汉中主栽柑橘品种对低温胁迫的生理响应
2016-02-06蒋景龙
蒋景龙
(陕西理工学院 生物科学与工程学院,陕西 汉中 723001)
汉中主栽柑橘品种对低温胁迫的生理响应
蒋景龙
(陕西理工学院 生物科学与工程学院,陕西 汉中 723001)

柑橘; 低温胁迫; 生理响应; 隶属函数; 抗寒能力
柑橘(Citrusreticulata)为芸香科柑橘亚属常绿小乔木或灌木,主要分布在热带和亚热带地区,性喜温暖湿润气候。陕西省汉中柑橘栽培历史悠久,距今已有2 000多年历史,柑橘已成为汉中农业主导产业之一,是当地农业收入的重要来源之一[1]。然而,汉中地区地处亚热带的最北缘,也是中国种植柑橘的最北缘,属柑橘冻害的多发地带,近几年陕南柑橘连续遭受周期性大面积冻害。2012—2013年冬持续的低温天气,致陕南柑橘产区再次遭遇近30 a来最严重冻害,受冻面积达10 672 hm2,占全省柑橘种植总面积的25%,汉中市的城固县等主产区受灾尤为严重[2]。
低温冻害是影响柑橘生产的常发性自然灾害,极端地区甚至年年发生。柑橘一般只能抵抗-3 ℃左右的低温,当最低温度低于-5 ℃时,极易发生冻害,冻害程度还与低温持续时间有关[3]。低温胁迫对柑橘生长发育、膜系统、渗透调节物质和抗氧化系统等生理生化方面均造成一定影响[4]。李卫等[5]研究5种柑橘原生质体含量和田间植株的叶片抗寒性后,发现原生质体含量与田间植株正常叶片抗寒性呈正相关。周晓音等[6]通过检测叶片荧光参数来确定6个柑橘品种的耐冻性强弱,结果发现,甜橘柚Fm和Fv/Fm的变化最大,耐低温胁迫能力最弱,而宫川温州蜜柑的Fm和Fv/Fm的变化不显著,耐冻性最强。马文涛等[7]以贵州3个野生柑橘的1年生枝条为试材,低温冷冻处理24 h后检测丙二醛(MDA)含量、可溶性糖含量、过氧化物酶(POD)和超氧化物歧化酶(SOD)活性等生理指标,结果表明抗寒性强的柑橘枝条具有更强的抗氧化胁迫能力和渗透调节能力。张颖等[8]经过反复试验, 发现在人工控制条件下生长部位一致、年龄相同的离体叶片叶绿素含量、膜透性和细胞生长状况等生理特性在同一基因型内的变化很小,可以准确鉴定杨树的耐盐潜力。曾小美等[9]研究显示,拟南芥连体和离体叶片光合作用光响应结果一致。李自龙等[10]采用不同品种油橄榄的离体叶片来研究渗透胁迫下的生理响应及其抗旱机制;同时,令凡等[11]采用不同品种油橄榄的离体叶片研究其对低温胁迫的生理响应。本研究以汉中地区引种的抗冻性较强的品种日南1号和兴津及本地培育并推广种植的抗冻性相对较差品种泛亚特早的离体叶片为材料,分析-5 ℃低温胁迫下离体叶片中与抗寒性相关的生理指标,并通过主成分分析法评价各个测定指标的贡献率,筛选柑橘抗寒性分析的重要指标,以期为汉中地区柑橘抗寒品种的筛选和鉴定提供重要参考。
1 材料和方法
1.1 材料培养与处理
参试的兴津、日南1号和泛亚特早3个柑橘品种均由汉中泛亚绿色食品有限公司苗圃基地提供,3种橘苗均为1年生发育良好的嫁接苗(砧木为枸橘,穗木分别为3个品种的1年生穗条)。将3种盆栽橘苗先在人工气候培养箱内预培养,温度为25 ℃/18 ℃(昼/夜),空气相对湿度为80%,光照时间为16 h/8 h(昼/夜),光照强度为4 000 lx。培养2周后,选取生长一致橘苗,摘取植株中部健康且大小一致的柑橘叶片,置于-5 ℃的冰箱分别低温胁迫3 h、6 h、9 h、12 h,以处理前25 ℃/18 ℃(昼/夜)人工气候培养箱培养的柑橘离体叶片为对照,处理温度误差为±0.5 ℃,所有指标均设置3个重复。
1.2 测定项目与方法
1.2.1 叶片相对电导率(REC)的测定 对照组和处理组的柑橘叶片分别用自来水和蒸馏水冲洗3次,去离子水冲洗1次后用滤纸吸干表面水分,将叶片避开主脉剪成长条,快速称取3份,每份0.1 g,分别置于含20 mL去离子水的刻度试管中,室温下浸泡处理12 h,用雷磁DDSJ-308F型电导率仪测定叶片浸提液电导率,记为R1;然后将装有叶片浸提液的试管放入水浴锅沸水浴30 min,冷却至室温后摇匀,再次测定浸提液电导率,记为R2,计算叶片REC,REC=(R1/R2)×100%。
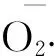
1.3 隶属函数与各单项指标的主成分分析
各指标的隶属函数分析参照孟艳琼等[16]和司剑华等[17]的方法。参考廖景容等[18]的方法,隶属度按5级制划分标准:隶属度≥0.7为强抗(Ⅰ级);0.6≤隶属度<0.7为抗(Ⅱ级);0.4≤隶属度<0.6为中抗(Ⅲ级);0.3≤隶属度<0.4为弱抗(Ⅳ级);隶属度<0.3为不抗(Ⅴ级)。抗寒系数=低温胁迫后指标测定值/对照组指标测定值×100%,各单项指标的主成分分析参照刘杜玲等[19]的方法。
1.4 数据分析
试验数据采用Excel 2007、Origin 7.5 SR1和SPSS 16.0软件进行制图和统计分析。
2 结果与分析
2.1 不同低温胁迫下3种柑橘离体叶片的形态变化
如图1所示,与对照组相比,在-5 ℃处理3 h时兴津与日南1号叶片无明显变化(图1B、G),泛亚特早叶片开始卷曲(图1L);处理6 h时,兴津和日南1号叶片仍无明显变化(图1C、H),泛亚特早叶片卷曲严重(图1M);当处理时间持续到9 h时,兴津和日南1号叶片开始卷曲(图1D、I),泛亚特早叶片开始渗水、萎蔫(图1N);低温处理时间持续到12 h时,3种柑橘的叶片均开始出现不同程度的渗水萎蔫,其中日南1号最轻(图1J),兴津次之(图1E),泛亚特早最为严重(图1O)。以上3种柑橘叶片在不同低温胁迫下的形态变化表明,日南1号抗冻性较强,兴津次之,泛亚特早最弱。

A—E为兴津品种叶片; F—J为日南1号品种叶片; K—O为泛亚品种叶片; A、F、K为对照组叶片; B、G、L为-5 ℃处理3 h叶片;C、H、M为-5 ℃处理6 h叶片; D、I、N为-5 ℃处理9 h叶片; E、J、O为-5 ℃处理12 h叶片图1 不同低温胁迫时间下3种柑橘叶片形态变化
2.2 低温胁迫对柑橘离体叶片的伤害程度
2.2.1 REC 3种柑橘离体叶片REC均随低温胁迫时间的持续而逐渐升高(图2)。与对照组相比,处理3 h时,泛亚特早叶片REC显著升高而兴津和日南1号无显著性变化(P>0.05);处理6 h时,泛亚特早和兴津叶片REC分别较对照显著升高16.96%和9.28%(P<0.05),而日南1号叶片REC虽升高了1.19%,但差异不显著(P>0.05);处理9 h及以后,3种柑橘离体叶片REC均较对照显著性升高 (P<0.05),泛亚特早、兴津和日南1号离体叶片REC在处理12 h时分别比对照显著升高58.14%、39.98%和19.27%。可见,3种柑橘离体叶片REC(与对照组相比)出现显著性升高的时间依次为泛亚特早3 h、兴津3 h和日南1号9 h,由此可以推断在-5 ℃低温胁迫下离体叶片细胞膜受到的伤害程度也以泛亚特早最大,兴津次之,而日南1号最小。

不同字母表示同一品种低温处理不同时间在0.05水平差异显著。下图同
2.3 低温胁迫对柑橘离体叶片渗透调节物质含量的影响
由表1可见,在-5 ℃低温胁迫条件下,兴津和泛亚特早离体叶片中Pro含量均在处理6 h时开始显著升高,处理9 h时达到最大值,此时分别比对照显著提高53%和42%(P<0.05);而日南1号则从处理3 h时开始显著升高,处理6 h时达到最大值(P<0.05),此时比对照显著提高66%。即日南1号比其他2个品种Pro含量显著升高的时间要早,升高的幅度更大。同时,3种柑橘叶片中可溶性糖含量随着低温胁迫时间延长均呈逐渐升高趋势,均在处理12 h时达到最大值,此时泛亚特早、兴津和日南1号离体叶片分别比对照显著提高34%、48%和92%(P<0.05);但各品种开始出现显著性升高的时间有差异性,日南1号、兴津和泛亚特早分别在处理6 h、9 h和12 h时出现显著性升高。以上3个品种离体叶片2种渗透调节物质含量的变化表明,日南1号渗透调节物质积累对低温胁迫的响应更敏感、更强烈,在低温逆境下的渗透调节能力明显强于另外两品种,泛亚特早则最弱。
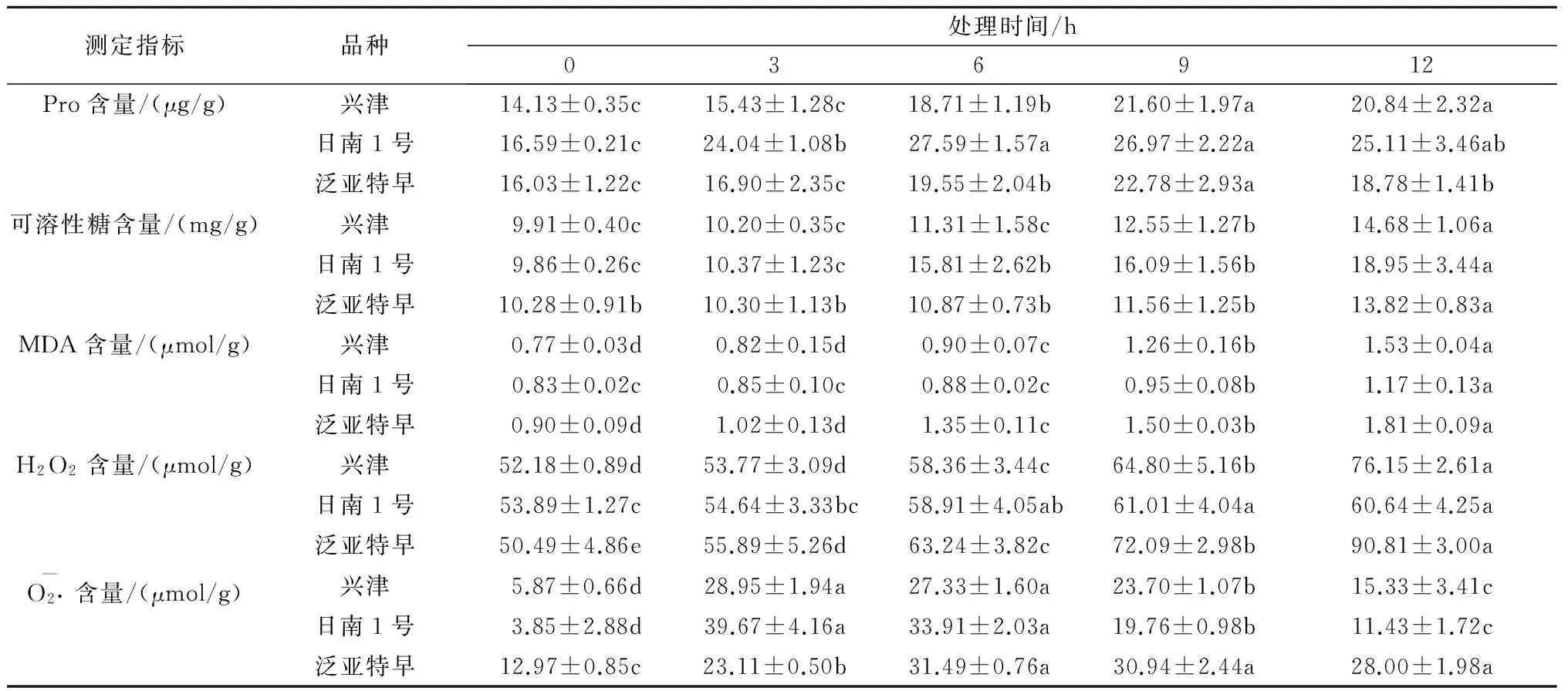
表1 低温胁迫不同时间3种柑橘叶片渗透调节物质、MDA和活性氧水平的变化
注:同行不同字母表示不同处理时间在0.05水平存在显著性差异。
2.4 低温胁迫对柑橘离体叶片抗氧化酶活性的影响 图3显示,随着低温胁迫的持续,泛亚特早和兴津离体叶片SOD活性呈现出先升高后降低的趋势,并均在处理6 h时达到最高,在处理9 h时开始下降,而日南1号则呈现持续显著升高的变化趋势(P<0.05),泛亚特早、兴津和日南1号三者的最高值分别比对照显著提高57%、112%和115%。同时,随着低温胁迫持续,3个品种柑橘离体叶片CAT活性均呈现先升高后降低的变化趋势,其中的泛亚特早和兴津均在处理6 h时达到最高值,随后开始下降,而日南1号则在处理9 h时达到最高值随后也开始下降,三者的最高值分别比对照显著提高34%、23%和97%。另外,泛亚特早和兴津离体叶片中的POD和APX活性也随着低温胁迫持续而呈现出先升高后降低的趋势,且均在处理6 h时达到最高值随后开始下降;日南1号叶片POD活性在处理3 h时已升至最高,并保持到9 h后开始下降,而其叶片APX活性在处理6 h时升至最高,其后一直保持显著高于对照组的状态。泛亚特早、兴津和日南1号叶片POD活性峰值分别比对照显著提高77%、60%和101%,APX活性峰值分别比对照显著提高22%、50%和42%。可见,随着低温胁迫持续,泛亚特早和兴津离体叶片中的SOD、CAT、POD和APX活性均表现出先升高后降低的变化趋势,而日南1号叶片中的SOD和APX活性则几乎呈现出持续升高的趋势,其CAT和POD活性则表现出先升高后又降低的趋势,但比其他2个品种下降的时间较晚,一般在处理9 h后开始下降,而泛亚特早和兴津则在处理6 h后就开始下降;与其他品种相比,日南1号叶片抗氧化酶系统对低温胁迫的响应更迅速,活性更强且持续时间更长,表现出更强的活性氧清除能力。
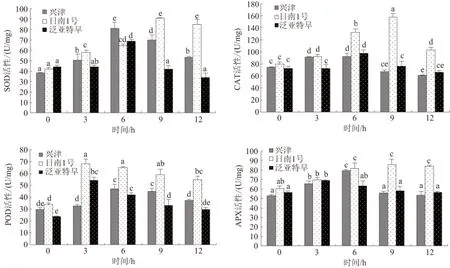
图3 低温胁迫不同时间3种柑橘叶片抗氧化酶的变化
2.5 抗寒性比较与综合指标贡献率分析
应用隶属函数法对各项生理指标进行综合比较3种柑橘抗寒能力的大小,其值越大则抗寒能力越强。平均隶属度分析表明,3种柑橘抗寒能力依次为日南1号>兴津>泛亚特早(表2)。3种柑橘平均隶属度均在0.4~0.6(表2),根据廖景容等[18]的隶属度5级制划分标准,抗寒性属于中抗等级。对3种柑橘10项生理指标的抗寒系数进行主成分分析,REC和Pro含量的综合指标贡献率分别为79.141%和18.045%,累积贡献率为97.186%(表2),一般累积贡献率大于85%具有较强的代表性,因此可以用这2个指标进行柑橘的抗寒性分析。
表2 低温胁迫下柑橘各指标隶属函数值及各综合指标
抗寒系数及贡献率

测定指标品种兴津日南1号泛亚特早综合指标RECPro含量REC0.6030.6850.542-0.995 0.096Pro含量0.4670.4830.3820.6230.782可溶性糖含量0.3890.4870.3140.990-0.143 MDA含量0.6310.6880.550-0.995 0.102O2·-含量0.3850.5060.3410.9990.048H2O2含量0.6370.6030.4250.9990.039SOD活性0.4810.5430.3730.8620.507CAT活性0.5190.4360.3550.909-0.417 POD活性0.5040.6480.4220.463-0.886 APX活性0.3310.6260.3340.9930.118平均隶属度0.4950.5710.404抗寒性排序213贡献率/%79.14118.045累计贡献率/%79.14197.186
3 结论与讨论
叶片是植物进行光合作用和呼吸的主要器官,同时也是冻害最容易侵袭的部位。重冻时,柑橘叶片呈桶状卷曲,很快萎凋干枯,落叶严重,光合作用停止,轻冻时可使柑橘叶片表皮细胞受害,蒸腾作用加强,叶片慢慢卷缩或部分落叶,光合作用减弱。该研究中-5 ℃处理12 h过程中,3种柑橘叶片均出现了不同程度的叶片卷曲和渗水萎蔫现象,但泛亚特早低温处理3 h即开始出现卷曲,时间较早,后期卷曲较为严重,而日南1号和兴津在处理9 h开始出现叶片卷曲。在低温处理12 h时3种柑橘均出现不同程度渗水萎蔫,泛亚特早最为严重,日南1号最轻。REC是反映植物受逆境胁迫后细胞膜透性变化的重要生理指标,与植物的抗寒能力呈负相关,低温胁迫下抗寒能力弱的植物膜系统稳定性较差,细胞内容物外渗量大,REC较高[20]。本研究中3种柑橘离体叶片的REC随低温胁迫的持续呈逐渐上升趋势,表明低温胁迫时间越长,柑橘离体叶片细胞膜受伤害越严重。马翠兰等[21]对蜜柚进行-5 ℃处理,发现随着低温处理的持续,叶片REC持续增加,处理12 h时REC达到51.94%,处理24 h时REC急剧增至92.46%。本研究中-5 ℃处理12 h泛亚特早叶片的REC已达到82.39%,兴津为57.78%,均超过了50%,而日南1号为36.32%。令凡等[11]认为低温胁迫后当REC超过50%,叶片细胞膜受到严重伤害;REC未超过50%,细胞膜透性受到轻度伤害,并且REC与植物的抗寒能力呈负相关。由此可见,当-5 ℃处理持续12 h时,泛亚特早和兴津叶片细胞膜已收到严重伤害,但泛亚特早最为严重,而日南1号仍处于轻度伤害,日南1号抗寒性最强,兴津次之,泛亚特早最弱。植物的抗寒能力与MDA含量呈负相关,即抗寒能力越强,MDA含量越小[22]。本试验结果表明,各柑橘离体叶片MDA含量随低温的持续呈增加趋势,它们MDA含量相对增幅表现为日南1号<兴津<泛亚特早,再次证明日南1号遭受低温伤害的程度小,抗寒性较其他2个品种强。
Pro和可溶性糖的积累提高了细胞渗透势,减少了水分的流失,还能直接与细胞组分分子相连,从而对细胞膜和酶起到稳定作用[23]。研究中,随着低温胁迫的持续,柑橘离体叶片的渗透调节物质Pro和可溶性糖含量均增加,且日南1号叶片渗透调节物质的积累显著高于泛亚特早和兴津,其较强的渗透调节能力在一定程度上提高了其抗寒能力。邵怡若等[24]的研究结果显示,随着低温胁迫时间的持续,盐肤木和假连翘幼苗的Pro含量均持续上升,葛藤幼苗的Pro含量上升后趋于平稳,除了盐肤木叶片的可溶性糖含量显著上升后小幅波动,其余幼苗可溶性糖含量均持续增加,这与本研究结果相似。本研究中柑橘叶片SOD、CAT、POD和APX活性随低温的持续总体上呈先升后降的趋势,说明低温胁迫起初诱发抗氧化酶活性上升,但随着胁迫加剧,这些抗氧化酶活性下降,即严重的低温胁迫积累了过量的活性氧,使活性氧防御系统的动态平衡遭到了破坏,从而加剧了膜脂过氧化作用[25]。严岳华等[26]发现,-5 ℃低温处理2 d,6种观赏柑橘叶片的CAT活性均呈上升的趋势。武雁军等[27]对低温胁迫下的厚皮甜瓜幼苗生理指标检测结果显示,随低温处理时间的延长,2种厚皮甜瓜的SOD、POD和CAT活性均上升,但抗寒性强的品种SOD、POD和CAT等保护酶活性更高。植物的抗寒性受多种生理因素的影响,综合抗寒性评价更能保证结果的可靠性[28-29]。本研究综合抗寒性评价结果表明,日南1号的抗寒能力较泛亚特早和兴津强,可用于抗寒品种的改良和筛选,生产中可作为主栽品种。泛亚特早和兴津的抗寒性较差,栽培管理中应注意防寒措施,如合理施用有机肥、采果后喷叶面肥、坡地橘园在降霜期进行树冠喷水等措施均有助于提高树体抗寒能力。对3种柑橘的10项生理指标的抗寒系数进行主成分分析,REC和Pro含量的综合指标的累积贡献率大于85%,具有较强的代表性,可以作为柑橘的抗寒性分析的重要指标。
[1] 张永平,万恩梅,王永琦.汉中市柑橘生产存在的问题及对策[J].陕西农业科学,2011,57(5):163-164.
[2] 李新生,江海,郭念文.陕西柑橘资源与产业化开发研究[M].北京:科学出版社,2014:11-76.
[3] 朱海涛,周勍,巫丽君,等.镇江市柑橘冻害研究[J].江苏农业科学,2012,40(2):274-277.
[4] 陈志远,邸丽俊,王国栋,等.柑橘冻害抵抗分子机理研究进展[J].北方园艺,2014,38(1):188-191.
[5] 李卫,孙中海,章文才,等.柑橘抗寒育种早期鉴定的一种指标[J].植物学报,1998,40(9):827-830.
[6] 周晓音,饶建民,叶伟其.低温对柑橘叶片荧光参数的影响[J].浙江柑橘,2010,27(3):9-11.
[7] 马文涛,樊卫国.贵州野生柑橘的抗寒性测定和综合评价[J].西北植物学报,2014,34(10):2063-2069.
[8] 张颖,杨迎霞,郏艳红,等.利用离体叶片鉴定杨树耐盐潜力[J].植物学报,2011,46(3):302-310.
[9] 曾小美,袁琳,沈允钢.拟南芥连体和离体叶片光合作用的光响应[J].植物生理学通讯,2002,38(1):25-26.
[10] 李自龙,徐雪风,焦健,等.不同品种油橄榄离体叶片对渗透胁迫的生理响应及其抗旱机制[J].西北植物学报,2014,34(9):1808-1814.
[11] 令凡,焦健,李朝周,等.不同油橄榄品种对低温胁迫的生理响应及抗寒性综合评价[J].西北植物学报,2015,35(3):508-515.
[12] Jiang J L,Su M,Chen Y R,etal.Correlation of drought resistance in grass pea(LathyrussativusL.)with reactive oxygen species scavenging and osmotic adjustment[J].Biologia,2013,68(2):231-240.
[13] Farooq M,Basra S M A,Wahid A,etal.Improving the drought tolerance in rice(OryzasativaL.) by exogenous application of salicylicacid[J].Journal of Agronomy and Crop Science,2009,195:237-246.
[14] 李玲,李娘辉,蒋素梅,等.植物生理学实验技术[M].北京:科学出版社,2014:83-104.
[15] Jiang J L,Su M,Wang L Y,etal.Exogenous hydrogen peroxide reversibly inhibits root gravitropism and induces horizontal curvature of primary root during grass pea germination[J]. Plant Physiology and Biochemistry,2012,53:84-93.
[16] 孟艳琼,张令峰,王雷宏,等.低温胁迫对6种彩叶藤本植物抗寒性生理指标的影响[J].安徽农业大学学报,2009,36(2):172-177.
[17] 司剑华,卢素锦.低温胁迫对5种柽柳抗寒性生理指标的影响[J].中南林业科技大学学报,2010,30(8):78-81.
[18] 廖景容,郑中阳,张卫星,等.玉米不同杂交组合抗旱性、丰产性的研究[J].安徽农业科学,2005,33(1):7-10.
[19] 刘杜玲,张博勇,孙红梅,等.早实核桃不同品种抗寒性综合评价[J].园艺学报,2015,42(3):545-553.
[20] 武辉,张巨松,石俊毅,等.棉花幼苗对不同程度低温逆境的生理响应[J].西北植物学报,2013,33(1):74-82.
[21] 马翠兰,刘星辉,胡又厘.柚品种间的耐寒性差异及其机理[J].福建农业大学学报,1998,27(2):160-165.
[22] 陈颖,陈昕,汪南阳,等.低温胁迫下西番莲叶片的生理反应及超微结构变化[J].西北植物学报,2012,32(3):532-539.
[23] 李娜,房伟民,陈发棣,等.切花寒菊小花对低温胁迫的生理响应及其抗寒性分析[J].西北植物学报,2010,30(4):741-746.
[24] 邵怡若,许建新,薛立,等.低温胁迫时间对4种幼苗生理生化及光合特性的影响[J].生态学报,2013,33(14):4237-4247.
[25] 张军,孙树贵,王亮明,等.孕穗期低温对冬小麦生理生化特性和产量的影响[J].西北植物学报,2013,33(11):2249-2256.
[26] 严岳华,何少波,李觅路.低温胁迫对观赏柑桔CAT活性与水势的影响[J].湖南农业科学,2005(4):104-106.
[27] 武雁军,刘建辉.低温胁迫对厚皮甜瓜幼苗抗寒性生理生化指标的影响[J].西北农林科技大学学报(自然科学版),2007,35(3):139-143.
[28] 南丽丽,师尚礼,陈建纲,等.不同根型苜蓿根系对低温胁迫的响应及其抗寒性评价[J].中国生态农业学报,2011,19(3):619-625.
[29] 吕优伟,贺佳圆,白小明,等.9个野生早熟禾对低温胁迫的生理响应及苗期抗寒性评价[J].草地学报,2014,22(2):326-333.
Research on Physiological Response to Low Temperature Stress ofCitrusreticulataCultivars in Hanzhong
JIANG Jinglong
(School of Biological Science and Engineering,Shaanxi University of Technology,Hanzhong 723001,China)

Citrusreticulata; low temperature stress; physiological response; subordinate function; cold resistance ability
2015-11-19
陕西省科技厅自然科学基础研究计划项目(2014JQ3113);陕西省教育厅自然科学研究项目(14JK1158);陕西理工学院科研基金项目(SLGKY15-41)
蒋景龙(1980-),男,山东枣庄人,讲师,博士,主要从事逆境植物学方面的研究。 E-mail:jiangjinglong511@163.com
S666
A
1004-3268(2016)03-0106-07
