人工模拟降雨格局变化对白刺幼苗生长的影响
2016-02-23张荣单立山李毅段桂芳段雅楠
张荣,单立山,2,李毅*,段桂芳,段雅楠,
张正中1,Жигунов Анатолий Васильевич3
(1.甘肃农业大学林学院,甘肃 兰州 730070;2.中国科学院寒区旱区环境与工程研究所,甘肃 兰州 730000;
3.圣彼得堡林业科学研究院,圣彼得堡 191028)
人工模拟降雨格局变化对白刺幼苗生长的影响
张荣1,单立山1,2,李毅1*,段桂芳1,段雅楠1,
张正中1,Жигунов Анатолий Васильевич3
(1.甘肃农业大学林学院,甘肃 兰州 730070;2.中国科学院寒区旱区环境与工程研究所,甘肃 兰州 730000;
3.圣彼得堡林业科学研究院,圣彼得堡 191028)
摘要:白刺是干旱荒漠区重要的建群植物种,研究其幼苗在不同降雨格局下的响应特征,可以为进一步研究降雨格局变化下白刺灌丛退化以及加快荒漠区植被恢复提供理论依据。本文通过人工控水试验,设置3个降雨量(增加30%、减少30%、不变)和2个降雨间隔时间(增加、不变)研究其对降雨格局变化的响应。结果表明,1)生长季白刺幼苗根长、叶生物量、地上生物量、总生物量和根冠比的变化主要由总降雨量引起;总降雨量对白刺生长的效应与降雨间隔时间紧密相关,但二者无显著交互作用。2)当降雨量一致时,增加降雨间隔时间有利于白刺幼苗冠幅、基径和各器官生物量的累积,尤其使叶生物量增加81%,从而使地上生物量增幅远大于地下生物量,导致根冠比降低。3)当降雨间隔时间一致时,降雨量减少30%对茎、地下生物量无显著影响,仅使根长显著增加86%,叶、地上生物量和总生物量分别显著减少67%,48%,27%,根冠比显著增加74%,高降雨量条件下的生物量均增加但无显著影响。因此,适当的增加降雨量和降雨间隔时间有利于白刺幼苗的生长及促进其植被恢复。
关键词:降雨格局;白刺;生物量;根冠比
近年来,全球气候剧烈变化直接导致陆地生态系统的结构和功能发生相应的改变,而降雨格局这一影响因素显得尤为突出[1-2]。据气象数据分析显示,亚洲中部干旱地区降水格局变化明显,主要表现为:降雨量增加、降雨时间分配改变、降雨强度增强[3]。同时,我国以荒漠生态系统为主的西北干旱地区在未来也将出现春、夏两季降雨明显增多的情况[4]。这势必影响土壤水分的时空动态变化[5],造成土壤含水量发生更大的变异性,进而导致植物的生理生态[6]及群落结构特征发生改变[7]。荒漠生态系统是典型的水分驱动型生态系统之一,与其他气候指标变化相比,荒漠生态系统对降雨格局的变化更为敏感[8],也更为剧烈。因此,通过控制降雨格局变化,研究其对荒漠生态系统的影响对于我们更加深入理解全球气候变化对植物种群动态过程具有重要的理论意义。
目前,虽然生态学家已经开展了大量关于降雨格局变化对荒漠生态系统结构与功能影响的研究,但研究结果与结论差异较大。Heisler-White等[9]研究表明,保持生长季总降雨量不变,延长降雨间隔时间且增加平均单次降雨量的降雨格局使地上净初级生产力(aboveground net primary production)显著增加。然而,Fay等[10]和Knapp等[11]在美国堪萨斯州Konza大草原(年平均降雨量835 mm)的研究中发现,总降雨量不变且增加降雨间隔时间会使地上净初级生产力减少。也有研究发现,当年降水量>200 mm时,有利于深根性的木本植物生长;<200 mm时,有利于浅根性的草本植物生长,且只有在生物学上有意义的有效降水才能促进植物生长和生殖[12]。可见,荒漠植物随降雨格局变化呈现不同的响应对策,但目前大部分关于降雨格局变化的研究都局限于植物的地上部分,对地下部分以及根冠比的研究则十分缺乏[13]。
白刺(Nitrariatangutorum)为蒺藜科白刺属强旱生灌木,是干旱荒漠区重要的建群植物种,也是对降雨最为敏感的荒漠植物之一[14]。目前对于荒漠植物幼苗生长随降雨量的变化已有大量报道,吴玉等[15]对干旱草本植物研究发现植株在低降雨量条件下总生物量和根生物量显著降低。李秋艳和赵文智[16]研究发现,不同降水梯度下泡泡刺(Nitrariasphaerocarpa)幼苗生物量及其分配随着降雨量的增加而呈抛物线变化趋势。可见,在不同的降雨条件下,荒漠植物幼苗生物量呈现出不同的分配策略。但大多数研究集中在单一降雨量对其生长的影响,并未涉及降雨量和降雨间隔期同时变化下的生长规律。因此本研究设置不同降雨量和降雨间隔期进行模拟试验,旨在探讨白刺在生长旺季形态特征及各器官生物量变化对模拟降水格局变化的响应和差异,进而为科学地预测白刺荒漠生态系统变化趋势和有效地防止白刺灌丛退化以及加快荒漠区植被恢复提供理论依据。
1材料与方法
1.1试验材料
供试苗木为白刺的2年生容器苗,供试土壤采用黄绵土,中等肥力,田间持水量为20.12%,土壤取回后过筛,去除杂质后备用。
1.2试验方法
1.2.1试验小区设计试验考虑降雨量、降雨间隔时间2个因素,分别设有3 个水平(降雨量增加、减少、不变)和2个水平(降雨间隔时间增加、不变),共设置 6 个降雨处理,每个处理重复3次,每个降雨处理占1个小区,共设计6个小区,每小区为4 m2(2 m×2 m)。相邻灌溉小区应用塑料做防渗处理,防渗膜隔离深度1 m。
1.2.2降雨格局变化设计根据全球变化模型预测结果,未来极端降水事件将增多,降雨间隔期也将延长。根据民勤荒漠区气象资料(1961-2008年)分析了研究区的降雨特征:1961-2008年的年平均降雨量为116 mm,降雨呈增加趋势,增加速率为4.462 mm/10 a;1985-2008年1~10 mm 的降雨强度占年降水总量的59.55%,10~20 mm 降雨强度占多年平均降水量的22.24%;根据24年当中10~20 mm 降水强度的增加速率最明显[3-4],且降水多集中于7-9月来设置降雨格局的变化,分别为降雨量减少30%(W-)、不变(W)、增加30%(W+);降雨间隔时间不变(T)和增加(T+),共设置 6 个降雨处理:1)自然降雨(以民勤地区近4年生长季月平均降雨量为对照),2)与自然降雨量一致,增加降雨间隔时间,3)增加降雨量,增加降雨间隔时间,4)减少降雨量,增加降雨间隔时间,5)增加降雨量,与自然降雨间隔时间一致,6)减少降雨量,与自然降雨间隔时间一致。降雨量不变是指与自然降雨量相同,降雨量增加或减少,指模拟雨量是在自然降雨量基础上增加或减少30%;降雨间隔时间不变是指人工模拟降雨的时间与自然降雨一致,降雨间隔时间增加,是指把试验期内每月第二次模拟降雨的时间定为两次自然降雨事件的间隔期延长50%的那一天,并将第二次自然降雨后与这一天之前发生的自然降雨一并添加,以达到延长降雨间隔期并增加大降雨事件次数的目的。
试验采用室外盆栽种植,花盆中装入供试土壤,花盆外径为38 cm,内径为36 cm,高度为26 cm。5月初每盆植入1株,定植后充分灌水以保证成活率,苗木成活后进行控水试验。7月初置于可移动的防雨棚内,通过遮雨棚避免当地降雨对幼苗的影响。试验于 2014年7 月4日至10月末进行,根据每月月平均降雨量的不同,以每个自然降雨事件的降雨量、降雨间隔时间为基准,通过人工添加水分,调节每月模拟降雨的水量和频次(表1),每隔30 d进行一次破坏性取样,即取样时间分别为2014年8月6日、9月6日、10月6日和11月6日。实验期间为防止土壤板结,需定期松土、除草和防止病虫害。
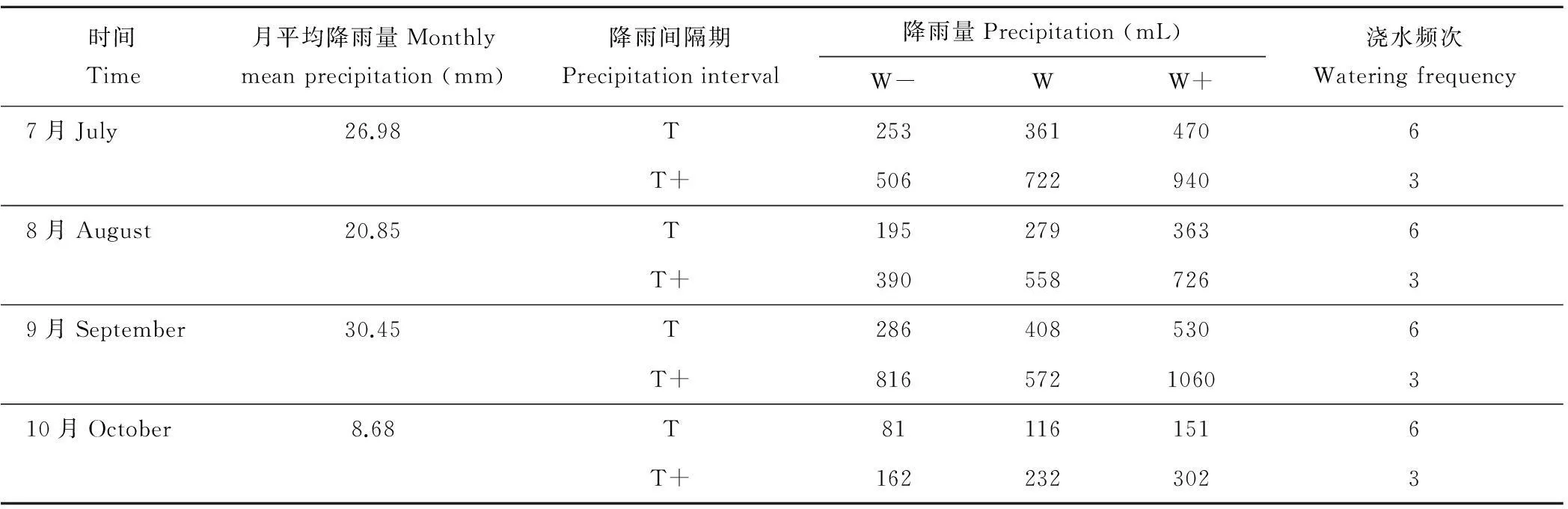
表1 实验中的降雨量和降雨间隔时间设置
注:T,降雨间隔时间为5 d;T+,降雨间隔时间为10 d;W,平均月降雨量;W-,减水30%;W+,加水30%。下同。
Note: T, mean precipitation interval of 5 days; T+, precipitation interval of 10 days; W, mean monthly precipitation; W-,water reduction by 30%;W+,water addition by 30%.The same below.
1.2.3取样与测定于2014年9月6日采集生长旺季白刺幼苗,取样时用手轻轻拍打花盆外侧使土体中的根系与土壤分离,用卷尺测量冠幅,主根长,游标卡尺测量基径后将完整的根系转入自封袋带回实验室进行室内处理,从基径处剪断幼苗,分装其地上部分(叶和枝)和地下部分,并在烘箱中以80℃恒温烘至恒重后称重, 得到白刺幼苗以下指标数据: 构件生物量、地上生物量、地下生物量、总生物量和根冠比。
1.3数据处理
采用Microsoft Excel进行数据分析和作图,采用SPSS 17.0软件进行one-way ANOVA法方差分析,显著性检验采用LSD法,用一般线性模型对总降雨量和降雨间隔时间的交互效应进行双因素方差分析。
2结果与分析
2.1降雨格局变化对白刺幼苗冠幅、基径和主根长的影响
由表2可以看出,总降雨量、降雨间隔时间及二者的交互作用对白刺幼苗冠幅和基径均无显著影响(P>0.05),而降雨量对主根长有显著影响(P<0.05),且总降雨量和降雨间隔时间交互作用对根长有极显著的影响(P<0.01)。由图1可知,在相同降雨量条件下,除主根长之外,延长降雨间隔时间白刺幼苗冠幅和基径均增加,其中冠幅平均增加39%,基径平均增加15%,但二者差异不显著(P>0.05)。在相同降雨间隔时间,降雨量的变化对白刺幼苗冠幅、基径和根长有不同程度的影响,其中降雨量增加30%各指标差异均不显著(P>0.05),而降雨量减少30%仅使主根长显著增加86%(P<0.05),且在W-T处理中达到最大,冠幅和基径则差异不显著(P>0.05)。
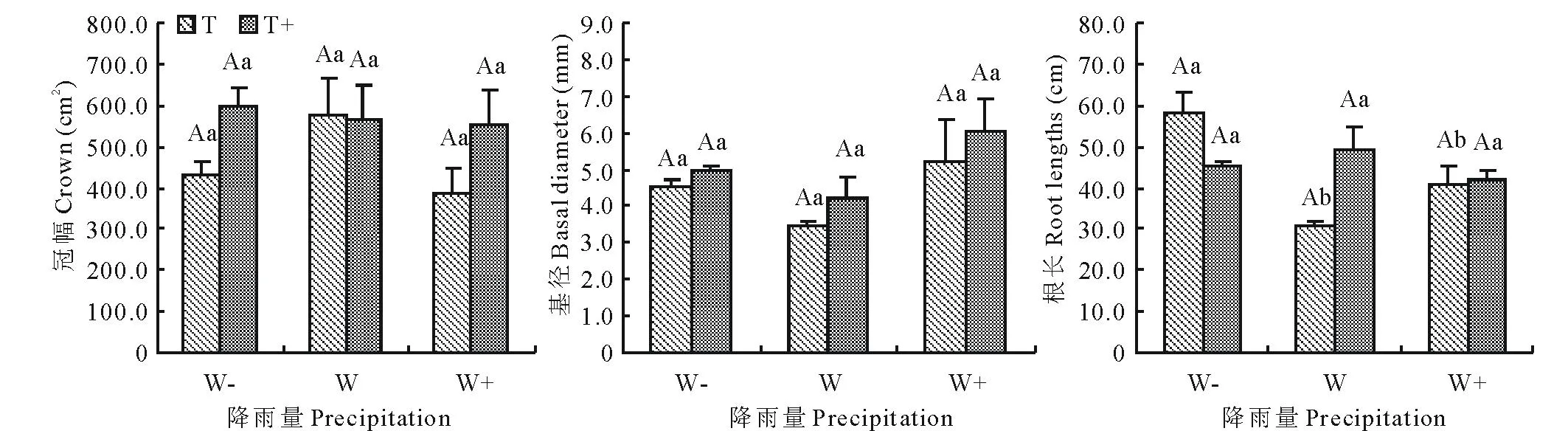
图1 不同降雨格局下白刺幼苗冠幅、基径和主根长的变化(平均值±标准误差, n=3)Fig.1 Dynamics of crown, basal diameter and main root length of N. tangutorum seedlingsin different precipitation patterns (mean±SE, n=3) 不同小写字母表示在相同的降雨间隔时间,降雨量变化与对照间差异显著(P<0.05);不同大写字母表示相同降雨量条件下,降雨间隔时间与对照间的差异显著(P<0.05)。下同。Different lowercase letters indicate significant difference between precipitation change and control within same precipitation interval (P<0.05); Different uppercase letters indicate significant difference between precipitation interval and control at same precipitation (P<0.05).The same below.
2.2降雨格局变化对白刺幼苗构件生物量的影响
双因素方差分析发现,总降雨量对叶生物量达到极显著水平(P<0.01),但降雨间隔时间、总降雨量和降雨间隔时间的交互作用对白刺幼苗构件(叶、茎)生物量均无显著影响(P>0.05)(表2)。由图2可知,在相同降雨条件下,降雨间隔时间延长使叶片生物量平均增加28%,茎生物量变化不规则,且二者差异不显著(P>0.05)。在相同降雨间隔时间,降雨量的变化对白刺幼苗茎、叶生物量有不同程度的影响,其中降雨量增加30%,叶生物量和茎生物量平均分别增加24%,15%,且二者差异不显著(P>0.05),但降雨量减少30%使茎、叶生物量均降低,特别是叶生物量平均显著地减少67%;可见,延长降雨间隔期增加大降雨事件有利于白刺幼苗茎、叶生物量的积累(图2)。
2.3降雨格局变化对白刺幼苗地上、地下和总生物量的影响
由表2可知,降雨量对白刺幼苗地上生物量和总生物量有显著影响(P<0.05),而对地下生物量影响不大(P>0.05)。降雨间隔时间、降雨量和降雨间隔时间的交互作用对地上、地下和总生物量无显著影响(P>0.05)。从图3可以看出,当降雨量一致时,地上、地下和总生物量的变化较不规则,但总的来讲降雨间隔时间增加各生物量均呈增加的变化趋势,且二者差异不显著(P>0.05)。当降雨间隔时间一致时,除地下生物量之外,随降雨量增加白刺幼苗地上生物量和总生物量均增加,其中降雨量增加30%使地上、地下和总生物量分别平均增加18%,48%和25%,且根生物量增加幅度远大于地上生物量,各处理间差异不显著(P>0.05);但降雨量减少30%仅使地上生物量平均显著减少48%,总生物量平均减少27%。地上生物量和总生物量的差异主要由降雨量引起,在降雨量较大的处理下(W+T和W+T+)各生物量更高。

图2 不同降雨格局下白刺幼苗叶片生物量和茎干生物量的变化(平均值±标准误差, n=3)Fig.2 Dynamics of leaf biomass, stem biomass of N. tangutorum seedlings in different precipitation patterns (mean±SE, n=3)

自变量Independentvariable变异来源(自由度)Variancesource(df)冠幅Crown基径Basaldiameter主根长Mainrootlength叶干重Leafbiomass枝干重Stembiomass地上生物量Abovebiomass地下生物量Belowbiomass总生物量Totalbiomass根冠比Root/shootT33.6720.1640.8221.8110.0070.1030.1060.4460.01W31.0511.4834.607*15.633**1.0893.875*1.4534.729*4.80*T×W61.0740.94711.054**0.0190.3510.2570.6150.0052.84
注:*表示显著水平(P<0.05); **表示极显著水平(P<0.01); T,降雨间隔时间效应;W,总降雨量效应;T×W,总降雨量与降雨间隔时间交互效应。
Note: “*” and “**” indicate significant correlation at the level of 0.05 and 0.01 respectively. T, precipitation interval effect; W, total precipitation quantity effect; T×W, precipitation×precipitation interval interaction effect.
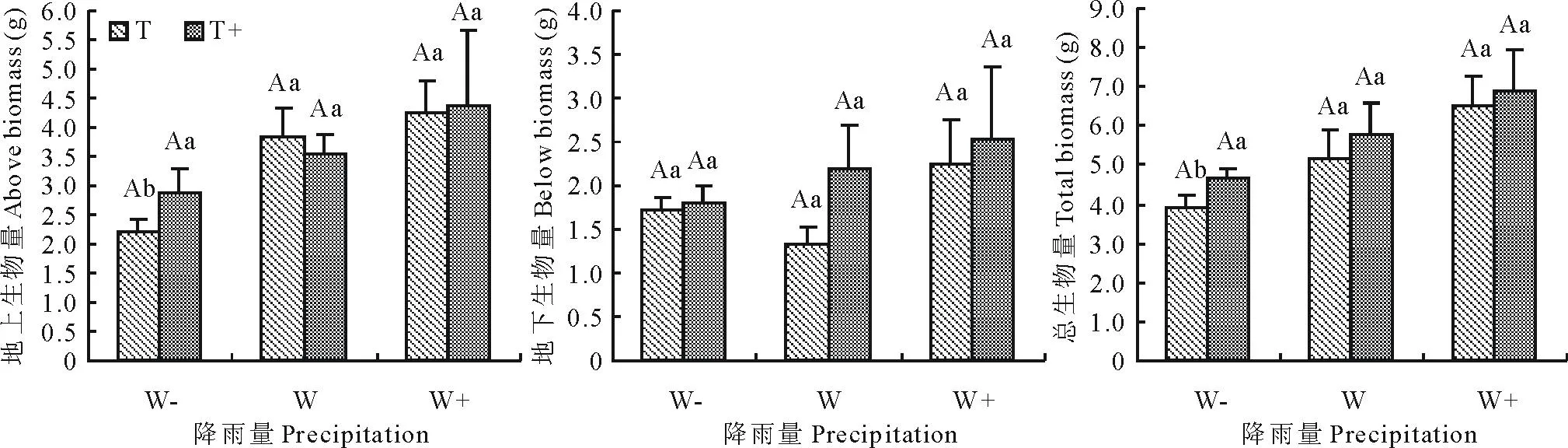
图3 不同降雨格局下白刺幼苗地上生物量、地下生物量和总生物量的变化(平均值±标准误差, n=3)Fig.3 Dynamics of above biomass, below biomass and total biomass of N. tangutorum seedlingsin different precipitation patterns (mean±SE, n=3)
2.4降雨格局变化对白刺幼苗生物量分配的影响

图4 不同降雨格局下白刺幼苗根冠比的变化(平均值±标准误差, n=3)Fig.4 Dynamics of root/shoot ratio of N. tangutorumseedlings in different precipitation patterns(mean±SE, n=3)
双因素方差分析结果表明,降雨间隔时间,降雨量和降雨间隔时间的交互作用对白刺幼苗根冠比无显著影响(P>0.05),而降雨量对其有显著影响(P<0.05)(表2)。单因素方差分析表明,当降雨量相同时,延长降雨间隔时间白刺幼苗地上生物量增幅远大于地下生物量(图3),因此,根冠比不同程度减小,但二者差异不显著(P>0.05)。当降雨间隔时间相同时,降水量减少或增加白刺幼苗根冠比均增加,但仅使低降雨量条件下的根冠比平均显著增加74%,高降雨量条件下差异不显著(P>0.05)。低降雨量条件下(W-T和W-T+)白刺幼苗根冠比更大。这说明低降雨量显著影响白刺幼苗地上部分的生长,而白刺叶片生物量的锐减对根冠比的增加也有较大的贡献,这也进一步说明白刺作为复苏植物,在水分减少的情况下可通过叶片脱落来增大根冠比(图4)。
3结论与讨论
3.1降雨格局变化对白刺幼苗冠幅、基径和主根长的影响
大量研究表明,增雨能促进荒漠灌木的生长,如新疆大叶苜蓿(Medicagosativacv. Xinjiang Daye)苗高、冠幅、基径和分枝数随土壤含水量的增加而增加[17],头状沙拐枣(Calligonumcaput-medusae)幼苗灌溉后的苗高、冠幅、基径均显著高于未经灌溉的植株[18],本试验中延长降雨间隔时间的大降水事件导致白刺幼苗冠幅、基径均增加的结果与此一致。根系是植物生长最重要的器官,植物对环境特别是对水分因子变化的响应主要是通过根系来实现的,在水分胁迫条件下可作为抗旱评价的指标[19]。本研究发现降雨量减少30%,白刺幼苗主根长显著增加86%,这与单立山等[20]研究红砂(Reaumuriasoongorica)在水分胁迫条件下缩小根直径,延长根系长度的研究结果一致,这可能在水分减少时植物体内存在某种信号转导和生理调节机制,通过增加垂直根生长来有效地利用深层土壤水,提高吸收能力,这样有利于苗木在干旱期利用地下水资源维持其成活和生长[21],表明白刺幼苗根系具有较强的抗旱协调能力,干旱胁迫条件下通过增加主根长等适应策略增强其抗旱能力。
3.2降雨格局变化对白刺幼苗生物量的影响
生物量作为植物群落最重要的数量特征之一,直接反映了生态系统生产者的物质生产量,是生态系统生产力的重要体现[22-23],而水分作为限制半干旱区沙地生态系统植被生产力和植被恢复的重要因子,它的增加能刺激植物生长,有效地提高植物生产力[24]。因此研究和分析降雨格局变化对荒漠植物幼苗生物量的影响显得十分必要。已有研究表明,降雨间隔期的变化直接影响内蒙古地区优势树种大针茅(Stipagrandis)幼苗的生长,等量的降水如果缩短降雨间隔期,土壤水分补充更好,能有效地促进大针茅各器官生物量的积累[25]。也有研究指出,综合降水大小和频率来看,等量的一次大降水事件会提高土壤的渗透率[26],进而使其产生更大的径流以及更少的蒸发损耗[27],将更有利于植物生物量的积累[28]。本试验中发现,降雨间隔时间由5 d增加到10 d白刺幼苗地上生物量显著增加,尤其是叶生物量显著增加,这与周双喜等[25]模拟降雨格局变化对内蒙古典型草原优势种大针茅幼苗影响的研究结果一致,与Fay等[10]在北美较湿润的草原地区进行的研究结果相悖,原因可能为干旱区的植物在单次大降雨事件中有相当一部分水分通过植物蒸腾或地表蒸发返回到了大气中[29-30],但由于大降雨事件中的水分向更深的土层中渗透, 通过蒸发而损失掉的水分比例相对较小,随降雨间隔时间增加深层土壤含水量增加,土壤中可供植物利用的水量更多, 而且存在时间更长[31],从而使水分可利用性提高。因此,增加降雨间隔期提高了土壤水分利用效率,从而促进白刺幼苗生长。
据已有研究表明,在水分供应充足时,植物幼苗通常会提高光合速率,利用其形态的可塑性来建立有利于截获光的足够叶面积指数(leaf area index)和充分利用光能的合理叶面积,保证植株在整体水平维持较高的碳获取能力,保持旺盛生长,从而提高各构件生物量的积累[32]。然而处于水分胁迫下的植物由于细胞增大与增殖最先受到抑制,光合面积减小,生长速率降低,植物各个构件生物量以及总生物量都会降低[33]。本试验中降雨量增加30%,白刺幼苗各器官生物量均呈增加趋势,这与大多数荒漠植物研究结果一致[34-35]。而降雨量减少30%白刺幼苗的茎、地下生物量差异不显著,仅使叶生物量极显著地减少,进而使地上生物量和总生物量均显著减少,这与肖春旺等[36]研究毛乌素沙地油蒿(Artemisiaordosica)随施水量变化的研究结果一致。这可能是生长季植株在水分减少的状态下,蒸腾量过大,植株萎蔫,气孔关闭影响CO2进入,影响体内有机物积累而使根系因得不到有机营养而停止发育和活力下降,致使其吸收力降低甚至停止,导致各项生理指标降低,植株停止生长,且白刺作为复苏植物通常以萎蔫老叶、发出新芽的方式来适应干旱,从而免受降水减少的影响。这表明白刺幼苗具有较强的抗旱性,水分减少时通过叶片脱落或休眠增加叶生物量分配模式,以降低植物蒸腾和光合作用来储存更多的水分和养分,忍耐水分亏缺。同时,这也证实了Enquist和Niklas[37]提出的植物在受到资源限制时,往往面临两难选择,即如何在叶(使碳的获取达到最大)和根(使地下部分资源的获取达到最大)之间分配生物量,从而使植物获得养分和水分最大限度的平衡的观点。
3.3降雨格局变化对白刺幼苗生物量分配的影响
大量研究表明,荒漠草原植物的根冠比较其他地方的大,这种规律证实了植物处于胁迫生境条件下能够分配更多的干物质到地下的假说[38]。另外,植物生物量产生和分配与外界环境密切相关,当植物根部受到土壤水分胁迫时,植物主动将光合有机物质较多的输送到根系,减缓地上部分生长,加速主根的生长,从而使植物根系向富水区延伸,扩大吸收面积,提高竞争力[39]。毛伟等[40]对沙质草地不同生活史植物生物量研究发现,一年生植物对环境扰动比较敏感, 如添加氮素和水分后一年生植物繁殖生物量比重和叶生物量比重发生明显改变。本实验过程中,低降雨量较短降雨间隔时间的条件下白刺叶片出现不同程度的凋落,叶干重相应极显著的减少,导致地上生物量大幅度减小,进而使根系生物量显著高于地上生物量,根冠比更大,这可能是由于水分减少增加了分配到根系的光合产物的比例,根冠比增大,这与李阳等[41]研究结果一致。表明白刺具有一定的自我调节保护能力,即在出现土壤干旱时,其光合产物倾向于向地下部分转移,增强自身竞争能力,最大可能的保持自身物质和水分平衡,白刺抗旱性的形成是其形态结构和生理生态功能对干旱环境长期适应的结果。而延长降雨间隔期增加降雨量,白刺幼苗根冠比小幅度减小,这可能是对白刺而言水分供应已经相对充足, 同时由于上文提到的降雨间隔时间适当增长引起的水分可利用性提高, 使由于地上生长增强而导致的对水分的较高需求可以得到满足, 因而没有形成更多的根, 根冠比降低。虽然降雨间隔期的作用很重要,但是降雨量的影响更大,总生物量,叶干重和根冠比的差异主要由降雨量引起,这可能与研究对象的生长时间较短有关。
综上所述,本实验得到以下结论:1)降雨量和降雨间隔时间对白刺幼苗的生长都有不同的影响,总降雨量对白刺生长的效应与降雨间隔时间紧密相关。2)相同降雨量,增加降雨间隔时间有利于白刺幼苗冠幅、基径和各器官生物量的积累;相同间隔时间,降雨量减少30%显著减少根长、叶干重和地上生物量,从而使根冠比远大于其他处理;降雨量增加30%根系生物量增加幅度大于地上生物量,根冠比降低。3)生长季白刺幼苗根长、叶干重、地上生物量、总生物量和根冠比的变化主要由总降雨量引起(P<0.05)。本研究仅考虑了水分变化对优势植物生长的影响,而事实上,植物生长对全球气候变化的响应还受到CO2浓度和气温变化以及CO2浓度、温度和水分之间交互作用的影响,对于这方面有待进一步研究。
References:
[1]Smith S D, Huxman T E, Zitzer S F,etal. Elevated CO2increases productivity and invasive species success in an arid ecosystem. Nature, 2000, 408: 79-82.
[2]Schwinning S, Sala O E, Loik M E. Thresholds, memory and seasonality: understanding pulse dynamics in arid/semiarid ecosystems. Oecologia, 2004, 141: 191-193.
[3]Gong D Y, Shi P J, Wang J A. Daily precipitation changes in the semi-arid region over northern China. Journal of Arid Environment, 2004, 59: 771-784.
[4]Xu L G, Zhou H F, Liang C,etal. Multi time scale variability of precipitation in the desert region of north China. Journal of Hydraulic Endineering, 2009, 40(8): 1002-1011.
[5]Pan Y X, Wang X P. Spatial variation of soil moisture in Revegetated desert area. Journal of Desert Research, 2007, 27(2): 250-256.
[6]Su Z Z, Lu Q, Wu B,etal. Potential impact of climatic change and human activities on desertification in China. Journal of Desert Research, 2006, 26(3): 329-335.
[7]Giorgi F, Mearns L O, Shields C,etal. Regional nested model simulations of present day and 2×CO2climate over the central plains of the U.S. .Climatic Change, 1998, 40: 457-493.
[8]Weltzin J F, Loik M E, Schwinning S,etal. Assessing the response of terrestrial ecosystems to potential changes in precipitation. Bioscience, 2003, 53: 941-952.
[9]Heisler-White J L, Knapp A K, Kelly E F. Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia, 2008, 158: 129-140.
[10]Fay P A, Carlisle J D, Knapp A K,etal. Altering rainfall timing and quantity in a mesic grassland ecosystem: design and performance of rainfall manipulation shelters. Ecosystems, 2000, 3: 308-319.
[11]Knapp A K, Fay P A, Blair J M,etal. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science, 2002, 298: 2202-2205.
[12]Walter H. Natural savannahs as a transition to the arid zone. Ecology of Tropical and Subtropical Vegetation[M]. Edinburgh: Oliver and Boyd, 1971: 238-265.
[13]Chou W W, Silver W L, Jackson R D,etal. The sensitivity of annual grassland carbon cycling to the quantity and timing of rainfall. Global Change Biology, 2008, 14: 1382-1394.
[14]He Y H, Tian Y L, Ye D M,etal. Model of aboveground biomass ofNitrariatangutorumand relationship between biomass and leaf area. Journal of Desert Research, 2005, 25(4): 541-546.
[15]Wu Y, Zheng X J, Li Y,etal. Photosynthetic responses and biomass allocation strategies of desert herbaceous plants under different precipitation patterns. Chinese Journal of Ecology, 2013, 32(10): 2583-2590.
[16]Li Q Y, Zhao W Z. Responses of seedings of five desert species to simulated precipitation change. Journal of Glacilogy and Geocryology, 2006, 28(3): 414-420.
[17]Luo Y Z, Li G. The effect of water stress on growth and biomass ofMedicagosativacv. Xinjiangdaye. Acta Prataculturae Sinica, 2014, 23(4): 213-219.
[18]Song C, Zeng F J, Liu B,etal. Influence of water condition on morphological characteristics and biomass ofCalligonumcaputmedusaeSchrenk seedings. Chinese Journal of Ecology, 2012, 31(9): 2225-2233.
[19]Ma T C, Yu R R, Chen R J,etal. Effect of drought stress simulated with PEG-6000 on root system in rice seeding. Chinese Journal of Eco-Agriculture, 2010, 18(6): 1206-1211.
[20]Shan L S, Li Y, Duan Y N,etal. Response of root morphology and water use efficiency ofReaumuriasoongoricato soil water change. Acta Botanica Boreali-Occidentalia Sinica, 2014, 34(6): 1198-1205.
[21]Canadell J, Zedler P H. Ecology and Biogeography of Mediterranean Ecosystems in Chile, Califonia and Australia[M]. New York: Springer Verlag, 1995: 177-210.
[22]Zhang F, Shangguan T L, Li S Z. Improvement on the modeling method of biomass brush. Chinese Journal of Ecology, 1993, 12(6): 67-69.
[23]Li J W. Forest Ecology[M]. Beijing: China Forestry Publishing House, 1994: 100-103.
[24]Neff J C, Townsend A R, Gleixner G,etal. Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature, 2002, 419: 915-917.
[25]Zhou S X, Wu D X, Zhang L,etal. Effects of changing precipitation patterns on seedlings ofStipagrandis, a dominant plant of typical grassland of Inner Mongolia, China. Chinese Journal of Plant Ecology, 2010, 34(10): 1155-1164.
[26]Schwinning S, Sala O E. Hierarchy of responses to resource pulses in arid and semiarid ecosystems. Oecologia, 2004, 141(2): 211-220.
[27]Wainwright J, Mulligan M, Thomes J. Plant sand water in dry lands. In: Baird A J, Wilby R L. Ecohydrology: Plants and Water in Terrestrial and Aquatic Environment[M]. London: Routledge, 1999: 78-126.
[28]Sala O E, Lauernroth W K. Small rainfall events: an ecologica role in semiarid regions. Oecologia, 1982, 53(3): 301-304.
[29]Noy-Meir I. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics, 1973, 4: 25-51.
[30]Sala O E, Lauenroth W K, Parton W J. Long-term soil-water dynamics in the shortgrass steppe. Ecology, 1992, 73: 1175-1181.
[31]Heisler-White J L, Blair J M, Kelly E F,etal. Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Global Change Biology, 2009, 15: 2894-2904.
[32]Ackerly D D, Bazzaz F A. Leaf dynamics, self shading and carbon gain in seedlings of a tropical pioneer tree. Oecologia, 1995, 101: 28-29.
[33]Fernández R J, Wang M B, Reynolds J F. Do morphological changes mediate plant responses to water stress? A steady-state experiment with two C4grasses. New Phytologist, 2002, 155: 79-83.
[34]Liu Z, Zeng F J, An G X,etal. Influence of irrigation amounts on seeding growth and biomass allocation ofCalligonumcaputmedusaeat the southern fringe of the Taklimakan desert. Journal of Desert Research, 2012, 32(6): 1655-1661.
[35]Zhong Z B, Zhou G Y, Yang L C,etal. The biomass allocation patterns of desert shrub vegetation in the Qaidam Basin, Qinghai, China. Journal of Desert Research, 2014, 34(4): 1042-1048.
[36]Xiao C W, Zhou G S, Zhao J Z. Effect of different water conditions on growth and morphology ofArtemisiaordosicaKrasch. seedlings in Maowusu sandland. Acta Ecologica Sinica, 2001, 21(12): 2136-2140.
[37]Enquist B J, Niklas K J. Invariant scaling relations across treedom inated communities. Nature, 2001, 410: 655-660.
[38]Wang W, Peng S S, Fang J Y. Biomass distribution of natural grasslands and it response to climate change in north China. Arid Zone Research, 2008, 25(1): 90-97.
[39]Wei Q S, Zhao M, Li C L,etal. Growth and biomass allocation ofChilopsislinearisunder different soil water stresses. Chinese Journal of Ecology, 2006, 25(1): 7-12.
[40]Mao W, Li Y L, Cui D,etal. Biomass allocation response of species with different life history strategies to nitrogen and water addition in sandy grassland in Inner Mongolia. Chinese Journal of Plant Ecology, 2014, 38(2): 125-133.
[41]Li Y, Qiman Y, Zhu Y. Effects of water stress on photosynthetic characteristics and biomass partition ofElaeagnusmoorcroftii. Acta Botanica Boreali-Occidentalia Sinica, 2006, 26(12): 2493-2499.
参考文献:
[4]徐利岗, 周宏飞, 梁川, 等. 中国北方荒漠区降水多时间尺度变异性研究. 水利学报, 2009, 40(8): 1002-1011.
[5]潘颜霞, 王新平. 荒漠人工植被区浅层土壤水分空间变化特征分析. 中国沙漠, 2007, 27(2): 250-256.
[6]苏志珠, 卢琦, 吴波, 等. 气候变化和人类活动对我国荒漠化的可能影响. 中国沙漠, 2006, 26(3): 329-335.
[14]何炎红, 田有亮, 叶冬梅, 等. 白刺地上生物量关系模型及其与叶面积关系的研究. 中国沙漠, 2005, 25(4): 541-546.
[15]吴玉, 郑新军, 李彦, 等. 荒漠草本植物在不同降雨模式下的光合响应和生物量分配策略. 生态学杂志, 2013, 32(10): 2583-2590.
[16]李秋艳,赵文智. 5种荒漠植物幼苗对模拟降水量变化的响应. 冰川冻土, 2006, 28(3): 414-420.
[17]罗永忠, 李广. 土壤水分胁迫对新疆大叶苜蓿的生长及生物量的影响. 草业学报, 2014, 23(4): 213-219.
[18]宋聪, 曾凡江, 刘波, 等. 不同水分条件对头状沙拐枣幼苗形态特征及生物量的影响. 生态学杂志, 2012, 31(9): 2225-2233.
[19]马廷臣, 余蓉蓉, 陈荣军, 等. PEG-6000模拟干旱对水稻幼苗期根系的影响. 中国生态农业学报, 2010, 18(6): 1206-1211.
[20]单立山, 李毅, 段雅楠, 等. 红砂幼苗根系形态特征和水分利用效率对土壤水分变化的响应. 西北植物学报, 2014, 34(6): 1198-1205.
[22]张峰, 上官铁梁, 李素珍. 关于灌木生物量建模方法的改进. 生态学杂志, 1993, 12(6): 67-69.
[23]李景文. 森林生态学[M]. 北京: 中国林业出版社, 1994: 100-103.
[25]周双喜, 吴冬秀, 张琳, 等. 降雨格局变化对内蒙古典型草原优势种大针茅幼苗的影响. 植物生态学报, 2010, 34(10): 1155-1164.
[34]刘镇, 曾凡江, 安桂香, 等. 塔克拉玛干沙漠南缘头状沙拐枣幼苗生长和生物量分配对不同灌溉量的响应. 中国沙漠, 2012, 32 (6): 1655-1661.
[35]钟泽兵, 周国英, 杨路存, 等. 柴达木盆地几种荒漠灌丛植被的生物量分配格局. 中国沙漠, 2014, 34(4): 1042-1048.
[36]肖春旺, 周广胜, 赵景柱. 不同水分条件对毛乌素沙地油蒿幼苗生长和形态的影响. 生态学报, 2001, 21(12): 2136-2140.
[38]王娓, 彭书时, 方精云. 中国北方天然草地的生物量分配及其对气候的响应. 干旱区研究, 2008, 25(1): 90-97.
[39]尉秋实, 赵明, 李昌龙, 等. 不同土壤水分胁迫下沙漠葳的生长及生物量的分配特征. 生态学杂志, 2006, 25(1): 7-12.
[40]毛伟, 李玉霖, 崔夺,等.沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应. 植物生态学报, 2014, 38(2): 125-133.
[41]李阳,齐曼·尤努斯, 祝燕. 水分胁迫对大果沙枣光合特性及生物量分配的影响. 西北植物学报, 2006, 26(12): 2493-2499.
*Effect of change to simulated precipitation patterns on seedling growth ofNitrariatangutorum
ZHANG Rong1, SHAN Li-Shan1,2, LI Yi1*, DUAN Gui-Fang1, DUAN Ya-Nan1, ZHANG Zheng-Zhong1, Жигунов Анатолий Васильевич3
1.CollegeofForestrySciences,GansuAgriculturalUniversity,Lanzhou730070,China; 2.ColdandAridRegionsEnvironmentalandEngineeringResearchInstitute,ChineseAcademyofSciences,Lanzhou730000,China; 3.SaintPetersburgAcademyofForestrySciences,SaintPetersburg191028,Russia
Abstract:Nitrariatangutorumis an important, dominant species in arid desert. Our objective was to explore the responses ofN.tangutorumseedlings to various precipitation patterns. This approach will provide basic theoretical data to predict seedling survival in degradedN.tangutorumscrub vegetation, and accelerate desert region recovery. In this article, we report an experiment using simulated rainfall to examine the effect of precipitation (increased by 30%, reduced by 30%, unchanged) and precipitation interval (increased, unchanged) onN.tangutorumseedlings. Differences in root length, leaf biomass, above ground biomass, total biomass and root-shoot ratio under the various simulated precipitation regimes are reported. The total precipitation and precipitation interval both strongly affectedN.tangutorumgrowth, but had no significant interaction. Plant crown, basal diameter and biomass were increased by extended precipitation interval, with the same total precipitation. As a result, the leaf biomass was increased by 81%, so that the above ground biomass accumulation was far greater than for below ground biomass, and the root-shoot ratio was decreased. With unchanged precipitation interval, reduced precipitation had no significant effect on stem and below ground biomass, but root length was increased by 86%; while leaf biomass, above ground biomass and total biomass were reduced by 67%, 48%, and 27%, respectively, and the root-shoot ratio was increased by 74%. The treatment in which precipitation was increased by 30% had no significant effect on biomass. Therefore, appropriate increase of precipitation and precipitation interval promote growth ofN.tangutorumseedlings, and vegetation restoration where this shrub is present.
Key words:precipitation patterns;Nitrariatangutorum; biomass; root-shoot ratio
*通信作者Corresponding author. E-mail:liyi@gsau.edu.cn
作者简介:张荣(1991-),女,甘肃天水人,在读硕士。E-mail: gsauzr@163.com
基金项目:中国博士后科学基金(2014M552514),国家自然科学基金项目(41361100,31360205),国家国际科技合作专项(2012DFR30830),甘肃省科技支撑计划项目(1204NKCA084)和科技部农业科技成果转化资金项目(2014GB2G100134)共同资助。
*收稿日期:2015-02-05;改回日期:2015-04-30
DOI:10.11686/cyxb2015076
http://cyxb.lzu.edu.cn
张荣,单立山,李毅,段桂芳,段雅楠,张正中,Жигунов Анатолий Васильевич. 人工模拟降雨格局变化对白刺幼苗生长的影响. 草业学报, 2016, 25(1): 117-125.
ZHANG Rong, SHAN Li-Shan, LI Yi, DUAN Gui-Fang, DUAN Ya-Nan, ZHANG Zheng-Zhong, Жигунов Анатолий Васильевич. Effect of change to simulated precipitation patterns on seedling growth ofNitrariatangutorum. Acta Prataculturae Sinica, 2016, 25(1): 117-125.
