祁连山东段不同放牧强度下高原鼢鼠栖息地选择分析
2016-02-23楚彬花立民周延山刘丽李帅
楚彬,花立民,周延山,刘丽,李帅
(甘肃农业大学草业学院,甘肃 兰州 730070)
祁连山东段不同放牧强度下高原鼢鼠栖息地选择分析
楚彬,花立民*,周延山,刘丽,李帅
(甘肃农业大学草业学院,甘肃 兰州 730070)
摘要:本研究以祁连山东段高寒草甸栖息的高原鼢鼠为研究对象,探讨不同放牧强度下高原鼢鼠栖息地特征及选择因素,为合理控制草原鼠害及保护草地生物多样性提供科学依据。本研究选择4个不同放牧强度小区,调查每个小区的高原鼢鼠相对种群密度、土壤紧实度和容重、植物学组成、植物物种丰富度、地上生物量、可食牧草产量及各植物功能群产量,并分析高原鼢鼠相对种群密度与这些要素之间的关系。结果表明,放牧强度与高原鼢鼠相对种群密度有正相关关系但不显著(P>0.05),重度放牧区下高原鼢鼠相对种群密度最高,禁牧区高原鼢鼠相对种群密度最低;高原鼢鼠相对种群密度与土壤紧实度呈显著负相关(P<0.05),与土壤容重和地上生物量呈负相关但不显著(P>0.05);高原鼢鼠相对种群密度与杂类草产量有正相关关系但不显著(P>0.05),与可食牧草产量、禾草产量和物种丰富度有负相关关系但不显著(P>0.05);多元线性回归方程表明影响高原鼢鼠栖息地选择的主要因素是土壤紧实度(Y=205.36-0.145X1,X1为土壤紧实度)。
关键词:高原鼢鼠;放牧强度;栖息地特征;栖息地选择
青藏高原高寒草地是世界上海拔最高、面积最大的草地生态系统,在畜牧生产、涵养水源和维持生物多样性等方面发挥着重要作用[1]。近年来,由于全球气候变化及人为因素的影响,高寒草地退化严重,生产力下降以及生态环境恶化已经严重威胁着区域乃至全国社会和经济的可持续发展[2-3]。在影响草地生态系统的生物因子中,家畜的过度放牧以及啮齿动物种群数量的增加都具有重要的干扰作用[4]。放牧不仅作用于植物群落和土壤[5-7],而且也影响到草原啮齿动物栖息地选择[8]。啮齿动物栖息地适合度增加可以导致种群数量增加和栖息地面积扩大,加剧草地退化[9]。但是,放牧强度与地下啮齿动物栖息地适合度的关系,目前尚不清晰。因此,研究放牧干扰对草原啮齿动物栖息地选择影响,对明晰其在草地生态系统中的地位和作用,以及科学防治草原鼠害具有重要意义。
高原鼢鼠(Eospalaxbaileyi)是仓鼠科中4种营地下生活的啮齿动物之一,广泛分布于青藏高原,是青藏高原生物多样性的重要组成部分,在草地生态系统食物网及其相对应的能量流通和物质循环中有着独特的地位[10-12]。但高原鼢鼠采食牧草以及挖掘过程中向地表推出土丘,对草地生产和生态也会产生负面影响。同其他动物一样,高原鼢鼠栖息地的选择受诸多因素影响,以往的研究主要集中在对高原鼢鼠栖息地植被特征的分析,较少考虑到放牧强度的影响[13-14]。而本研究重点考虑不同放牧强度下,通过调查不同放牧强度区内高原鼢鼠相对种群密度以及土壤和植被因素,重点分析放牧干扰下高原鼢鼠栖息地的选择。
1材料与方法
1.1研究样地概况
样地设于甘肃省武威市天祝藏族自治县抓喜秀龙乡马营滩地区,该区位于东祁连山的天祝金强河河谷,地理坐标N 37°12′,E 102°46′,海拔2937 m,气候寒冷潮湿,太阳辐射强。年均温-0.1℃,1月平均温度-18.3℃,7月平均温度12.7℃,>0℃年积温1380℃;年降水量416 mm,多为地形雨,集中于7,8,9三个月。植被类型为高寒草甸,以莎草科、禾本科植物为主,混有其他杂类草,草群盖度达到90%左右,土壤为高寒草甸土,主要植物有珠芽蓼(Polygonumviviparum)、鹅绒委陵菜(Potentillaanserina)、早熟禾(Poaannua)、矮嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifoli)、垂穗披碱草(Elymusdahuricus)、球花蒿(Artemisiasmithii)、黄芪(Astragalusmembranaceus)等[15]。
1.2样地设置及实验时间
样地选取冬季牧场,依据不同放牧强度选择4块样地,每块样地面积3 hm2。放牧时间为当年11月至次年5月,放牧期均在3年以上,而禁牧区为已连续3年禁牧的草地。放牧强度以家畜头数、草地面积和利用时间而定,依照当地平均放牧强度,分别划分为重度放牧40羊单位/hm2(high grazing, HG)、中度放牧27羊单位/hm2(moderate grazing, MG)、轻度放牧10羊单位/hm2(light grazing, LG)、将禁牧区作为对照(control check, CK)。样地皆做相互独立的围栏保护,该围栏的设置是防止家畜在另一样地进行采食。所有植被和土壤调查时间均在2014年8月。
1.3高原鼢鼠相对种群密度调查
依据何俊龄等[16]单位面积新鼠丘数(当年8月以前所形成)表示各样地高原鼢鼠的相对种群密度。即相对种群密度=当年8月前新生鼠丘数量/样地面积。新土丘数量调查采用RTK(real-time kinematic)技术。RTK是目前使用测绘、建筑等方面的快速高精度定位技术[17]。首先设置RTK基准站,并将手持移动终端与基准站建立通讯联系。然后将手持移动终端放置在每个新土丘上,并记录其准确的经纬度位置,最后将手持移动终端记录的新鼠丘位置输入Arcgis软件,统计不同样地的新鼠丘数作为高原鼢鼠相对种群密度。根据高原鼢鼠相对种群密度,依次划分为高密度区(Ⅰ)、次高密度区(Ⅱ)、中密度区(Ⅲ)以及低密度区(Ⅳ),与之相对应的放牧强度区为HG、LG、MG和CK(图1)。
1.4植被性状调查
1.4.1地上生物量及植物学组成每个放牧区设置3个重复,每个重复面积为1 hm2。利用DWR(dry-weight rank)法测量地上产草量和植物学组成,统计植物学组成按照其可食牧草所占比例,计算出可食牧草的产量[18-27]。
1.4.2植物物种丰富度利用DWR草地快速监测法,记录每一样地出现的植物种数。
1.4.3植物功能群生物量按照DWR法所记录的植物学组成以及地上生物量,将其划分为禾本科、莎草科、杂类草3个经济功能群,按照每个功能群所占比例计算各功能群生物量。
1.5土壤性状调查
1.5.1土壤紧实度在每个样地均匀选择27个点,每个点用SC-900土壤紧实度仪每隔2.5 cm测量土壤紧实度,深度为0~20 cm,并在此点附近重复3次,然后求平均值。
1.5.2土壤容重每个样地内均匀选取27个点,每个点附近挖取1个剖面,深度20 cm,自表土向下每隔10 cm用100 cm3环刀取一个土壤样品并装入铝盒,采用铝盒烘干法测定其容重[28]。
1.6数据分析
用Excel 2007绘制图表;采用SPSS 17.0软件对不同样地间土壤、植被因子进行One-way ANOVA分析,分析样地间因子差异显著性;对高原鼢鼠相对种群密度和土壤、植被因子进行简单相关性分析及多元回归分析,制作相关系数矩阵及最优方程。
2结果与分析
2.1高原鼢鼠相对种群密度与放牧强度的关系

图1 高原鼢鼠相对种群密度与放牧强度的关系Fig.1 The relationship between grazing intensity andrelative population density of plateau zokor
从图1可以看出,放牧强度(羊单位/hm2)依次是HG(40)>MG(27)>LG(10)>CK(0),种群密度(土丘/hm2)的变化为Ⅰ区(55.67)>Ⅱ区(30.00)>Ⅲ区(16.67)>Ⅳ区(6.67)。HG区高原鼢鼠相对种群密度最高,LG区和MG区次之,CK区高原鼢鼠相对种群密度最低。高原鼢鼠种群相对密度与放牧强度有正相关关系但不显著(P>0.05,R=0.646)。
2.2高原鼢鼠相对种群密度与地上生物量和可食牧草产量的关系
由图2可以看出,高原鼢鼠低密度区(Ⅳ)地上生物量与可食牧草产量显著高于其他3个区(P<0.05)。高原鼢鼠高密度区(Ⅰ)、次高密度区(Ⅱ)、中密度区(Ⅲ)三者产草量无显著性差异(P>0.05)。
2.3高原鼢鼠相对种群密度与植物物种丰富度的关系
从图3可以看出,植物物种丰富度随着高原鼢鼠相对种群密度的升高,出现先升后降的现象,其峰值出现在高原鼢鼠次高密度区(Ⅱ),最低值出现在高原鼢鼠低密度区(Ⅳ)。
2.4高原鼢鼠相对种群密度与植物功能群产量的关系
由图4可以看出,禾草功能群和莎草功能群产量随着高原鼢鼠相对种群密度的升高表现出降低的趋势,低密度区(Ⅳ)的禾草和莎草产量显著高于其他3个区(P<0.05)。而对于杂类草功能群产量来说,随着种群密度的上升呈现增大趋势,最大值出现在高原鼢鼠高密度区(Ⅰ),最小值出现在高原鼢鼠低密度区(Ⅳ),低密度区的杂类草产量显著低于其他3个区(P<0.05)。
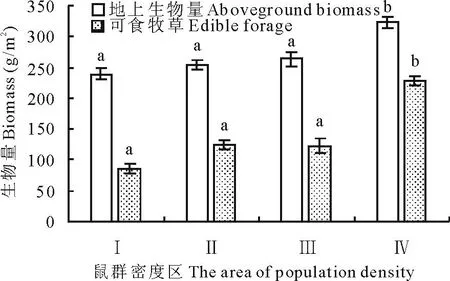
图2 高原鼢鼠相对种群密度与地上生物量和可食牧草产量的关系 Fig.2 The relationship between aboveground biomass,edible herbageyield and relative population density of plateau zokor
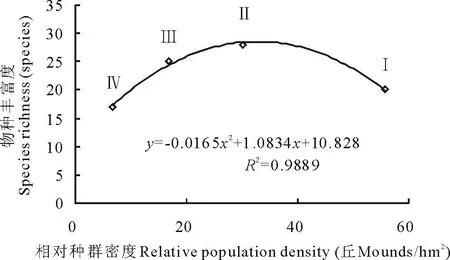
图3 高原鼢鼠相对种群密度与物种丰富度的关系Fig.3 The relationship between plants richnessand relative population density of plateau zokor
不同字母表示处理间差异显著(P<0.05),下同。Values with different letters indicate significant difference among treatments atP<0.05, the same below.
2.5高原鼢鼠相对种群密度与土壤紧实度的关系

图4 高原鼢鼠相对种群密度与植物功能群产量的关系 Fig.4 The relationship between the field of plant functionalgroups and relative population density of plateau zokor
从图5可以看出,土壤深度在0~20 cm,高原鼢鼠中密度区(Ⅲ)的土壤紧实度最大,高密度区(Ⅰ)最小,高原鼢鼠高密度区(Ⅰ)的土壤紧实度显著低于次高密度区、中密度区和低密度区(P<0.05),次高密度区(Ⅱ)、中密度区(Ⅲ)和低密度区(Ⅳ)三者无显著差异(P>0.05)。说明高原鼢鼠在紧实度较低的土壤分布相对较多。
2.6高原鼢鼠相对种群密度与土壤容重的关系
从图6可以看出,土壤深度在0~20 cm,高原鼢鼠低密度区(Ⅳ)的土壤容重最大,高密度区(Ⅰ)最小,且高原鼢鼠低密度区(Ⅳ)的土壤容重显著大于其他3个区(P<0.05),而高原鼢鼠高密度区(Ⅰ)、次高密度区(Ⅱ)、中密度区(Ⅲ)三者土壤容重无显著性差异(P>0.05)。
2.7土壤、植被性状与高原鼢鼠相对种群密度相关性及回归分析
土壤、植被性状与高原鼢鼠相对种群密度相关系数表明(表1),高原鼢鼠相对种群密度与土壤紧实度有显著负相关关系(P<0.05);杂类草产量与土壤容重有显著负相关关系(P<0.05),地上生物量与土壤容重有显著正相关关系(P<0.05),禾草产量与杂类草产量有显著负相关关系(P<0.05)。
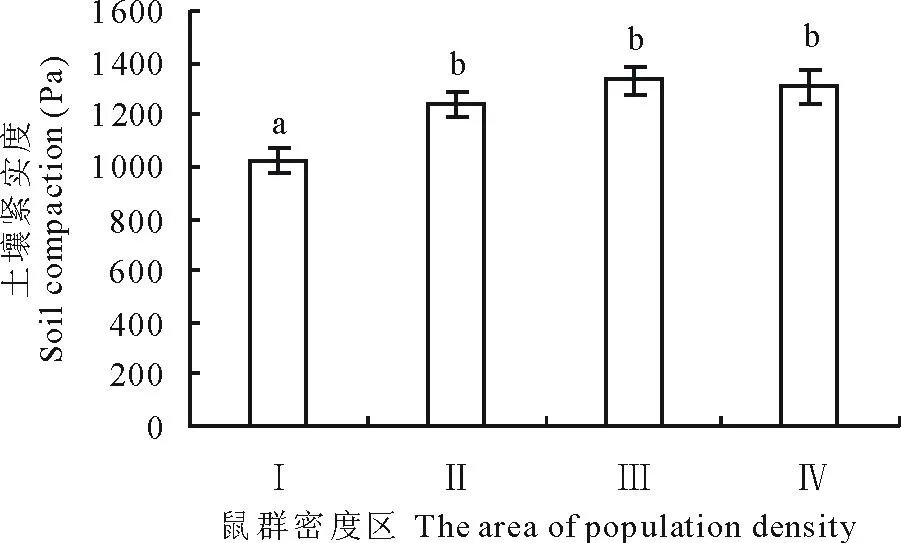
图5 高原鼢鼠相对种群密度与土壤紧实度的关系Fig.5 The relationship between soil compaction andrelative population density of plateau zokor
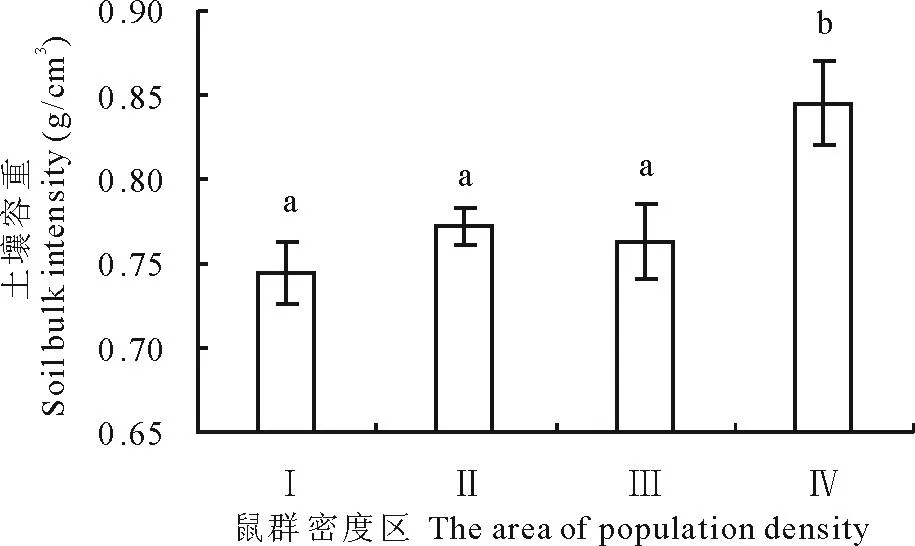
图6 高原鼢鼠相对种群密度与土壤容重的关系Fig.6 The relationship between soil bulk density andrelative population density of plateau zokor

项目ItemX0X1X2X3X4X5X6X10.577X2-0.986*-0.667X30.995**0.611-0.976*X40.983*0.631-0.959*0.996**X5-0.5380.1840.389-0.560-0.579X60.995**0.637-0.984*-0.999**0.994**-0.523X7-0.778-0.956*0.831-0.810-0.8290.098-0.827
X0:土壤容重;X1:土壤紧实度;X2:杂类草产量;X3:禾草产量;X4:地上生物量;X5:植物物种丰富度;X6:可食牧草产量;X7:高原鼢鼠相对种群密度。“*”和“**”表示两种因子的相关性达到显著水平(P<0.05)和极显著水平(P<0.01)。
X0:Soil bulk density;X1:Soil compaction;X2:Forb biomass;X3:Grass biomass;X4:Aboveground biomass;X5:Plants richness;X6:Edible forage biomass;X7:Population density of plateau zokor.“*” and “**”indicate significant correlation at the level of 0.05 and 0.01 respectively.
为了更好说明高原鼢鼠相对种群密度与植被、土壤因子的关系,采用逐步回归分析法得到最优回归方程Y=205.36-0.145X1,X1为土壤紧实度。该方程表明土壤紧实度对高原鼢鼠相对种群密度影响最大。
3讨论与结论
国内对高原鼢鼠栖息地研究开展较早且取得多项成果[29]。但是大部分研究没有考虑放牧干扰的影响,而是重点研究高原鼢鼠种群密度与土壤及植被特性的关系[14,30]。本试验通过划分4个不同放牧强度,研究不同放牧强度下高原鼢鼠相对种群密度与土壤物理性质、植被性状的关系,发现高原鼢鼠喜欢栖息地土壤疏松、杂类草产量高的草地。
栖息于不同放牧区的高原鼢鼠种群密度不尽相同。随着放牧强度的增加,优良牧草减少而杂类草增多,而杂类草又是高原鼢鼠喜爱的食物资源。本试验重牧区的杂类草产量最高(157.05 g/m2),高原鼢鼠相对种群密度也高于其他3个区(55.67丘/hm2)。而禁牧区的杂类草产量最低(95.34 g/m2),相应的鼠群密度也最低(6.67丘/hm2),因此高原鼢鼠偏好于杂类草产量较高的栖息地。此研究结果与刘伟等[30]研究不同放牧强度对植物及啮齿动物作用所得出的结果一致。在物种丰富度方面,张军等[14]在植被性状与高原鼢鼠栖息地适合度的关系研究中认为适宜的鼠密度有利于增加物种多样性。而本试验关于物种丰富度与种群密度的关系中可以看出其符合“中度干扰理论”[31],即一定的种群密度及放牧强度有利于增加物种的丰富度,高原鼢鼠种群密度较低的禁牧区物种丰富度反而最低,但是黄彬[32]认为植物的物种数随着鼢鼠种群密度的递增而出现下降趋势,二者呈现显著负相关关系,本试验的结果与此结果有所出入,有可能是因为本试验增加了放牧因素,较低的放牧强度可能有利于增加物种数。在地上生物量方面,重牧区的地上生物量显著低于禁牧区(P<0.05),高原鼢鼠相对种群密度与地上生物量有负相关关系,但不显著(R=-0.829,P>0.05),因为重牧区家畜的采食程度远远大于禁牧区,而且高原鼢鼠采食地下根系从而影响地上植物的生长,因此出现这种现象。
高原鼢鼠营地下生活,其独特的生活习性导致每年都将大量的土壤掘出地面形成土丘,土壤物理性质决定着高原鼢鼠的种群密度及分布。不同的放牧强度,家畜的践踏程度不同,导致土壤物理性质的改变。本研究结果表明,重牧区的土壤紧实度、容重显著低于其他3个区,高原鼢鼠相对种群密度与土壤紧实度呈现显著负相关关系(R=-0.956,P<0.05),与土壤容重有负相关关系但不显著(R=-0.778,P>0.05)。说明在重牧区虽然家畜的践踏程度要高于其他3个区,使表层土壤更加紧实,但是重牧区的高原鼢鼠相对种群密度也较大,因此地下洞道数量多,结构复杂,而所研究区域的高原鼢鼠平均洞道深度在0~20 cm内,所以会出现重牧区的土壤紧实度显著低于其他3个区(P<0.05)。同时,本试验通过多元线性回归分析表明,土壤紧实度是影响高原鼢鼠栖息地选择的主要因素。Wang等[33]在研究高原鼢鼠的挖掘行为与土壤硬度的关系中认为高原鼢鼠明显选择土壤硬度较小的地方栖息,黄彬[32]研究高原鼢鼠种群密度与土壤特性的关系中也表明随着鼠群密度的增加,土壤硬度、土壤容重下降,本试验的研究结果与此结果相似。土壤紧实度是一个反映土壤容重和水分以及质地的综合指标[34],本研究表明适宜高原鼢鼠栖息的生境选择主要倾向于土壤疏松的草地,其他因子如杂类草影响次之。
栖息地选择是动物生态学的一个传统研究领域,也是动物生态学众多分支的基础,同时栖息地选择研究可为动物保护措施的制定、利用生态手段防治草原鼠害提供科学依据[35]。但是目前关于栖息地选择理论的研究还相当匮乏,基础还很薄弱[36]。目前有关栖息地质量评价的指标存在很多争议,但国内大多数学者认为动物种群密度的高低依然是评价栖息地质量高低的指标[37]。因此,本研究依然将高原鼢鼠种群密度作为重要指标来确定其栖息地质量的好坏,从本研究的结果可以看出,在放牧干扰下,重度放牧区鼠群密度最高,栖息地特征表现为土壤紧实度、容重降低,杂类草产量较大,说明高原鼢鼠偏好此类型的栖息地。但是,由于自然条件下各种栖息地特征的非独立性,难以从中判断动物个体栖息地偏好实际所依据的信号,而且动物偏好某一类型的栖息地既可能由遗传决定,也可能受后期生长的影响,还可能受所处生活史阶段等生理状况的影响。因此,对于高原鼢鼠栖息地选择研究还需要大量的工作要做。同时,目前关于栖息地选择研究大多集中于对“栖息地特征”的研究[38],多数研究均建立在对自然条件下的调查数据的相关性分析,而相关性分析的缺陷是相关现象未必存在必然的因果关系,因此为了更准确分析动物个体的栖息地偏好,就需要可控试验的介入,通过对条件控制,严格设计栖息地选择试验,这也是在今后关于栖息地研究应该参考的方向。
References:
[1]Zhou X R, Guo Z G, Guo X H. The role of plateau pika and plateau zokor in alpine meadow. Pratacultural Science, 2010, 27(5): 38-44.
[2]Li W F, Tang Z M. Environment and safety of Qinghai-Tibet railway. Journal of Safety and Environment, 2003, 3(3): 65-67.
[3]Fan N C, Shi Y Z. A revision of the zokors of subgenusEospalax. Acta Theriologica Sinica, 1982, 2(1): 183-200.
[4]Melladoa M, Olverab A, Querob A. Dietary overlap between prairie dog (Cynomysmexicanus) and beef cattle in a desert rangeland of northern Mexico. Journal of Arid Environments, 2005, 62(3): 449-458.
[5]Rosiere R E. An evaluation of grazing intensity influence on california annual range. Range Manage, 1987, 40(2): 160-165.
[6]Zhang Y W, Han J G, Li Z Q. A study of the effects of different grazing intensities on soil physical properties. Acta Agrestia Sinica, 2002, 1(4): 74-78.
[7]Du Y G, Cao G M, Wang Q L,etal. Effect of grazing on surface character and soil physical property in alpine meadow. Journal of Moutain Science, 2007, 25(3): 338-343.
[8]Harald S, Atle M, Gunnar A. Sleep grazing and rodent populations: Evidence of negative interaction from a landscape scale experiment.Oecologa, 2005, 143(3): 357-364.
[9]Cui Q H, Jiang Z G, Lian X M,etal. Factors influencing habitat selection of root voles (Microtusoeconomus). Acta Theriologica Sinica, 2005, 25(1): 45-51.
[10]Fan N C, Wang Q Y, Zhou W Y,etal. The relationship between the plateau zokor population and vegetation damage. Proceedings of Alpine Meadow Ecosystem Congress[C]. Beijing: Science Press, 1988: 109-115.
[11]Zhang Y M, Liu J K. The biological characteristics of subterranean rodents and their roles in ecosystem. Acta Theriologica Sinica, 2002, 22(5): 144-153.
[12]Wu R N, Su B D, Bai M L. The influencing of subterranean rodents in agricultural ecosystem. Animal Husbandry and Feed Science, 2004, (6): 105-106.
[13]Han T H, Zhang W G, Ding L S. Features of vegetation inMyospalaxbaileyihabitat. Acta Prataculturae Sinica, 1999, 8(2): 43-49.
[14]Zhang J, Ge Q Z, Zhang W G,etal. Correlations between vegetation properties and fitness of plateau zokor habitat. Pratacultural Science, 2011, 28(5): 836-840.
[15]Li F X, Zhang D G. Indicators and recovery approaches of degenerated grassland in China. Grassland and Turf, 2005, (1): 24-28.
[16]He J L, Zhang J S, Yang Y B,etal. The study patterns and characteristics of plateau zokor mound. Acta Prataculturae Sinica, 2006, 15(1): 107-112.
[17]Wu J, Wang J C. Principle and advantages of RTK. China Investigation Design, 2009, (9): 52-53.
[18]Mannetje L T. The dry-weight-rank method for the botanical analysis of pasture. Grass and Forage Science, 1963, 18(4): 268-275.
[19]Jones R M, Hargreaves J N G. Improvements to the dry-weight-rank method for measuring botanical composition. Grass and Forage Science, 1979, 34(3): 181-189.
[20]Gillen R L, Smith E L. Evaluation of the dry-weight-rank method for determining species composition in tall grass prairie. Journal of Range Management, 1986, 5(7): 283-285.
[21]Barnes D L, Odendaal J J, Beukes B H. Use of the dry-weight-rank method of botanical analysis in the eastern Transvaal Highveld. Proceedings of the Annual Congresses of the Grassland Society of Southern Africa, 1982, 17(1): 79-83.
[22]Friedel M H, Chewings V H, Bastin G N. The use of comparative yield and dry-weight-rank techniques for monitoring arid rangeland. Journal of Range Management, 1988, 41(5): 430-435.
[23]Dowhower S L, Teague W R, Ansley R J,etal. Dry-weight-rank method assessment in heterogenous communities. Journal of Range Management, 2001, 16(6): 71-76.
[24]Dekker B, Kirkman K P, Du Plessis S I. Use of the dry-weight-rank method of botanical analysis in semi-arid savanna communities. African Journal of Range and Forage Science, 2001, 18(1): 63-66.
[25]Hughes L E. Grazing systems:more thoughts and observations. Rangelands, 2011, 33(1): 35-40.
[26]Rajput B S, Bhadauria U P S, Rajput R L. Study on specific leaf area and its relation with leaf area, leaf dry weight and seed yield in Mung bean. Journal of Life Sciences, 2014, 11(1): 247-250.
[27]Haydock K P, Shaw N H. The comparative yield method for estimating dry matter yield of pasture. Animal Production Science, 1975, 15(76): 663-670.
[28]Lv Y Z, Li B G. Edaphology[M]. Beijing: Chinese Agricultural Press, 2006.
[29]Zhou J W, Hua L M, Zuo S T,etal. Research progress in habitat selection of plateau zokor. Pratacul Tural Science, 2013, 30(4): 647-653.
[30]Liu W, Zhou L, Wang X. Responses of plant and rodents to different grazing intensity. Acta Ecologica Sinica, 1999, 19(3): 376-382.
[31]Wen L Y, Li Z F. The effects of disturbance on maintaining mechanism of species diversity. Journal of Northwest Normal University (Natural Science), 2006, 42(4): 87-91.
[32]Huang B. The Relationship between Plateau Zokor and Vegetation, Soil Factors[D]. Lanzhou: Lanzhou University, 2014.
[33]Wang Q Y, Zhou W Y, Wei W H,etal. The burrowing behaviour myospalax bailey and its relation to soil hardness. Acta Theriologica Sinica, 2000, 20(4): 277-283.
[34]Liu W G, Li L X, Xie H R,etal. Effect of soil bulk density on root morphology and biomass of vetiver grass seedlings. Acta Prataculturae Sinica, 2015, 24(4): 214-220.
[35]Dai Q, Gu H J, Wang Y Z. Theories and models for habitat selection. Zoological Research, 2007, 28(6): 681-688.
[36]Zhao Q S, Lou Y Q, Sun Y H. A review of statistical methods for evaluating animal habitat selection. Chinese Journal of Zoology, 2013, 48(5): 732-741.
[37]Jin L R, Sun K P, He H S,etal. Research advances in habitat suitability index model. Chinese Journal of Ecology, 2008, 27(5): 841-846.
[38]Bellis L M, Navarro J L, Vignolo P E,etal. Habitat preferences of lesser rheas in Argentine Patagonia. Biodiversity and Conservation, 2006, 15(9): 3065-3075.
参考文献:
[1]周雪荣, 郭正刚, 郭兴华. 高原鼠兔和高原鼢鼠在高寒草甸中的作用. 草业科学, 2010, 27(5): 38-44.
[2]李午峰, 唐祯敏. 青藏铁路的环境与安全. 安全与环境学报, 2003, 3(3): 65-67.
[3]樊乃昌, 施银柱. 中国鼢鼠(Eospalax)亚属分类研究. 兽类学报, 1982, 2(1): 183-200.
[6]张蕴薇, 韩建国, 李志强. 放牧强度对土壤物理性质的影响. 草地学报, 2002, 1(4): 74-78.
[7]杜岩功, 曹广民, 王启兰, 等. 放牧对高寒草甸地表特征和土壤物理性状的影响. 山地学报, 2007, 25(3): 338-343.
[9]崔庆虎, 蒋志刚, 连新明, 等. 田鼠栖息地选择的影响因素. 兽类学报, 2005, 25(1): 45-51.
[10]樊乃昌, 王权业, 周文扬, 等. 高原鼢鼠种群数量与植被破坏程度的关系-高寒草甸生态系统国际学术讨论会文集[C]. 北京: 科学出版社, 1988: 109-115.
[11]张堰铭, 刘季科. 地下鼠生物学特征及其在生态系统中的作用. 兽类学报, 2002, 22(5): 144-153.
[12]乌日娜, 苏布道, 白梦兰. 地下鼠对农业生态系统的影响. 畜牧与饲料科学, 2004, (6): 105-106.
[13]韩天虎, 张卫国, 丁连生. 高原鼢鼠栖息地植被特征. 草业学报, 1999, 8(2): 43-49.
[14]张军, 葛庆征, 张卫国, 等. 植被性状与高原鼢鼠栖息地适合度的关系. 草业科学, 2011, 28(5): 836-840.
[15]李凤霞, 张德罡. 草地退化指标及恢复措施. 草原与草坪, 2005, (1): 24-28.
[16]何俊龄, 张金沙, 杨莹博, 等. 高原鼢鼠土丘空间格局及主要特征研究. 草业学报, 2006, 15(1): 107-112.
[17]吴娇, 王军成. RTK工作原理及优缺点. 中国勘察设计, 2009, (9): 52-53.
[28]吕贻忠, 李保国. 土壤学[M]. 北京: 中国农业出版社出版, 2006.
[29]周建伟, 花立民, 左松涛, 等. 高原鼢鼠栖息地选择研究进展. 草业科学, 2013, 30(4): 647-653.
[30]刘伟, 周立, 王溪. 不同放牧强度对植物及啮齿动物作用的研究. 生态学报, 1999, 19(3): 376-382.
[31]文陇英, 李仲芳. 干扰对物种多样性维持机制的影响. 西北师范大学学报, 2006, 42(4): 87-91.
[32]黄彬. 高原鼢鼠种群密度与植被土壤因素的关系[D]. 兰州: 兰州大学, 2014.
[34]刘晚苟, 李良贤, 谢海蓉, 等. 土壤容重对野生香根草幼苗根系形态及其生物量的影响. 草业学报, 2015, 24(4): 214-220.
[35]戴强, 顾海军, 王跃招. 栖息地选择的理论与模型. 动物学研究, 2007, 28(6): 681-688.
[36]赵青山, 楼瑛强, 孙悦华. 动物栖息地选择评估的常用统计方法. 动物学杂志, 2013, 48(5): 732-741.
[37]金龙如, 孙克萍, 贺红士, 等. 生境适宜都指数模型研究进展. 生态学杂志, 2008, 27(5): 841-846.
*A study of habitat selection by plateau zokor in paddocks with different grazing intensities in the eastern Qilian Mountain region
CHU Bin, HUA Li-Min*, ZHOU Yan-Shan, LIU Li, LI Shuai
CollegeofRangelandScienceofGansuAgricultureUniversity,Lanzhou730070,China
Abstract:The objective of this study was to analyze the habitat selection and habitat characteristics of plateau zokor (Eospalaxbaileyi) in paddocks with different grazing intensities in the eastern Qilian Mountain region, in order to provide scientific information for controlling plateau zokor and conserving biodiversity in alpine meadows. Four paddocks with different grazing intensities were selected to investigate the zokor population density and habitat characteristics, including soil compaction and bulk density (g/cm3), vegetation botanical composition, plant species richness, aboveground plant biomass, and palatable forage biomass. A multiple regression analysis was used to determine the relationship between the plateau zokor population density and habitat characters. The zokor population density was highest in paddocks with the highest grazing intensity, and lowest in ungrazed paddocks (P>0.05). The zokor population density had a significant negative correlation with soil compaction (P<0.05) and a non-significant negative correlation with soil bulk density and aboveground plant biomass (P>0.05). The multiple regression for the relation between plateau zokor density and soil compaction, forb biomass, grass biomass, plants richness indicated that soil compaction was the dominant factor that influenced the plateau zokor habitat selection.
Key words:plateau zokor; grazing intensity; habitat factors; habitat selection
*通信作者Corresponding author. E-mail:hualm@gsau.edu.cn
作者简介:楚彬(1991-),男,甘肃兰州人,在读硕士。E-mail:ryan_713@163.com
基金项目:国家自然科学基金项目(No.31460635),农业部公益性行业科研项目(No.201203041)和甘肃省科技厅国际合作项目(No.1304WCGA174)资助。
*收稿日期:2015-05-12;改回日期:2015-07-17
DOI:10.11686/cyxb2015234
http://cyxb.lzu.edu.cn
楚彬,花立民,周延山,刘丽,李帅. 祁连山东段不同放牧强度下高原鼢鼠栖息地选择分析. 草业学报, 2016, 25(1): 179-186.
CHU Bin, HUA Li-Min, ZHOU Yan-Shan, LIU Li, LI Shuai. A study of habitat selection by plateau zokor in paddocks with different grazing intensities in the eastern Qilian Mountain region. Acta Prataculturae Sinica, 2016, 25(1): 179-186.
