退化红壤区针阔混交林3种优势树种树干液流特征比较
2016-02-09王永文
涂 洁,王永文
(南昌工程学院 生态与环境科学研究所,江西 南昌 330099)
es(T)=0.511×exp[17.502Ta/(Ta+240.97)]
VPD=es(T)-ea=es(T)(1-RH)
退化红壤区针阔混交林3种优势树种树干液流特征比较
涂 洁,王永文
(南昌工程学院 生态与环境科学研究所,江西 南昌 330099)
运用Granier热扩散法对退化红壤区针阔混交林3种优势树种杉木(Cunninghamialanceolata)、马尾松(Pinusmassoniana)和木荷(Schimasuperba)树干液流速率进行测定,结合自动气象站同步监测太阳净辐射、空气温度、空气相对湿度等气象因子,为退化地区植被恢复林分结构配置以及林地水分管理提供理论依据。结果表明:3树种晴天树干液流呈“昼高夜低”的单宽峰型曲线,但液流启动、到达峰值的时间以及数值大小存在差异。木荷比杉木、马尾松提前0.5 h启动、到达峰值。木荷液流速率平均值分别为杉木、马尾松的4倍和1.5倍。3树种雨天树干液流呈多峰型曲线,液流速率值明显低于同时期晴天水平。3树种树干液流与太阳净辐射、水汽压亏缺昼夜变化间存在时滞时间,表现为树干液流滞后太阳净辐射,而提前于水汽压亏缺。偏相关分析结果表明:树干液流速率与空气温度、太阳净辐射、水汽压亏缺呈极显著正相关,与空气相对湿度呈极显著负相关。在考虑液流速率与气象因子间时滞现象的情况下,采用逐步剔除法建立各生长时期液流速率与气象因子的多元线性回归方程。所有回归方程的拟合效果良好,复相关系数基本在0.8以上,经F检验,均达到极显著水平。在整个生长季,太阳净辐射、空气温度和空气相对湿度共同解释杉木液流速率的81.3%~92.2%,太阳净辐射和水汽压亏缺共同解释马尾松液流速率的91.9%~93.6%。对于木荷而言,在4、5、6、7月,太阳净辐射和水汽压亏缺共同解释液流速率的72.1%~92.5%,而在8、9、10月,太阳净辐射和空气相对湿度共同解释液流速率的80.7%~92.7%。因此,在整个生长季,太阳净辐射为影响3树种液流速率的主导因子。
树干液流; 退化红壤区; 杉木; 马尾松; 木荷; 气象因子
红壤是我国南方地区重要的土壤资源,总面积占全国土地面积的22.7%。过去由于自然与人为活动的干扰,该区曾一度成为我国水土流失范围最广、严重程度仅次于黄土高原的区域。近年来,基于人工植被重建的生态恢复措施在退化红壤治理中得到了广泛应用,多数研究围绕植被生物量与生产力[1]、植物群落多样性[2]、土壤碳循环[3]、生态环境效应[4]等方面展开,而有关植被恢复树种水分利用特征及其对环境因子响应的研究仅见少量报道[5]。树干液流是一个重要的水分生理生态参数,反映了树木的水分利用特征及其对环境的响应[6]。采用热扩散法可以在自然状态下连续测定树干液流,测得的数据能够准确地反映水分在树木体内的传输速率,由此估算整树或林分蒸腾速率[7]。如果与空气和土壤因子传感器相结合,可以掌握土壤-植物-大气连续体(SPAC)水分传输的动态变化规律[8]。热扩散法因其具有精度高、操作简便、对树木损害小等优点,已在树木生理学、森林生态学和森林水文学相关研究中得到广泛应用。目前,国内对树干液流特征及其与环境因子关系的研究十分活跃,其中对南方地区树种树干液流的报道多数集中于单一树种[9-12],关于多个树种液流特征的研究不多[13-14],尤其是对退化红壤区针阔混交林优势树种树干液流的对比研究更是少见报道。
采用热扩散法对马尾松、杉木、木荷树干液流速率进行连续测定,探讨同一立地条件下针阔混交林3种优势树种液流日变化规律以及树种间差异,不仅可以科学评价不同植被恢复树种对环境水分的利用和调节能力,还可为退化地区植被恢复林分结构配置以及林地水分管理提供理论依据。
1 材料和方法
1.1 试验样地的选择与概况
研究区位于江西省泰和县中国科学院千烟洲试验站(26°44′48″N,115°04′1″E),海拔多在100 m左右,相对高度20~50 m,地形起伏,丘陵众多,坡度多在10°~30°。林地土壤为红壤,母质由白垩系红色和紫红色砾岩、砂岩和泥岩组成,呈典型的亚热带红壤丘陵景观。区内年均气温17.9 ℃,≥0 ℃活动积温6 523 ℃,太阳总辐射4 223 MJ/m2,年日照时数1 306 h,无霜期323 d,年降水量1 542 mm,年均相对湿度84%,具有典型的亚热带季风气候特征。样地设在试验站核心区1985年前后营造的针阔混交林内,面积149.19 m2,造林密度2 334株/hm2,郁闭度0.9以上。乔木层为以杉木(Cunninghamialanceolata)、马尾松(Pinusmassoniana)、木荷(Schimasuperba)等为主的针阔混交林,林下分布少量山矾(Symplocoscaudate)、黄檀(Dalbergiahupeana)、鳞毛蕨(Dryopterisfilix-mas)、淡竹叶(Lophatherumgracile)等。
1.2 研究方法
1.2.1 树干液流与环境因子的测定 根据探针长度和被测木具有代表性的原则,选取生长良好、树干通直、无被挤压的3种优势树种各3株(表1)。为避免阳光直射引起的测量误差,将探针(TDP-30,Dynamax公司)安装在树干北面距地面1.3 m处。刮去外层死树皮,把两孔距为40 mm的钻孔模块放在待测位置上,用直径为1.32 mm钻头钻取2个30 mm长的孔,两孔尽量平行并与树干垂直。将30 mm长、直径1.3 mm的加热针和参考针分别插入上部孔和下部孔。探针另一端与数采器(DT-50,Data Taker 公司)连接。环境因子利用Dynamax公司提供的空气温度、空气相对湿度、太阳辐射传感器和数据采集器组成的全自动微型气象站测定。所有数据的采集时间间隔均设为30 min。

表1 样木基本特征
采用水汽压亏缺(VPD)综合表达空气温度与空气相对湿度的协同效应,其计算公式是:
es(T)=0.511×exp[17.502Ta/(Ta+240.97)]
(1)
VPD=es(T)-ea=es(T)(1-RH)
(2)
式中,Ta为空气温度,RH为空气相对湿度,es(T)为Ta空气温度下的饱和水汽压(kPa),ea为周围气体水汽压(kPa)。
树干液流速率v[cm3/(cm2·s)]由Granier(1987)经验公式计算得到。
(3)
式中,ΔT为两探针间的温差,ΔTmax为连续7~10 d所测ΔT数据中的最大值[15]。
1.2.2 数据处理 利用Dynamax公司提供的软件进行数据下载和保存,采用Kaleida Graph 3.6软件绘图,并运用 SPSS软件进行种间差异显著性t检验、树干液流速率与环境因子间相关性以及多元线性回归分析。为了分析树干液流速率与环境因子间的时滞时间,采用错位对比法,按照观测时间顺序,建立液流速率与对应太阳净辐射(NR)、水汽压亏缺的数据系列,然后将其逐次按30 min进行错位移动,分析错位移动后数据的相关关系。当相关系数达到最大值时,所对应的错位时间即为液流速率与对应太阳净辐射、水汽压亏缺的时滞时间[16]。
2 结果与分析
2.1 3树种树干液流速率日变化
选取3树种生长季典型晴天和雨天树干液流数据作为研究对象,分析不同天气条件下杉木、马尾松和木荷的树干液流速率日变化规律及种间差异。
晴天(8月28日),3树种树干液流日变化呈“昼高夜低“的单宽峰曲线(图1c),其表观变化趋势与太阳净辐射、水汽压亏缺曲线基本吻合(图1a)。液流于6:30—7:00开始启动,其中杉木、马尾松的启动时间约比木荷晚0.5 h。随着太阳净辐射的增强,气温逐渐升高,空气相对湿度降低,叶片内外水汽压亏缺增大,液流速率不断增大。各树种液流于10:30—11:00达到峰值,其中杉木、马尾松到达峰值的时间比木荷推迟约0.5 h。之后于峰值附近持续6 h,形成由多个小峰组成的“高峰平台”。16:30起液流速率急剧下降,20:00后下降速度减慢并缓慢到达谷底值。不同树种树干液流速率间存在极显著差异(P<0.01),其中木荷液流速率日均值最高[(0.004 7±0.000 9)cm3/(cm2·s)],马尾松[(0.003 1±0.000 5)cm3/(cm2·s)]次之,杉木[(0.001 2±0.000 3)cm3/(cm2·s)]最低。木荷液流速率日均值分别为杉木、马尾松的4倍和1.5倍。
雨天(9月6日),根据当地自动气象站监测数据,降雨时段为3:00—14:00,加上此时太阳净辐射弱,空气温度低,空气相对湿度高(图1b),极大地降低了叶片气孔内外的蒸汽压梯度,降雨期间3树种液流均没有明显的启动,一直维持在较低的水平。14:30以后,随着降雨的停止,太阳净辐射和水汽压亏缺逐渐增大,3树种液流速率才开始显著升高并持续了3 h的小幅波动,16:30起液流速率急剧下降,20:00以后下降速度减慢并缓慢降低到较低水平(图1d)。3树种雨天日均液流速率均显著(P<0.05)低于同时期晴天液流水平,木荷[(0.001 1±0.000 4)cm3/(cm2·s)]和马尾松[(0.001 0±0.000 2) cm3/(cm2·s)]日均液流速率接近,杉木[(0.000 2±0.000 0)cm3/(cm2·s)]最小,几乎为木荷和马尾松的1/5。相比晴天,3树种雨天液流启动推迟,结束提前,液流曲线峰值低,其中木荷、马尾松、木荷液流速率分别是同生长时期晴天液流水平的1/4、1/3、1/6。因此,雨天对3树种液流流速均有不同程度的减弱作用,减弱程度与植物本身的生态学特性有关。
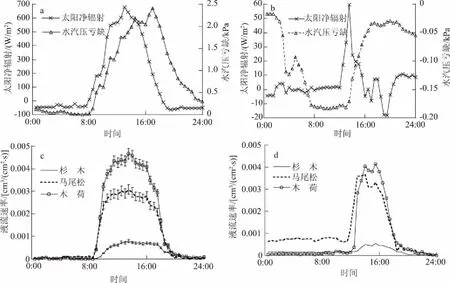
a、b分别为晴天、雨天太阳净辐射和水汽压亏缺日变化; c、d分别为晴天、雨天3树种液流速率日变化
2.2 3树种树干液流速率与气象因子的相关性
通过对3树种典型日液流与气象因子的连续观测发现,杉木、马尾松、木荷树干液流均滞后太阳净辐射,而提前于水汽压亏缺。其中木荷树干液流与太阳净辐射间时滞时间为30 min,与水汽压亏缺间时滞时间为60 min。杉木、马尾松树干液流与太阳净辐射间时滞时间均为60 min,与水汽压亏缺间时滞时间均为90 min。为了分析生长季各月气象因子对3树种树干液流的影响,分别对生长季各月液流速率与气象因子进行偏相关分析(表2),即通过控制相关变量,分析主变量与液流速率的相关性,排除进入方程的其他相关变量的影响,从而更好地了解主变量对液流的影响。
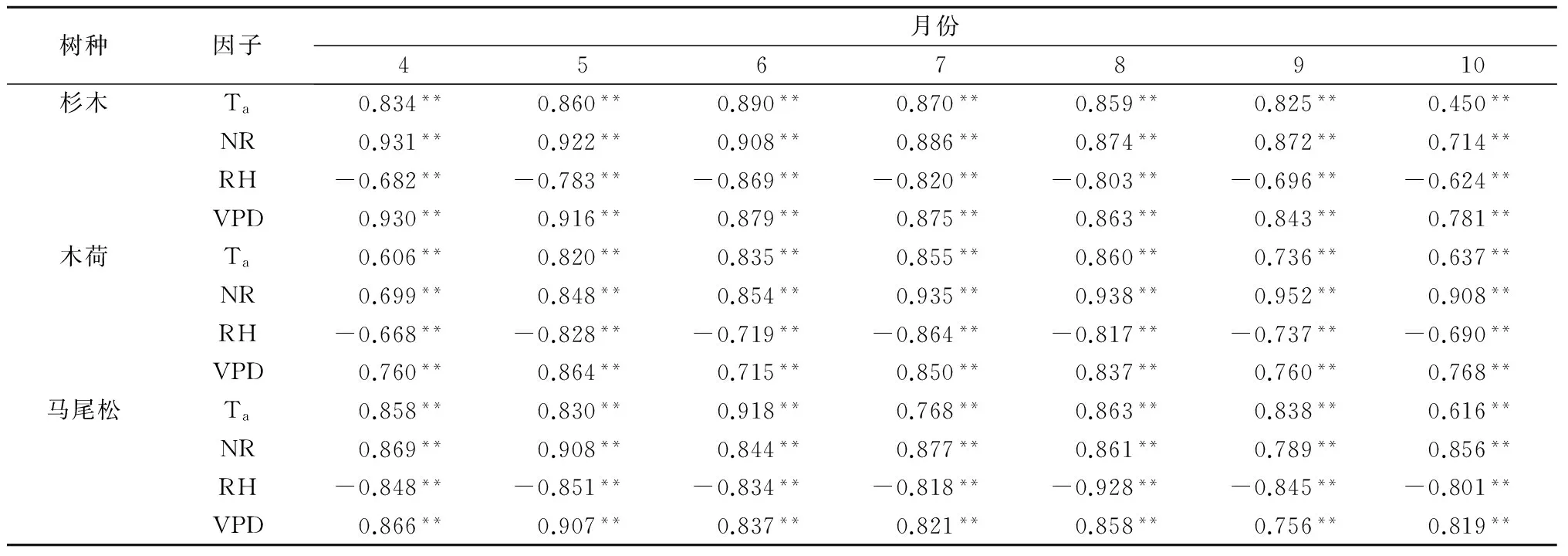
表2 3树种树干液流与气象因子偏相关分析
注:** 表示在0.01水平极显著相关(双侧)。
偏相关分析结果显示,3树种液流速率均与空气温度、太阳净辐射、水汽压亏缺呈极显著正相关,与空气相对湿度呈极显著负相关,说明这些气象因子对液流的影响趋势是一致的。除10月外,杉木液流速率与太阳净辐射相关性最强(偏相关系数r2=0.872~0.931),其次为水汽压亏缺(r2=0.843~0.930)或空气温度(r2=0.825~0.890)。而在10月,杉木液流速率与水汽压亏缺相关性最强(r2=0.781),其次为太阳净辐射(r2=0.714)。除4、5月外,木荷液流速率均与太阳净辐射相关性最强(r2=0.854~0.952),其次为空气温度(r2=0.736~0.860)或水汽压亏缺(r2=0.715~0.768)。而在4、5月,木荷液流速率主要由水汽压亏缺决定(r2=0.760~0.864),其次为太阳净辐射(r2=0.699~0.848)。除6、8、9月外,马尾松液流速率主要由太阳净辐射决定(r2=0.856~0.908),其次为水汽压亏缺(r2=0.819~0.907)。而在6月,马尾松液流速率与空气温度相关性最强(r2=0.918),在8、9月,马尾松液流速率与空气相对湿度相关性最强(r2=0.845~0.928)。
为了分析生长季各月气象因子对3树种液流速率的综合影响,在考虑液流速率与气象因子间时滞效应的情况下,采用逐步剔除法进行多元线性回归,以1%的可靠性作为因变量入选和剔除临界值,建立3树种液流速率与气象因子的多元线性回归方程(表3)。

表3 3树种液流速率与环境因子的多元线性回归方程
结果表明,所有回归方程的拟合效果良好,复相关系数R2基本在0.8以上,F检验以及常数、相关系数的t检验均通过0.01水平上的显著性检验,说明这些回归方程均能较好地揭示3树种液流速率与气象因子的变化规律。然而,树干液流速率对气象因子的响应程度随树种和生长时期不同而存在较大差异。从回归方程的复相关系数看出,空气温度、太阳净辐射和空气相对湿度3个因子可以共同解释杉木液流速率的81.3%~92.2%。在4、5、6、7月,太阳净辐射和水汽压亏缺对木荷液流速率起主导作用,两者可以共同解释木荷液流速率的72.1%~92.5%。而在8、9、10月,木荷液流速率主要由太阳净辐射和空气相对湿度共同决定,两者可以共同解释木荷液流速率的80.7%~92.7%。在整个生长季,太阳净辐射和水汽压亏缺可以解释马尾松液流速率的91.9%~93.6%。总的来说,太阳净辐射与杉木、马尾松、木荷液流速率都具有极显著的相关性,说明太阳净辐射是影响树干液流的最直接因子。
3 结论与讨论
不同天气条件下,3树种液流日变化规律存在显著性差异。晴天,杉木、马尾松、木荷液流呈基本一致的“单宽峰”曲线,与前人得出的杉木[17]、马尾松[18]、木荷[13,19]日变化规律相同,而与加勒比松(Pinuscaribaea)[20]、胡杨(Populuseuphratica)[21]等液流日变化呈“多峰型”曲线的结论不同。3树种液流启动、到达峰值的时间以及数值大小不同,主要表现在杉木、马尾松启动和到达峰值的时间比木荷晚0.5 h,说明木荷树干液流随环境因子的变化更加迅速。木荷液流速率日均值分别为杉木、马尾松的4倍和1.5倍。对于处于生长季的针阔混交林,阔叶树种木荷的蒸腾能力明显比针叶树种马尾松、杉木强,与黄德卫等[13]对鼎湖山针阔混交林的研究相同。这主要与针叶树和阔叶树间的生物学差异有关,针叶树马尾松水分在木质部中是通过管胞传输,而阔叶树木荷水分在木质部中通过导管传输,管胞的长度及直径都小于导管,使得马尾松木质部导水性比阔叶树差,加之厚的针叶结构,限制了马尾松叶片水平的蒸腾速率[22]。雨天,3树种树干液流速率均低于晴天,液流速率相对稳定,与天目柳杉(Cryptomeriafortunei)[10]雨天液流的日变化规律相同。主要原因可能是雨天太阳辐射低,空气相对湿度大,叶片内外的蒸汽压梯度显著降低,而且降雨还促使叶片气孔关闭,因而对液流上升有较大的制约作用,导致液流速率很低。
杉木、马尾松、木荷树干液流均滞后于太阳净辐射,而提前于水汽压亏缺,这是由于根据SPAC理论,土壤-植物-大气间存在一定大小的水势梯度,当大气温度升高时,大气水势与叶水势间产生水势差,树木通过蒸腾拉力从根系吸水,再通过叶片蒸腾到空气中去,而叶片、树枝和树干存在水容调节,水分从树干传输到叶片需要一段时间,从而表现为液流滞后于气象因子[23]。Granier等[24]把针叶树产生时滞的真正原因归结为树木组织内在的吸收和释放。本研究发现,木荷树干液流滞后于净辐射30 min,提前于水汽压亏缺60 min。杉木、马尾松树干液流滞后于净辐射60 min,提前于水汽压亏缺90 min。郑怀舟等[18]得出长汀水土流失区马尾松树干液流与太阳辐射间存在3 h左右的时滞,时滞时间显著大于本研究结果,其原因值得进一步的探究。刘鑫等[25]认为杉木秋、冬两季液流启动时间滞后于太阳辐射约2.5 h,而春、夏两季则无明显的时滞效应,各季节液流达到峰值的时间均滞后太阳辐射约2.5 h。黄德卫等[13]发现干湿季马尾松、木荷液流速率与光合有效辐射、水汽压亏缺间的时滞性存在明显的季节差异,与王慧梅等[26]对落叶松(Larixgmelinii)、孙振伟等[14]对大叶相思(Acaciaauriculaeformis)的研究结论一致。因此,综合衡量树木的时滞时,需要具体考虑某个月份的时滞效应。Oguntunde等[27]发现木薯(Manihotesculenta)液流与总辐射之间没有时滞,而液流提前于饱和水汽压差110 min。Ford等[28]得出32年生火炬松(Pinustaeda)树干液流滞后水汽压亏缺的时间为0~15 min,滞后太阳辐射的时间则为15~30 min。不同树种的时滞时间存在差异,这主要与木质部的结构、树干和冠层的水分储备、环境条件和树木个体在群落中所处的空间位置有关[22],而且还受树高、地径、冠幅、边材面积以及夜间水分补充量的影响[29]。
在整个生长季,杉木、马尾松和木荷树干液流速率与空气温度、太阳净辐射、水汽压亏缺呈极显著正相关,与空气相对湿度呈极显著负相关,而且太阳净辐射是影响3树种树干液流的主导环境因子。黄德卫等[13]对鼎湖山马尾松和木荷、梅婷婷等[19]对木荷优势木的研究中也得出过类似结论。这是因为蒸腾作用首先决定于植物自身的生理特性,太阳辐射直接影响植物的光合生理特性,在太阳辐射强烈的时候,植物的光合作用加强,各项生理指标都比较活跃,因此树干液流也明显较大。而郑怀舟等[18]则发现福建长汀马尾松树干液流与太阳辐射的相关性较其他因子小,究其原因可能与液流和太阳辐射间3 h的时滞有关。然而,树木作为一个独立的个体的同时,本身也是一个开放的系统,外界环境条件的波动会在树体内部的液流上反映出来,再加之树种、树木生长季节和地域条件等因素的差异,在不同的研究中往往得出不同结论。张宁南等[20]认为加勒比松液流主要是由蒸汽压差决定的,周国逸等[30]认为空气温度和相对湿度是影响液流的2个重要环境因子。此外,影响树干液流的环境因子会随土壤水分条件[31]、天气条件[21]、树木所处的空间位置[32-33]、观测部位[19,34]以及研究时间尺度[35]的变化而产生差异。
致谢:感谢中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室千烟洲生态站王辉民研究员和杨凤亭老师为本论文基础数据的采集提供帮助。
[1] 蒋芳市,黄炎和,钟炳林,等.不同治理措施对侵蚀红壤区生物量及生产力的影响[J].水土保持通报,2009,29(5):12-16.
[2] 王芸,欧阳志云,郑华,等.南方红壤区3种典型森林恢复方式对植物群落多样性的影响[J].生态学报,2013,33(4):1204-1211.
[3] 江玉梅,陈成龙,徐志红,等.退化红壤区人工林土壤的可溶性有机物、微生物生物量和酶活性[J].应用生态学报,2010,21(9):2273-2278.
[4] 黄荣珍,樊后保,李凤,等.人工修复措施对严重退化红壤固碳效益的影响[J].水土保持通报,2010,30(2):60-64.
[5] 涂洁,裴青宝,王辉民,等.江西退化红壤区枫香液流特征及其与气象因子的关系[J].浙江大学学报(农业与生命科学版),2013,39(5):575-581.
[6] Wullschleger S D,Norby R J.Sap velocity and canopy transpiration in a sweetgum stand exposed to free-air CO2enrichment(FACE)[J].New Phytologist,2001,150:489-498.
[7] Madurapperuma W S,Bleby T M,Burgess S S O,etal.Evaluation of sap flow methods to determine water use by cultivated palms[J].Environmental and Experimental Botany,2009,66:372-380.
[8] 孙慧珍,周晓峰,康绍忠.应用热技术研究树干液流进展[J].应用生态学报,2004,15(6):1074-1078.
[9] 黄玉清,张中峰,何成新,等.岩溶区青冈栎整树蒸腾的季节变化[J].应用生态学报,2009,20(2):256-264.
[10] 蒋文伟,汤富彬,刘志梅,等.天目山柳杉古树的液流特征研究[J].林业科学研究,2012,25(1):58-64.
[11] 王华,赵平,蔡锡安,等.马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应[J].应用生态学报,2008,19(2):225-230.
[12] 周翠鸣,黄玉清,顾大形,等.尾巨桉树干木质部液流密度径向变化特征[J].生态学杂志,2015,34(8):2103-2108.
[13] 黄德卫,张德强,周国逸,等.鼎湖山针阔叶混交林优势种树干液流特征及其与环境因子的关系[J].应用生态学报,2012,23(5):1159-1166.
[14] 孙振伟,赵平,牛俊峰,等.外来引种树种大叶相思和柠檬桉树干液流和蒸腾耗水的季节变异[J].生态学杂志,2014,33(10):2588-2595.
[15] Lu P,Urban L,Zhao P.Granier’s Thermal Dissipation Probe(TDP) method for measuring sap flow in trees:Theory and practice[J].Acta Botanica Sinica,2004,46(6):631-646.
[16] 赵平,饶兴权,马玲,等.马占相思(Acaciamangium)树干液流密度和整树蒸腾的个体差异[J].生态学报,2006,26(12):4050-4058.
[17] 赵仲辉,康文星,田大伦,等.湖南会同杉木液流变化及其与环境因子的关系[J].林业科学,2009,45(7):127-132.
[18] 郑怀舟,朱锦懋,李守中,等.长汀水土流失区生长季马尾松树干液流密度特征[J].亚热带资源与环境学报,2007,2(2):21-29.
[19] 梅婷婷,王传宽,赵平,等.木荷树干液流的密度特征[J].林业科学,2010,46(1):40-47.
[20] 张宁南,徐大平,Jim M,等.雷州半岛加勒比松人工林旱季液流特征与耗水量研究[J].林业科学研究,2007,20(5):591-597.
[21] 朱亚,吕光辉,胡颖颖,等.艾比湖自然保护区胡杨树干液流特征研究[J].干旱区资源与环境,2013,27(11):69-73.
[22] 赵平,刘惠,孙谷畴.4种植物气孔对水汽压亏缺敏感度的种间差异[J].中山大学学报(自然科学版),2007,46(4):64-68.
[23] Chuang Y L,Oren R,Bertozzi A L,etal.The porous media model for the hydraulic system of a conifer tree: Linking sap flux data to transpiration rate[J].Ecological Modelling,2006,191:447-468.
[24] Granier A,Bimn P,Lemine D.Water balance,transpiration and canopy conductance in two beech stands[J].Agricultural and Forest Meteorology,2000,100:291-308.
[25] 刘鑫,张金池,汪春林,等.长三角地区典型树种杉木液流速率变化特征[J].南京林业大学学报(自然科学版),2014,38(2):86-92.
[26] 王慧梅,孙伟,祖元刚,等.不同环境因子对兴安落叶松树干液流的时滞效应复杂性及其综合影响[J].应用生态学报,2011,22(12):3109-3116.
[27] Oguntunde P G.Whole plant water use and canopy conductance of cassava under limited available soil waterand varying evaporative demand[J].Plant and Soil,2005,278:371-383.
[28] Ford C R,Goranson C E,Mitchell R J,etal.Diurnal and seasonal variability in the radial distribution of sap flow: Predicting total stem flow inPinustaedatrees[J].Tree Physiology,2004,24:951-960.
[29] 王华,欧阳志云,郑华,等.北京城区常见树种生长季树干液流的时滞特征[J].应用生态学报,2009,20(9):2111-2117.
[30] 周国逸,尹光彩,Jim M,等.华南桉树人工林树液流通量及蒸散作用[J].植物学报,2004,46(2):202-210.
[31] 张振振,赵平,倪广艳,等.华南丘陵植被恢复先锋树种木荷与马占相思的水分利用[J].应用生态学报,2014,25(4):931-939.
[32] 熊伟,王彦辉,于澎涛,等.华北落叶松树干液流的个体差异和林分蒸腾估计的尺度上推[J].林业科学,2008,44(1):34-40.
[33] 刘军,陈文荣,徐金良,等.毛红椿人工林树干液流动态变化对坡位的响应[J].应用生态学报,2014,25(8):2209-2214.
[34] 胡伟,闫美杰,杜峰,等.黄土丘陵区山杨展叶期树干液流分析[J].水土保持学报,2010,24(4):48-52,58.
[35] 王文杰,孙伟,邱岭,等.不同时间尺度下兴安落叶松树干液流密度与环境因子的关系[J].林业科学,2012,48(1):77-85.
Comparisons on Sap Flow Characteristics of Three Dominant Tree Species from Mixed Conifer-broadleaf Forest in Degraded Red Soil Area
TU Jie,WANG Yongwen
(Research Institute of Ecology & Environmental Sciences,Nanchang Institute of Technology,Nanchang 330099,China)
Sap flow velocity(v) of three dominant species(Cunninghamialanceolata,PinusmassonianaandSchimasuperba) from a mixed conifer-broadleaf forest in degraded red soil area was continuously monitored by using Granier’s thermal dissipation probe method in this study.In the meanwhile, related meteorological factors including net solar radiation(NR),air temperature(Ta) and relative air humidity(RH) were simultaneously recorded by automatic weather station nearby.The aims were to evaluate the ability of utilizing and adjusting environmental water available by re-vegetation species, and ultimately to provide theoretical basis for tree selection in restoration area.The results were as follows: On the sunny days during the growing season,sap flow velocities of the three species exhibited similar single-peak diurnal patterns,with high values in the daytime and low values at night.But there were differences among them in the time of starting, reaching peak and the value of sap flow velocity.The sap flow ofSchimasuperbastarted 0.5 h before that ofCunninghamialanceolataandPinusmassoniana,and the daily averagevofSchimasuperbawas about 4 times and 1.5 times of that inCunninghamialanceolataandPinusmassoniana, respectively.On the rainy days,diurnal variations ofvdisplayed multiple-peaked curves and the values were distinctly lower than those on the sunny days.A distinct time lag was observed between the diurnal variations of sap flow velocity and NR as well as vapor pressure deficit(VPD), with sap flow lagging behind NR but being ahead of VPD.Correlation analysis results showed that sap flow velocity was positively correlated with NR,Taand VPD,whereas negatively correlated with RH.In case of the time lag effect considered, multiple-linear regression equations were established with successive sweep method representing the relationships between sap flow velocity and meteorological factors.All regression models indicated good fitting results with allR2more than 0.8,which met the significance at 0.01 levels withFtest.During the entire growing season,Ta,NR and RH could jointly explain 81.3%—92.2% of the variation in sap flow velocity ofCunninghamialanceolata.At the same time, 91.9%—93.6% of the variation in sap flow velocity ofPinusmassonianacould be explained by NR and VPD together.ForSchimasuperba,NR and VPD could explain 72.1%—92.5% of the variation in sap flow velocity from April to July.From August to October,80.7%—92.7% of the variation in sap flow velocity could be explained by NR and RH.Therefore,NR was considered to be the leading factor affecting sap flow velocity throughout the growing season.
sap flow; degraded red soil area;Cunninghamialanceolata;Pinusmassoniana;Schimasuperba; meteorological factors
2016-06-07
国家自然科学基金项目(31260172);江西省自然科学基金项目(20151BAB214006)
涂 洁(1980-),女,江西南昌人,副教授,博士,主要从事森林碳-水耦合机制研究。E-mail:tujie8058@163.com
时间:2016-11-25 14:24:33
S791.27
A
1004-3268(2016)12-0120-07
网络出版地址:http://www.cnki.net/kcms/detail/41.1092.S.20161125.1424.031.html
