the corresponding transcrips shows organ-and cell-specific patterns of protein synthesis and gene expression[J].Plant Molecular Biology,1996,31:803-817.
[28] Norman R J, Guindo D, Wells B R,et al.Seasonal Accumulation and Partitioning of Nitrogen-15 in Rice[J].Soil Science Society of America Journal,1992,56(5):1521-1527.
[29] Schjoerring J K,Husted S,M ck G,et al.The regulation of ammonium translocation in plants[J].J.Exp.Bot.,2002,53:883-890.
[30] Purnell M P,Botella,J R.Tobacco isoenzyme 1 of NAD(H)-dependent glutamate dehydrogenase catabolizes glutamate in vivo[J].Plant Physiology,2007,143(1):530-539.
[31] Frechilla S,Lasa B, Aleu M,et al.Short-term ammonium supply stimulates glutamate dehydrogenase activity and alternative pathway respiration in roots of pea plants[J].Journal of Plant Physiology,2002,159:811-818.
[32] Mattsson M,Schjoerring J K.Senescence-induced changes in apoplastic and bulk tissue ammonia concentrations of ryegrass leaves[J].New Phytologist,2003,160(11):489-499.
[33] 吴小庆,徐阳春,沈其荣.植物叶片氨挥发研究进展[J].生态与农村环境学报,2006,22(2):80-84.
(责任编辑: 杨 林)
Nitrogen Metabolism Characteristics of Bijie Specific Tobacco Leaves at Leaf Decrepitude Period
QI Ying1, LI Xiaorui, WU Yunjie1, HOU Bingqing1, ZHANG Jinghua1, ZHANG Xiaoquan1, ZHAI Xin2, YANG Tiezhao1*
(1.CollegeofTobaccoScience,HenanAgriculturalUniversity,Zhengzhou,Henan450002;2.BijieTobaccoCompany,GuizhouTobaccoCompany,Bijie,Guizhou551700,China)
The randomized block design was used to study the effect of different nitrogen application amount on the related indexes of nitrogen metabolism in three specific tobacco varieties bred by Bijie and to provide the theoretical basis for rational application of specific quality tobacco varieties in Bijie. Results: The accumulation amount of plamochromic pigment, soluble protein and total nitrogen in tobacco leaves, leaf GS, GDH and NR activity, leaf ammonia gas compensation point and leaf nitrogen metabolism increase with increase of nitrogen application amount during the tobacco leaf maturation process but there are significant differences among three tobacco varieties. Bina 1 has the characteristics of rapid nitrogen nutrient matter degradation rate, lower GS and Nr activity, high GDH activity, higher ammonia gas compensation point. The fertilizer tolerance of Bina 1 is the strongest, followed by Jiucaiping 2 and Qianxi 1. The nitrogen management of tobacco leaves is performance of nitrogen absorption, assimilation and re-transfer ability and GS activity plays the leading role in nitrogen absorption, assimilation and re-transfers ability of tobacco leaves. The optimum nitrogen metabolism relative index for Bina 1, Jiucaiping 2 and Qianxi 1 is under 6 kg/667m2nitrogen application amount. Bina 1 with strong fertilizer tolerance, Jiucaiping 2 with moderate fertilizer tolerance and Qianxi 1 with weak fertilizer tolerance are suitable to 6~7.5 kg/667m2, 6 kg/667m2and 4.5~6 kg/667m2nitrogen application amount respectively.
tobacco; nitrogen metabolism; variety; nitrogen application rate; Bijie
2015-09-09; 2016-03-03修回
贵州省烟草公司科技攻关项目“毕节特色烤烟品种的深度挖掘与配套技术研究”(2010-16)
戚 莹(1990-)女,在读硕士,研究方向:烟草遗传育种与品质改良。E-mail:hnndqiying@163.com
*通讯作者:杨铁钊(1956-),男,教授,博士生导师,从事烟草遗传育种与品质改良研究。E-mail:yangtiezhao@126.com
1001-3601(2016)03-0113-0059-06
S572.062
A
