外显与内隐刻板印象的分布式表征及其激活过程*
——基于认知神经科学视角的探索
2016-02-01祝书荣张常洁张庆林
贾 磊 祝书荣 张常洁 张庆林
(1浙江工业大学心理系, 杭州 310023) (2西南大学心理学部, 重庆 400715)
1 问题与分析
1.1 传统社会认知理论下的内隐和外显刻板印象研究
1.1.1 内隐社会认知视角下的内隐和外显刻板印象
传统社会认知研究将刻板印象视作是联结某个特定群体与其行为特征和特质属性的特殊社会知识。这种社会知识通常以语义形式编码和储存,但却具有极其复杂的图式结构——它既是对群体的特质概括, 具有表征和解释群体范畴的知识属性;也是对群体进行自发社会分类所持有的内在信念,稳定而持久, 一经形成即难以发生改变(Hamilton &Trolier, 1986; Hilton & von Hippel, 1996)。这种复杂的语义图式结构决定了刻板印象激活(stereotyping)形式的多样性:一方面, 它既能在特定线索刺激驱动下, 以内隐的语义启动形式自动激活群体与特质行为间的知识联结; 另一方面, 它也能在期望和意图引导下, 以外显的概念驱动形式激活刻板的图式信念并由此做出社会判断。
为了明确个人主观意图与意识水平对刻板印象的调制影响, 内隐社会认知理论根据刻板印象激活方式的差异, 将其分为了两类表现型, 一类是能够被意识内省察觉的外显刻板印象, 另一类则是难以通过意识内省即时觉察的内隐刻板印象。其中, 前者体现的是个体对某一群体持有的社会观念, 它多以语义图式形式表征于概念系统,并以可控的概念驱动形式激活表达于意识层面。而后者指“无法或难以内省辨别, 但却会对社会群体及其成员的属性判断产生影响的过去经验痕迹”(Greenwald & Banaji, 1995), 它所体现的是过去经验对于群体认识所施加的潜在影响。相比意识概念化的外显刻板印象, 内隐刻板印象的激活处于前意识水平, 不依赖于特定的目的意图, 无需意识努力, 且过程更为流畅。反映在行为测量上, 外显刻板印象多采用直接外显的概念测量手段, 例如给予对象群体以自由的特质或印象描述(自由反应法), 从特质词表中选出最符合目标群体的特质(Katz-Braly法), 或是呈现对象群体后, 要求判断随后出现的特质词与其是否相符(“分类−确认”任务)。内隐刻板印象则多采用间接性的内隐测量手段, 如序列启动任务、内隐联想测验(Implicit Association Test, IAT)、外在情感西蒙任务(Extrinsic Affective Simon Task, EAST)、Go/Nogo任务、刻板解释偏差(Stereotypic Explanatory Bias, SEB)等。
内隐社会认知理论认为, 以 IAT等间接测量手段测得的内隐认知成分具有加工流畅、自动化,且不依赖于意识努力等特点, 因此机制上有别于具有精细编码和意义组织特性的外显认知成分。但对于该成分的实质, 内隐社会认知理论内部却产生了观点分歧。争论的焦点在此内隐认知成分究竟反映的是稳定的记忆表征, 还是建立在环境场合基础之上的背景依赖性的评估或评价, 由此形成了两种不同的理论取向(Payne & Gawronski, 2010)。
1.1.2 内隐与外显刻板印象机制探索的两种理论取向
内隐社会认知理论诞生的实证基础主要来自两个领域。首个领域是关于选择性注意资源分配的研究。这类研究最早提出了自动加工与控制加工的概念。这种双加工观点为内隐社会认知所继承, 进而演化为解释社会认知内隐/自动加工与外显/控制加工的一般性双加工理论。另一个领域则是内隐记忆的相关研究。此类研究在记忆层面区分了无意识的内隐记忆和有意识参与的外显记忆。这种内隐记忆的观点同样为内隐社会认知理论吸收, 转而用于解释和强调内隐社会认知有别于外显社会认知的独特记忆基础(Payne & Gawronski,2010)。具体至内隐和外显刻板印象研究中, 研究视角的不同间接导致了研究观点的分化, 由此形成了两种不同的理论探索取向。
第一种理论取向是将刻板印象的加工过程视作是一种建立在个人需要和背景场合基础上的社会评估或评价过程, 代表理论即是 Devine (1989)提出的双加工理论(Dual-process Model)。该理论将刻板印象加工过程分解为刻板信念的自动激活阶段(据Automatic简称A阶段), 以及根据个人意图和环境背景决定运用或是控制刻板印象的控制加工阶段(据Control简称C阶段)。根据该理论的早期观点, 内隐与无意识、自动加工等价, 而外显与有意识、控制加工等价。因此, 刻板印象内隐激活只出现在A阶段。而当刻板印象进入意识控制的C阶段, 个体须根据外在的环境需要和内在的动机意图, 选择接受、忽略或是控制已经激活的刻板信念时, 才会形成外显的刻板印象运用或是控制。根据这一观点, 比较和分析外显和内隐刻板印象的机制差异, 实质是将外显刻板印象运用或控制(A+C两阶段)同内隐刻板印象激活(仅A阶段)进行比较。这种“A+C”的两阶段划分虽可以解释刻板印象自动激活与控制加工两阶段过程中的资源动态分配, 但存在一定问题。首先, 这种两阶段的划分忽略了自由反应法、Katz-Braly法以及“分类−确认”任务这类概念驱动任务所直接激活的外显刻板印象, 难以将概念驱动激活的外显刻板印象同语义启动激活的内隐刻板印象加以比较, 也无法检验和比较二者的编码机制与记忆基础。其次, 由于早期研究视“内隐”为绝对的自动化和无意识, 因此内隐刻板印象激活也被视作是绝对、无条件的。但后来研究发现, 个体内在的动机倾向以及外在的背景和线索, 依然会影响内隐刻板印象的自动激活, 进而显示内隐加工亦是有条件且有意识控制参与的(Blair, 2002)。以Payne (2001)采用“武器−工具”辨别任务的内隐刻板印象实验为例, 在面对相同的内隐启动任务和刺激时, 同一个体在时间限制条件的错误辨别反应, 比无时间限制条件下的错误辨别反应要显现出更强的刻板偏差倾向。此时若将内隐等同自动无意识加工,否认认知控制的存在, 那么该启动任务下唯有内隐刻板印象自动激活一个加工阶段, 随即无法解释同一个体在相同内隐任务中所产生的行为差异。为此, 后期研究一方面提出了前意识自动化(Preconscious Automaticity)的概念, 以适应内隐自动过程的有条件性(Bargh, Schwader, Hailey, Dyer,& Boothby, 2012); 另一方面, 为了分离内隐刻板印象激活过程中自动成分与控制成分, 考察二者对内隐任务的各自贡献, 后期研究引入了过程分离程序(Process Dissociation Procedure, PDP)、四重模型(Quad-Model)等一批形式过程分析模型(Payne &Gawronski, 2010)。
另一种理论研究取向则更为关注内隐和外显刻板印象背后的记忆机制差异。此类研究认为内隐测量测得的认知成分实质反映的是区别于外显记忆的内隐记忆, 并据此将内隐与外显刻板印象的机制差异归结于内隐认知与外显认知的记忆基础差异。该取向的代表理论如Tulving (1985)提出的多重记忆系统模型(Multiple Memory Systems Model; 以下简称为MMS模型)。依照此模型观点,相比需要意识投入且以精细编码为主的外显记忆,内隐记忆具有自动激活且无须意识努力的特点。据此推论, 以语义形式编码和存储的刻板印象理应存在外显语义记忆和内隐语义记忆两种表征形态,从而以语义分布式的形式存在于长时记忆系统中。其中, 以外显语义记忆表征存储的外显刻板印象通过概念驱动形式予以激活, 而以内隐语义记忆表征存储的内隐刻板印象则通过语义启动形式予以激活。这一观点将MMS模型的“外显记忆−内隐记忆”假设作为理论核心, 成为解释外显和内隐刻板印象机制差异的另一种重要思想(Amodio & Ratner,2011)。需要指出的是, 虽然 MMS模型所认为的内隐记忆有着大量的证据支持, 但它仍不乏批评者。例如Roediger (1990)提出的迁移适当加工理论(Transfer-appropriate Processing Theory)即否认内隐记忆是一种独立的记忆形态。他认为外显测量的任务加工实质是依赖刺激项目语义(或意义)的概念驱动过程, 而内隐测量实际是对过去经验进行提取匹配的知觉驱动过程, 因此内隐和外显的机制差异实质是同一记忆系统下知觉匹配过程同概念评估过程的差异。总之, 外显与内隐刻板印象机制是否真如 MMS模型推测的那样, 传统的行为研究手段由于无法将知觉编码、认知评估过程与记忆存储加工进行机制分离, 并未提供足够的证据支持。
综上所述, 研究取向的不同直接导致了外显和内隐刻板印象的解释分歧——二者的机制差异既可解释为不同任务背景下相同刻板知识信念因知觉编码与认知评估机制不同所导致的加工差异,也可解释为刻板知识信念在内隐和外显记忆系统下的分布式表征。这种状况一直持续至认知神经科学的崛起。随着上述两种理论研究取向的各自发展, 一批重要实证发现与理论模型也随之诞生。这些研究成果从脑科学层面重新解读了刻板印象的认知实质, 也为解决外显和内隐刻板印象的机制差异问题提供了新的机遇。
1.2 刻板印象的分布式表征构想:多重记忆系统模型与分布式语义表征假设
相比传统行为手段所遭遇的“黑箱困境”, 现代认知神经科学技术的一大优势就是可以通过高空间分辨率的神经成像手段, 直接观测感兴趣脑区随任务发生的信号变化。因此就外显和内隐刻板印象的解释争论而言, 最直接有效的检验方式就是通过神经成像手段比较二者存在的记忆神经基础, 检验MMS模型的理论观点。
1.2.1 外显和内隐刻板印象的记忆解释:多重记忆系统神经模型
作为记忆的经典理论, MMS模型由 Tulving(1985)提出构想并由 Squire (2004)基于神经科学视角予以完善。该模型依据长时记忆的认知表现及其神经基础的不同, 将其分为外显和内隐两个子系统。其中, 外显记忆系统主要由内侧颞叶(medial temporal lobe, MTL)与间脑系统构成, 处理对象主要包括表征事件经历的情境记忆, 以及表征事实与客观知识的外显语义记忆。内隐记忆则形式更为多样, 包括程序性记忆、启动效应、简单条件反射和非联想性学习等多种类别。它所对应的神经系统构成也更为复杂, 与大脑新皮层、小脑以及皮层下的边缘系统均存在有广泛联系(Squire, 2004;Squire & Zola, 1996)。而依据该模型观点, 作为特殊语义知识的外显和内隐刻板印象, 它们应该分别表征于外显和内隐记忆系统。鉴于刻板印象以语义形式进行编码储存, 据此推论, 刻板印象表征涉及多个记忆系统, 而非单一记忆系统(Amodio& Ratner, 2011)。
1.2.2 刻板印象的语义分布式表征:多重记忆系统模型的假设与证据
在概念语义表征的解释上, MMS模型继承并发展了早期分布式语义表征理论的观点。这一理论可追溯至早年有关言语障碍的研究, 主张语义概念并非与记忆神经网络中的单个存储单位一一对应, 而是以多个单位共同参与的模式予以表征(Allport, 1985)。在此基础上, MMS模型主张将语义记忆分离为以事实和客观知识为主体的外显语义记忆, 和以语义联结形式存在的内隐语义记忆(Squire, 2004; Squire & Zola, 1996)。这种记忆分类得到了神经成像证据的支持。研究发现, 外显语义记忆的存储和提取, 与前额皮层、后部扣带回(posterior cingulate cortex, PCC)、颞叶后部的梭状回(fusiform gyrus)以及内侧颞叶构成的复杂神经网络有着紧密联系。而内隐语义记忆则通常以语义启动形式表征于前额及颞叶的新皮层, 同左半球额下回(inferior frontal gyrus, IFG)和外侧颞顶叶皮层构成的内隐语义启动通道有着直接联系(Binder & Desai, 2011; Golby et al., 2005; Rissman,Eliassen, & Blumstein, 2003)。在提取机制上, 外显和内隐语义记忆也存有明显差异。外显语义记忆具备主动提取能力, 此提取过程一般由额叶触发,且以意识参与的精细编码为主要特征。而内隐语义记忆没有主动提取过程, 但却带有自动激活与兴奋扩散的特性(Miyashita, 2004)。
脑成像研究观察到的刻板印象表征特性确实符合MMS模型的假设观点(Quadflieg & Macrae,2011)。一方面, ERP研究中发现了十分显著的刻板印象 N400效应, 且该效应既出现在外显的“分类−确认”任务下(王沛, 杨亚平, 赵仑, 2010; White,Crites, Taylor, & Corral, 2009), 也出现在内隐的语义启动和Stroop变型任务中(杨亚平, 王沛, 尹志慧, 陈庆伟, 冯夏影, 2015; Wang et al., 2011)。N400效应主要反映的是基于记忆经验的语义评估过程,其产生由多个语义加工环节构成(见图1)。因此上述结果证明外显和内隐刻板印象确实都依赖分布式的语义加工网络(Lau, Phillips, & Poeppel, 2008)。但另一方面, 内隐和外显刻板印象的语义表征机制又存在明显差异。实验发现, “分类−确认”任务或是“人物−地点”判断诱发的外显刻板印象, 主要与负责文字编码及外显语义知识表征的舌回(lingual gyrus)、梭状回及颞中回(middle temporal gyrus)存在明显联系(Contreras, Banaji, & Mitchell, 2012;Quadflieg et al., 2009)。但IAT和EAST所诱发的内隐刻板印象却主要激活了左侧额下回与外侧颞叶负责的内隐语义启动通道(Gozzi, Raymont, Solomon,Koenigs, & Grafman, 2009; Knutson, Mah, Manly, &Grafman, 2007; Wong, Harris, & Gallate, 2012; 贾磊, 2013)。有鉴于此, 单就记忆表征机制看, MMS模型能就内隐和外显刻板印象做出有效解释。

图1 基于元分析所得到的N400加工机制
1.2.3 多重记忆系统模型的局限与不足
然而, MMS模型只针对一般语义记忆(或概念知识)的表征机制, 但语义记忆层面的差异可能仅是外显和内隐刻板印象表征差异的一环, 实际还涉及社会特质与信念加工, 不同意识与注意水平的影响调制以及认知控制等问题。受此影响,MMS模型对外显和内隐刻板印象的理论解释也存在诸多不足:
第一, 存在对刻板印象社会性内容的解释缺陷。相比一般语义知识, 刻板印象的语义内容指向不同群体范畴的社会特质与行为特征。而MMS模型的现有框架无法区分刻板印象同一般知识在语义内容上的差异, 因此无力回答“刻板印象的群体特质如何表征”, “刻板印象为何带有牢固持久的信念与期望”等问题。而神经成像发现, 刻板印象区别于一般语义知识的本质特征在于其独特的社会认知评估机制——除有语义记忆脑区参与外, 还有内侧颞叶皮层(medial prefrontal cortex,MPFC)、扣带回、颞顶连接区(temporo-parietal junction, TPJ)等负责社会性信息加工脑区的参与。而这些脑区恰好是印象形成、特质表征和信念加工的关键区(Contreras et al., 2012; Quadflieg et al.,2009; van Overwalle, 2009)。以上证据暗示, 记忆表征差异只是外显和内隐刻板印象机制差异的一部分。而在记忆基础之外, 还存有认知编码与社会评估的机制差异。
第二, 存在对注意线索和任务意图作用机制的解释困难。MMS模型仅重视外显和内隐刻板印象表征的记忆基础差异。但依照内隐认知理论,决定刻板印象语义激活趋于内隐还是外显的关键因素是个体进行社会知觉时的注意线索、意识水平和任务意图(Blair, 2002; Greenwald & Banaji,1995)。根据迁移适当加工理论(Roediger, 1990),上述因素决定了个体的知觉加工是趋于概念驱动(外显)还是知觉驱动(内隐)。但MMS模型既不涉及一般性的注意编码问题, 也不涉及注意、意识或意图感知问题, 所以仅从它所强调的记忆表征基础出发, 无法解释注意、意识以及任务意图对外显和内隐刻板印象的影响作用机制。
实际研究表明, 刻板印象对社会信息的范畴分类加工可以先于 N400所代表的语义提取与评估, 发生在更早的注意或感知编码阶段:(1) ERP研究显示, 不管任务方式是面孔分类还是语义启动, 种族刻板印象激活导致的社会分类加工始终可以出现在 P2、N2代表的早期注意编码阶段(Correll, Urland, & Ito, 2006; Jia et al., 2012)。与此类似, 性别刻板印象也能在 N230成分代表的前语义(per-lexical)加工阶段对性别语义材料作出有效分类(贾磊, 张庆林, 李肖, 孙晓, 肖宵, 2014)。上述现象无法仅用记忆模型解释, 但却可以证明刻板印象激活确实存在概念驱动加工以外的知觉驱动加工。(2) fMRI研究也证明, 刻板印象激活确实会发生在较低层级的注意与知觉编码阶段。例如, 在采用图片分类任务的内隐刻板印象研究中,枕颞结合部负责注意识别的神经皮层, 如枕叶面孔识别区(occipital face area, OFA)、纹外区身体识别区(extrastriate body area, EBA)、梭状回的面孔识别区(fusiform face area, FFA)和身体识别区(fusiform body area, FBA), 均对图片中隐含的刻板违背信息极为敏感(Quadflieg et al., 2011)。而在外显任务诱发的刻板印象研究中, 负责注意线索监控调节的喙部前扣带回(rostral anterior cingulate cortex, rACC), 承担意图知觉的颞顶连接区, 以及代表意识和动机输入的脑岛(insula), 也对刻板信息敏感(Amodio, 2014; Contreras et al., 2012;Mitchell, Ames, Jenkins, & Banaji, 2009)。上述脑区都不是内隐和外显记忆的功能区, 而多是负责注意识别与社会评估的脑区。因此, 忽视任务意图和注意线索的调制作用而将刻板印象的表征和激活机制限定在语义记忆框架内, 显然低估了刻板印象语义表征机制的复杂性, 也无法合理解释内隐和外显刻板印象的机制差异。
第三, 对刻板印象语义激活过程中环境背景与认知控制的忽视。刻板印象的语义激活并不等于刻板知识信念的表达和运用。原因在于受个人意图和社会背景影响, 激活过程会受到中枢认知控制系统的监控。不管激活方式属外显还是内隐,其表达和运用过程始终如此。据此, 双加工理论主张将刻板印象激活解释为“A+C”两阶段加工(Devine, 1989), 其中A阶段代表刻板印象的自动激活, 而随后的 C阶段代表意图和背景卷入下个体选择运用、忽视, 还是控制刻板印象。作为对比, MMS模型仅涉及外显和内隐刻板印象的记忆表征问题, 并不涉及对认知控制机制的解释。然而, 表征机制的不同必然导致认知控制机制的差异。因此, 对认知控制机制的探讨可视作是表征机制问题的进一步延伸, 能够间接体现外显和内隐刻板印象的分布式语义表征机制。事实上近期确实有研究表明, 受语义表征机制差异的影响,中枢认知控制系统对外显和内隐刻板印象的认知控制机制的确存有差异。对于IAT和EAST诱发的内隐刻板印象, 对它的认知控制更为依赖左脑背外侧前额皮层与右脑前内侧额叶皮层构成的认知控制通道; 而对于“分类−确认”任务和“人物−地点”判断所诱发的外显刻板印象, 其认知控制过程则似乎更为依赖颞顶连接区负责社会语义冲突评估的下顶叶(inferior parietal lobe, IPL; 贾磊, 2013;Cattaneo, Mattavelli, Platania, & Papagno, 2011)。
1.3 刻板印象中的群体特质与社会信念评估:心理化系统与社会心理化加工的引入
社会知识区别于一般知识的实质在于其对象和内容带有的社会性。这种社会性难以通过传统行为手段予以过程展现, 但却可以通过社会认知神经科学所倡导的“社会脑”模型予以揭示。这一“社会脑”模型的独特之处在于, 它质疑一般记忆理论在社会认知下的推论, 强调社会知识表征机制与评估过程的特异性。由镜像神经元理论和心理理论升级而来的“镜像系统−心理化系统”模型即是该模型系列近年来的突出代表。
1.3.1 社会知识的表征框架:“镜像系统−心理化系统”社会脑模型
“镜像系统−心理化系统”模型由 van Overwalle等人(van Overwalle, 2009; van Overwalle & Baetens,2009)基于神经成像的元分析提出, 其观点实质是对镜像神经元系统(Mirror Neuron System, MNS)理论的深化。与后者不同的是, 它将单一的镜像神经元系统从功能上分解为镜像系统和心理化系统两个模块, 分别对应负责他人行为感知的社会知觉功能, 和负责特质、信念、自我参照等复杂加工的社会认知功能(见图2)。其中, 镜像系统因其分布的镜像神经元在行为感知与模仿中的镜像知觉功能得名, 主要参与对他人行为的模式识别、理解、模仿及对自我身体行为的执行监控。心理化系统则因心理理论(Theory of Mind, ToM)以及理解和推测他人心理状态的社会心理化(social mentalizing)加工而得名, 与社会认知的印象形成、特质表征、信念加工、自我参照以及社会道德存有紧密联系。相比前人理论, 该模型理论的独特之处在于它试图站在认知神经科学视角, 将镜像神经元理论代表的心理模拟论和传统心理理论统合在一个“社会脑”模型下。
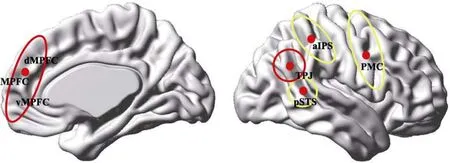
图2 心理化系统与镜像系统的脑区构成
1.3.2 群体特质与刻板信念:内侧额叶皮层与颞顶连接区的心理理论功能
据“镜像系统−心理化系统”模型, 刻板印象对社会他人的特质理解和信念认识体现的是心理理论能力, 属心理化系统的功能范畴。心理化系统由内侧额叶皮层、颞顶连接区及楔叶(cuneus)构成。其中, 内侧额叶皮层与颞顶连接区在印象形成、自我加工以及特质表征和信念加工中起关键作用, 二者被合称为核心心理化脑区。内侧额叶皮层被公认为社会认知的高级中枢, 它与顶叶和颞叶的功能皮层存在广泛神经连接, 据此可对顶颞部存储的内源记忆和感知获取的外源信息进行系统评估与整合。依照结构上的差异, 内侧额叶皮层可分为背内侧(dMPFC)和腹内侧(vMPFC)两部分。实验发现, 背内侧和腹内侧前额皮层均参与了特质和信念加工, 但彼此间存有功能差异。其中, 前者负责对陌生他人的特质和信念加工, 与印象形成及其改变密切相关(Gilron & Gutchess,2012), 而后者又被称为前喙部额叶皮层(arMPFC),对自我和熟悉他人敏感, 与特质和印象的熟悉性编码有关。
颞顶连接区则位于颞顶交汇处, 毗邻负责语义记忆的缘上回(supramarginal gyrus)及语义理解的角回(angular gyrus; 即Wernicke区)。据已有研究可将其认知功能归纳为三点:一是作为具身认知(embodiment cognition)中枢, 参与自我参照加工,并负责从知觉主体上区分自我与他人; 二是作为社会意图的初级评估中枢, 参与行为意图分析,并和内侧额叶皮层一同进行特质、信念、共情乃至道德等高级社会认知加工; 三是作为不随意(被动)注意的编码中枢, 参与刺激驱动的注意选择及内隐认知(van Overwalle, 2009; 贾磊等, 2015)。
内侧额叶皮层与颞顶连接区已被近期的 fMRI和TMS (Transcranial Magnetic Stimulation,经颅磁刺激)研究证明为刻板印象的关键功能区(Cattaneo et al., 2011; Knutson et al., 2007; Mitchell et al.,2009; Quadflieg et al., 2009)。这些研究发现, 内侧额叶皮层的心理化功能对外显和内隐刻板印象均至关重要。对临床病人的研究发现, 腹内侧额叶皮层受损后, 个体在 IAT中的内隐刻板印象表现会显著受影响(Gozzi et al., 2009; Milne & Grafman,2001)。作为对照, 杏仁核先天受损的威廉姆斯综合症(Williams syndrome)个体虽会失去情绪性的种族偏见能力, 但依旧保留有性别刻板印象能力(Santos, Meyer-Lindenberg, & Deruelle, 2010)。考虑到内侧额叶皮层对特质与信念加工的特殊意义,它很可能作为心理化加工中枢, 与负责行为意图初步分析的颞顶连接区, 以及负责刻板语义信息存储和提取的前部颞叶皮层(anterior temporal lobe,ATL), 协同参与了刻板印象的表征与激活过程。
1.4 内隐和外显刻板印象激活中的背景监视与意识控制
1.4.1 刻板印象的激活网络与控制网络
相较于 MMS模型, 双加工理论的优势在于能将刻板印象的认知控制过程同其自动激活过程加以区分。该理论虽然不适用于内隐和外显刻板印象在记忆表征机制上的差异解释, 但却可以解释和说明内隐和外显刻板印象激活涉及的背景监视与意识控制问题。有鉴于此, 近期仍有不少认知神经研究从双加工理论的视角出发, 探讨刻板印象的认知控制机制(Amodio, Devine, & Harmon-Jones, 2008; Cattaneo et al., 2011; Knutson et al.,2007; Wong et al., 2012)。这类研究多采用内隐测量任务, 获得的结论也大致相同, 均认为是由分布在外侧和背侧额叶的中枢控制系统承担刻板印象的认知控制加工。根据双加工理论“A+C”两阶段加工的观点, Amodio (2014)近期总结整理了上述研究的成果, 提出了刻板印象“表征网络+控制网络”的神经模型(见图3)。
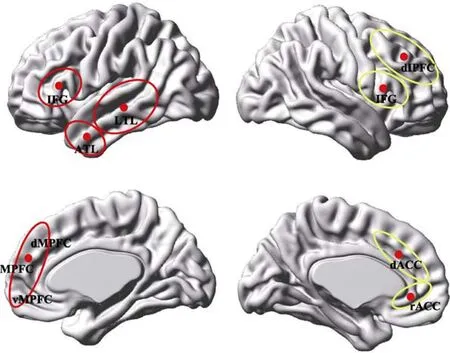
图3 刻板印象的语义表征脑区与认知控制脑区
在该模型中, 刻板印象表征网络主要由左侧额下回(IFG; 负责语义工作记忆和概念激活加工)、两半球的内侧额叶皮层(MPFC; 负责印象形成、自我参照及心理化加工)以及前部与外侧颞叶(ATL & LTL; 负责社会知识存储与提取)构成。控制网络则主要包括右侧额下回(IFG; 负责认知控制)、背侧前扣带回(dACC; 负责冲突监控)、背外侧额叶皮层(dlPFC; 承担行为选择功能)以及喙部前扣带回(rACC; 负责对外在线索进行注意监控)。
需要指出的是, 该模型仅限于刻板印象的认知解释, 其表征网络中部分脑区的特征定义与社会认知一般双加工理论的 X-C双系统模型(Lieberman,2007)有着一些重要差别。在一般双加工模型中, 腹内侧额叶皮层属于自动加工的X系统(取自automatic同义的 reflexive), 而背内侧额叶皮层则属于控制加工的C系统(取自controlled同义的reflective);此外, 位于外侧额叶的额下回也被纳入了具有控制加工特性的 C系统。相较于表征网络, 控制网络则完全符合X-C双系统模型下C系统控制加工的特征定义。
1.4.2 内隐与外显刻板印象激活中的意识控制与意图编码
依照 Amodio (2014)模型对于控制网络的机制解释, 喙部前扣带回能够基于它与额叶皮层间的复杂神经联络, 通过输入自我意识和信念意图,进而对社会准则以及环境背景等外在线索进行主动注意监测。背侧前扣带回则承担冲突监控职责,待注意系统监测有认知冲突后, 由背外侧额叶皮层进行认知和行为控制。上述解释契合了前扣带回与前额皮层不同脑区认知功能的研究发现, 除了能够解释自我意识、任务意图乃至信念期望输入所引发的自上而下的刻板印象控制过程外, 实际也能说明自我意识和信念意图以概念驱动形式激活外显刻板印象的过程。但问题在于, 相比先验性的有意状态, 究竟无意识或意图干预时刻板印象是如何被自动激活的, 目前还缺少相应的机制解释。由于有无意识或意图参与是调制刻板印象激活以形成内隐和外显刻板印象的关键, 因此解决了这一问题, 实际也就弄清了内隐和外显刻板印象机制差异的本质。
尽管尚未有认知神经研究探讨外显与内隐刻板印象中的意识和意图机制问题, 但相近和相关领域的新发现提供了一定启示。首先是目标推理和特质推理领域有关内隐和外显机制的比较研究(Ma et al., 2012; Ma, Vandekerckhove, van Overwalle,Seurinck, & Fias, 2010)。这些研究发现, 内隐性的自发特质(或目标)推理与外显性的有意特质(或目标)推理的认知加工均主要激活了颞顶连接区和内侧额叶皮层构成的核心心理化系统, 但二者的激活程度和先后顺序存在显著差别。其中, 自发特质推理更为偏好具有内隐和自动加工特性的颞顶连接区, 而有意特质(或目标)推理更偏好带有意识精细编码特征的内侧前额皮层。而且无论是在目标推理中, 还是在特质推理中, 具有自动化特性的颞顶连接区, 它的激活始终快于具有控制性精细编码特征的内侧前额皮层。以上发现说明, 心理化系统可以根据意识和意图编码水平的不同做出灵活的加工调整, 而负责被动注意和自动意图编码的颞顶连接区, 它很可能是内隐与外显机制分离的关键调节者。鉴于刻板印象是自发特质推理重要的内容来源, 因此内隐和外显刻板印象的机制分离很可能是心理化系统与记忆系统同时进行意识与意图不同水平编码的产物, 而主动和被动的注意选择刚好扮演了推动意识与意图编码的角色。其次是有关喙部前扣带回与脑岛意识功能的研究。近期研究发现, 分布在喙部前扣带回与脑岛的纺锤状神经元(又称 von Economo神经元)极可能与注意选择与意识水平的控制调节有关(Craig, 2009)。现有的刻板印象研究中也不乏这两个脑区的报告(e.g., Knutson et al., 2007; Mitchell et al.,2009), 然而却少有与此相关的讨论分析。这两个脑区都与内侧额叶有着广泛的联系, 因此就刻板印象激活过程而言, 对外源刺激线索的注意选择,以及内源性意识意图的输入和调节, 均可能与这两个脑区有关。对喙部前扣带回与脑岛的认知功能分析, 很可能就是解答外显和内隐机制差异的关键。
1.4.3 “激活网络+控制网络”模型的局限与不足
Amodio (2014)所提出“激活网络+控制网络”神经模型是对Devine (1989)双加工理论的深化发展。该模型继承了双加工理论有关刻板印象“自动激活+认知控制”的两阶段过程观, 并从脑科学视角归纳解释刻板印象的认知机制。它的优点在于能够较好解释内隐和外显刻板印象的认知控制过程。然而, 受限于双加工理论的固有框架, 这种二元划分的模型结构也存有明显的局限与不足。例如, 它无法区分外显和内隐刻板印象的机制差异,也无法体现社会心理化加工同语义记忆加工存在的本质功能差异。况且, 它对心理化系统的功能认识同样存在问题, 不仅忽视了颞顶连接区对于刻板印象的功能意义, 而且对内侧额叶皮层的认知特性分析也与当前社会认知神经科学的一般双加工理论存在观点冲突。
2 研究构想
综上所述, 以 MMS模型和双加工理论为代表的内隐社会认知理论, 其现有的理论框架在解释内隐和外显刻板印象的机制时已存有明显的局限和不足。针对以上的局限和不足, 本项目计划在借鉴当代认知神经科学研究成果与方法的基础上, 探索刻板印象在内隐和外显两种表现型上的机制差异, 进而构建理解刻板印象分布式语义表征机制的认知神经模型。
2.1 刻板印象语义分布式表征模型的研究构想
2.1.1 总体构想:从 MMS模型和“激活网络+控制网络”模型到三元模块的结构分析框架
在思考和设计新的刻板印象语义分布式表征模型时, 有必要再次分析既有MMS模型与“激活网络+控制网络”模型的优势与局限。MMS模型将刻板印象存在的语义记忆基础分为了外显语义记忆和内隐语义记忆, 这种二元划分侧重于解释刻板印象的记忆表征机制, 可以说明外显和内隐刻板印象在记忆层面的表征基础差异, 但不好解释刻板印象社会性的群体特质和信念特征, 也不好解释刻板印象激活过程中的认知控制机制。而“激活网络+控制网络”模型则是将刻板印象的认知神经基础分为了表征激活网络和控制加工网络, 这种二元划分更侧重解释刻板印象的认知控制机制,能够说明外显和内隐刻板印象激活过程中背景监视与意识控制成分, 但却不涉及记忆表征机制和社会心理化机制。
针对上述两个模型存在优势与局限, 一种合理的构想是将刻板印象的 MMS模型、“激活网络+控制网络”模型, 同已有的“社会脑”模型结合起来,全面分析外显和内隐刻板印象的加工机制差异。具体而言, 这一构想的实质是将刻板印象的表征机制进一步分解, 分离为 MMS模型强调的语义表征模块和“社会脑”模型强调的心理化模块, 加上双加工理论所强调的认知控制模块, 形成三元模块的结构分析框架。在这一框架中, 语义表征模块负责输入信息的语义编码, 以及在长时记忆系统下的存储和提取。根据MMS模型, 该模块应该同时具有内隐记忆和外显记忆两个记忆子系统, 分别表征内隐和外显的刻板语义内容。而根据内隐社会认知理论, 这一模块受意识和注意系统调制,进而决定信息编码、存储和提取方式是趋于知觉驱动(内隐), 还是概念驱动(外显)。社会心理化模块则主要由颞顶连接区和内侧额叶皮层构成, 负责对语义内容进行社会认知的心理化加工, 承担信念激活和特质内容评估等心理化加工任务。据现有证据推测, 此模块同样受意识状态和任务意图的影响, 它在外显和内隐任务下也可能存在加工差异。最后, 认知控制模块负责对社会背景环境的监测与分析, 同时对注意感知、认知评估及判断决策进行认知监控, 从而对前两个模块形成有效控制。
2.1.2 研究内容:问题的分解及其假设切入点
依据上述的三元模块构想, 分析和比较内隐和外显刻板印象的分布式表征机制, 可在认知研究的操作层面还原为检验和考察刻板语义信息在语义表征模块、社会心理化模块以及认知控制模块下的协作加工过程, 确认不同模块在外显和内隐刻板印象中相对的认知功能, 分析每个系统功能模块的认知特性, 并考察彼此间的交互影响。据此, 本项目拟将研究内容分解为三个主要问题,进行实验假设检验。这3个问题及其假设切入点具体如下:
问题一:内隐和外显刻板印象在表征机制上的异同。刻板印象既包括有关群体特质的语义知识, 也包括对于群体刻板化的内在信念。所以检验外显和内隐刻板印象的表征机制, 实际可以理解为检验二者在语义与信念上的表征机制差异。分布式并行加工网络模型认为, 同一过程阶段下的不同信息过程属并行加工, 并且这些加工又分别激活了大脑分布各异的神经网络。根据双加工理论有关刻板印象“自动激活+认知控制”两阶段过程观, 如果将刻板印象激活下的语义记忆加工和心理化加工视作同一自动化加工阶段下的并行过程, 那么检验外显和内隐刻板印象的表征机制的异同, 实质是检验这二者在语义表征模块和社会心理化模块上的加工机制差异。从脑认知角度看,这一机制差异不仅包括了外显和内隐刻板印象在语义表征模块和社会心理化模块上的脑区激活差异, 也包括了这两个功能模块脑区上的功能协作差异。所以本项目认为, 分析内隐和外显刻板印象表征机制的异同, 实际可围绕同一内容刻板印象的外显和内隐激活过程, 检验二者在语义记忆脑区和心理化系统脑区上的激活异同, 并考察上述脑区的功能连接(functional connectivity)和有效连接(effective connectivity)。
问题二:意识水平和任务意图影响内隐和外显刻板印象激活的神经机制。内隐社会认知理论将意识水平和任务意图视作是影响刻板印象激活趋于内隐还是外显的关键因素。当前虽有不少行为研究已经论证了这一点, 但依然缺少相应的脑科学证据。鉴于现有研究已将喙部前扣带回、脑岛视为意识影响主动注意的关键区, 而将颞顶连接区视为目的意图自动编码和被动注意的关键区,因此本项目认为, 可以检验上述三个脑区是否在内隐和外显刻板印象任务下存在显著激活差异,验证此差异是否能够预测语义表征脑区、社会心理化脑区在内隐和外显任务下的激活。如果答案是肯定的, 那么即证明它们确实是意识和意图影响刻板印象外显和内隐激活的关键脑区。
问题三:认知控制网络对于外显和内隐刻板印象的不同认知控制路径。本项目认为, 作为问题一的延伸, 若外显和内隐刻板印象存在语义表征机制和心理化评估机制上的不同, 那么在刻板印象冲突情境下, 中枢认知控制系统对于二者语义记忆提取和心理化评估的认知控制和干预, 理应也存在加工机制上的差异, 且此差异将通过ERP所反映的过程机制以及 fMRI所反映的脑区激活差异体现出来。
3 研究方案设计
根据研究的问题假设, 本项目设计了 4个研究展开实验验证。
3.1 研究一:基于“范畴−特质”语义的外显和内隐刻板印象机制比较
刻板印象的激活多建立在群体范畴与特质的语义关联基础上。根据这一特性, 前人研究中经常将群体的范畴类别作为先行的语义刺激, 激活刻板知识和信念, 并诱导其后的刻板运用和控制过程(Bodenhausen, Mussweiler, Gabriel, & Moreno,2001)。研究一计划采用“范畴−特质”的经典范式,检验内隐和外显刻板印象在语义表征、社会心理化以及认知控制机制上的差异。实验任务采用典型的“范畴−特质”刺激任务。根据“范畴−特质”的匹配/冲突与否, 分别设置刻板印象激活(语义匹配)与控制(语义冲突)两种。同时, 任务给范畴类别添加了额外线索(不同颜色), 进而操作范畴线索, 引导被试就同一特质完成外显的刻板语义匹配判断(判断范畴词与特质词是否匹配相符)或是内隐的语义启动任务(仅判断特质词效价正负),最终形成 2 (任务指向:外显/内隐) × 2 (“范畴−特质”刻板语义关联:匹配/冲突)的完全被试内设计。研究假设, 外显语义匹配判断任务下, 范畴与特质间的概念语义匹配与否将分别诱发外显刻板印象激活和控制过程。而在语义启动任务下, 虽然任务指向特质的情绪效价判断, 但基于“范畴−特质”间的语义关联, 范畴与特质间的语义匹配与否将分别诱发内隐刻板印象激活和控制过程。
研究一分两个实验开展研究。其中实验1将通过ERP技术比较内隐和外显刻板印象激活和控制机制。实验预期, 外显和内隐刻板印象在语义记忆表征、社会心理化加工以及认知控制上存有的机制差异, 将通过各自诱发的ERP成分(如反映特质心理化加工的 P300成分和反映语义评估的N400成分)差异体现出来, 并进一步体现在时间频率分析(Time-frequency analysis)和溯源性的低分辨率电磁断层成像分析(low resolution electromagnetic tomography analysis, LORETA)的结果上。
实验2则借助fMRI脑区定位精度高的优势,比较外显和内隐刻板印象的脑机制异同。实验预期, 外显和内隐刻板印象将会因彼此在语义记忆和社会心理化机制上的差异, 于刻板语义匹配条件下显现脑区激活的差异, 并且此差异将进一步体现在对它们的感兴趣区(Region of Interests, ROIs)分析和动态因果建模(Dynamic Causal Modeling,DCM)结果中。其次, 由于外显和内隐刻板印象的认知控制和心理化加工分别依赖于中枢认知控制系统和心理化系统, 因此二者在刻板语义匹配和冲突两种条件下仍可能激活相同的认知控制脑区和心理化脑区。但由于语义表征机制的差异, 这种共同脑区的激活程度极有可能存在差异; 而且以它们为种子区的心理生理交互(Psycho-physiological Interactions, PPI)分析, 也将会因外显和内隐激活任务的差异而获得不同的脑功能连接结果。最后,若外显和内隐任务间存在喙部前扣带回、脑岛或是颞顶连接区激活差异, 且以它们为种子区的ROIs和PPI分析能够预测语义记忆和社会心理化脑区在外显和内隐任务上的激活差异, 那么即可认为上述脑区确属意识水平和任务意图影响刻板印象外显和内隐激活的关键脑区。
3.2 研究二:基于印象形成的外显和内隐刻板印象机制比较
研究一采用的刻板印象激活任务侧重于考察记忆中固有的群体“范畴−特质”语义关联机制。但在很多社会情境中, 个体是通过“对象−身份(范畴)−特质”的印象形成途径来激活刻板印象的。对此, 研究二计划将外显和内隐刻板印象机制考察放在社会认知情境下, 从印象形成角度进一步检验外显和内隐刻板印象加工机制差异。此研究采用典型的特质判断任务, 任务分为两个阶段。阶段一为印象形成阶段。此阶段先呈现对象面孔并随后呈现与其对应的群体身份标签, 要求被试完全掌握对象面孔与其群体身份的配对联系。阶段二为词语判断阶段。此阶段任务流程为先呈现面孔, 后呈现与其身份描述分别匹配、无关或语义冲突的特质词, 并混合以假词。此阶段要求被试根据提示完成真词/假词判断(内隐)或是特质与对象身份是否匹配的判断(外显)。实验采用2 (任务指向:外显/内隐) × 3 (特质词刻板语义属性:匹配/冲突/无关)的被试内设计。研究假设, 由于对象身份的学习获得, 真假词判断任务下的语义匹配和冲突条件将分别诱发内隐刻板印象的激活和控制加工; 而在身份与特质匹配判断任务下, 它们将诱发外显的刻板印象激活和控制加工。
研究二同样分为两个实验, 其中实验3为ERP实验, 实验4为fMRI实验, 研究技术路径与研究一中的实验1和实验2相同。由于实验的因素设计相近, 研究二的结果预期也基本类似于研究一。但由于任务方案间差异, 二者间结果预期存在以下不同。首先, 任务中加入了印象形成加工, 而印象形成更需要心理化系统的参与(Mitchell, Macrae, &Banaji, 2005; van Overwalle, 2009)。因此, 实验4预期会在内隐和外显刻板印象激活条件下观察到心理化脑区和语义记忆脑区的更强激活, 二者的功能和有效连接也将更易表现在在 PPI和 DCM分析结果中。其次, 研究二设计中新加入了无关特质的基线条件, 这一基线条件的设置有利于将刻板印象的自动激活、认知控制与无关特质心理化加工加以比较, 分析刻板印象的认知独特性。最后, 由于不涉及特质的效价判断, 故该任务能够排除因特质效价判断而导致的无关情绪输入。
3.3 研究三:外显和内隐刻板印象的认知发展研究
刻板印象的认知发展研究是近年来该领域研究关注的一个新问题。国外研究发现, 6岁左右的儿童已经能够基于口头性的特质语义理解, 在口语调查和口语提示的IAT任务下展现外显和内隐的刻板印象(Cvencek, Meltzoff, & Greenwald, 2011)。这说明此年龄段的个体已具备了内隐和外显刻板印象的能力。另有研究显示, 7~8岁的儿童已经能够理解抽象特质, 并能较为稳定地做出外显的有意特质推理(Alvarez, Ruble, & Bolger, 2001)。但近期有国内研究表明, 正常 8岁儿童仍难以就行为描述进行特质概括, 而且也尚未能灵活有效地建立对象与特质间的自发联结。而上述能力通常被视为个体运用内隐刻板印象进行自发特质推理的前提条件。但到11岁后, 儿童不但具备了上述能力, 还能激活内隐刻板印象, 并且通过它来影响自发特质推理过程(闫秀梅, 2010)。这一证据暗示, 个体刻板印象的能力发展可能具备阶段性, 8~10岁儿童由于社会心理化能力和语言符号运用能力的限制, 其在掌握一般的刻板知识后, 仍不能立即高效地构建对象与特质间的自动联结并灵活地将其运用在内隐社会判断中。对此, 本项目有理由推测, 8~10岁儿童受有限自发特质联结能力的制约,他们在复杂特质联结情境(如研究二的任务情境)下的内隐刻板印象激活能力可能会受显著影响;而外显刻板印象无需建立构建对象与特质间的自动联结, 将不受此影响。
有鉴于此, 研究三的实验5拟以8、10、12岁三年龄段的小学儿童为研究对象, 采用研究二中实验3的任务设计, 形成2(任务指向:外显/内隐) ×3 (特质词刻板语义属性:匹配/冲突/无关) × 4(年龄组:8、10、12岁儿童, 加上实验3获得的大学生的数据)的混合实验设计, 探讨外显和内隐刻板印象与年龄发展间的关系。由于前人研究证明,外显和内隐刻板印象的语义表征均与 N400有着紧密关联(王沛等, 2010; 杨亚平等, 2015; Wang et al., 2011; White et al., 2009)。而8~10岁儿童的书面言语机制有别于成人, 且这一差异会反映在 N400的波幅和地形分布上(Atchley et al., 2006)。因此实验 5计划以反映刻板特质语义评估的 N400为神经指标, 从年龄发展层面检验外显和内隐刻板印象各自的认知发展机制。根据闫秀梅(2010)的研究结论, 实验 5预测, 由于社会心理化水平和语言符号掌握能力的年龄差异, 不同年龄段个体在外显和内隐任务下的行为表现和N400将存在相互间的显著差异。其中, 如果 8岁儿童因社会心理化能力和语言能力的限制, 无法灵活有效地建立对象与特质间的自发联结, 那么相比其他年龄段个体, 他们在内隐的真假词判断任务中将不会因特质词刻板语义属性的变化而出现内隐刻板印象激活与控制的加工差异; 但其外显概念化的刻板印象判断将不受影响, 依然随特质词刻板语义属性的变化而出现外显刻板印象激活与控制加工。
3.4 研究四:资源有限条件下的内隐和外显刻板印象的激活与控制加工
研究四计划在研究一和研究二的基础上, 进一步探索外显和内隐刻板印象认知控制与认知资源关系问题。现有研究认为, 中枢认知资源(尤其是工作记忆资源)是否充裕, 是认知控制系统能否进行有效认知控制和干预的关键。双加工理论认为, 当刻板印象激活并与给定信息产生认知冲突时, 中枢认知控制系统将会启动, 通过消耗认知资源来控制刻板印象。但双加工理论没有考虑的是, 语义记忆的有意提取以及心理化系统对特质的精细评估, 也须消耗一定的认知资源。所以当认知资源受限时, 不仅刻板印象的认知控制加工会受影响, 其有意记忆提取和心理化加工过程也应受影响。据此推测, 高认知负荷对外显和内隐刻板印象激活及控制的影响很可能是不一致的。
因此, 研究四计划采用 ERP技术, 检验高认知负荷任务下, 工作记忆资源受限是否会对内隐和外显刻板印象激活及控制过程产生不同影响。此研究分为两个实验(即实验6和实验7), 在前两项研究中的实验 1和实验 3设计基础, 加入了认知负荷这一新自变量。该变量的操作采用工作记忆资源高度消耗的Flanker任务, 通过是否在特质判断任务前加入Flanker任务, 进而形成高认知负荷和低认知负荷两种条件。为了平衡新加入这一变量, 研究四的两个实验改用被试内 Block平衡设计, 平衡内隐和外显Block的任务顺序。由此, 实验6形成2 (认知资源:高认知负荷/低认知负荷) ×2 (任务指向:外显/内隐) × 2 (特质词刻板语义属性:匹配/冲突)的完全被试内设计。实验7则形成2(认知资源:高认知负荷/低认知负荷) × 2 (任务指向:外显/内隐) × 3 (特质词刻板语义属性:匹配/冲突/无关)的完全被试内设计。研究预期, 认知负荷的改变将通过工作记忆资源分别影响外显和内隐刻板印象激活和控制加工。但由于外显和内隐刻板印象的语义记忆表征模块、社会心理化模块以及认知控制模块存在的机制差异, 认知负荷的改变对外显和内隐刻板印象激活和控制加工的作用影响将会是显著不同的。这种不同将以实验三因素的三阶段交互作用形式体现在实验 6和实验 7的行为指标以及反映刻板特质语义评估加工的N400成分上。

图4 项目预期获得的理论模型框架示意
4 预期与展望
从理论建构角度看, 本研究提出的“语义表征−社会心理化−认知控制”三元模块构想可视作是对现有MMS模型、双加工理论(包括“激活网络+控制网络”模型)以及“镜像系统−心理化系统”社会脑模型的理论整合。这一整合能够克服原有理论模型在解释外显和内隐刻板印象机制问题时遭遇的多个困难, 进而合理地解释二者的分布式表征机制。
根据上述构想, 我们有针对性地提出了实验验证方案, 旨在进一步探讨和解释外显和内隐刻板印象表征机制异同以及二者激活过程的差异。如果研究的脑成像结果确如我们所假设的那样, 结合已有证据, 即可构建起外显和内隐刻板印象表征的宏观认知模型, 具体如图 4所示。而脑电研究结果将进一步补充该模型所未能标注的加工时间顺序信息。我们相信, 本项目的开展将从“社会脑”视角重新构建对于刻板印象的认知解释, 促进对刻板印象分布式表征和加工灵活性的科学理解,进而推动内隐社会认知领域的理论研究发展。
贾磊. (2013).语义刻板印象认知神经机制(博士学位论文).西南大学.
贾磊, 张常洁, 张庆林. (2015). 自发特质推理的神经基础——基于“镜像系统-心理化系统”模型的解析.心理学探新, 35(5), 393–399.
贾磊, 张庆林, 李肖, 孙晓, 肖宵. (2014). 语义性别刻板印象对注意返回抑制的影响——来自行为和 ERP 的证据.心理学探新, 34(5), 410–415.
王沛, 杨亚平, 赵仑. (2010). 刻板印象的激活效应: 行为和ERPs证据.心理学报, 42(5), 607–617.
闫秀梅. (2010).自发特质联结的年龄特点、加工机制及其对外显特质判断的影响(博士学位论文). 山东师范大学.
杨亚平, 王沛, 尹志慧, 陈庆伟, 冯夏影. (2015). 刻板印象激活的无意图性及其大脑神经活动特征.心理学报,47(4), 488–502.
Allport, D. (1985). Distributed memory, modular subsystems and dysphasia. In S. K. Newman & R. Epstein (Eds.),Current perspectives in dysphasia(pp. 32–60). Edinburgh:Churchill Livingstone.
Alvarez, J. M., Ruble, D. N., & Bolger, N. (2001). Trait understanding or evaluative reasoning? An analysis of children's behavioral predictions.Child Development, 72,1409−1425.
Amodio, D. M. (2014). The neuroscience of prejudice and stereotyping.Nature Reviews Neuroscience, 15, 670–682.
Amodio, D. M., Devine, P. G., & Harmon-Jones, E. (2008).Individual differences in the regulation of intergroup bias:The role of conflict monitoring and neural signals for control.Journal of Personality and Social Psychology, 94(1),60–74.
Amodio, D. M., & Ratner, K. G. (2011). A memory systems model of implicit social cognition.Current Directions in Psychological Science, 20(3), 143–148.
Atchley, R. A., Rice, M. L., Betz, S. K., Kwasny, K. M.,Sereno, J. A., & Jongman, A. (2006). A comparison of semantic and syntactic event related potentials generated by children and adults.Brain and Language, 99(3), 236–246.
Bargh, J. A., Schwader, K. L., Hailey, S. E., Dyer, R. L., &Boothby, E. J. (2012). Automaticity in social-cognitive processes.Trends in Cognitive Sciences, 16(12), 593–605.
Binder, J. R., & Desai, R. H. (2011). The neurobiology of semantic memory.Trends in Cognitive Sciences, 15(11),527–536.
Blair, I. V. (2002). The malleability of automatic stereotypes and prejudice.Personality and Social Psychology Review,6(3), 242–261.
Bodenhausen, G. V., Mussweiler, T., Gabriel, S., & Moreno,K. N. (2001). Affective influences on stereotyping and intergroup relations. In J. P. Forgas (Ed.),Handbook of affect and social cognition(pp. 319–343). Mahwah, NJ,US: Lawrence Erlbaum Associates Publishers.
Cattaneo, Z., Mattavelli, G., Platania, E., & Papagno, C.(2011). The role of the prefrontal cortex in controlling gender-stereotypical associations: A TMS investigation.NeuroImage, 56(3), 1839–1846.
Contreras, J. M., Banaji, M. R., & Mitchell, J. P. (2012).Dissociable neural correlates of stereotypes and other forms of semantic knowledge.Social Cognitive and Affective Neuroscience, 7(7), 764–770.
Correll, J., Urland, G. R., & Ito, T. A. (2006). Event-related potentials and the decision to shoot: The role of threat perception and cognitive control.Journal of Experimental Social Psychology, 42(1), 120–128.
Craig, A. D., (2009). How do you feel—now? The anterior insula and human awareness.Nature Reviews Neuroscience,10(1), 59–70.
Cvencek, D., Meltzoff, A. N., & Greenwald, A. G. (2011).Math–gender stereotypes in elementary school children.Child Development, 82(3), 766–779.
Devine, P. G. (1989). Stereotypes and prejudice: Their automatic and controlled components.Journal of Personality and Social Psychology, 56(1), 5–18.
Gilron, R., & Gutchess, A. H. (2012). Remembering first impressions: Effects of intentionality and diagnosticity on subsequent memory.Cognitive, Affective, & Behavioral Neuroscience, 12(1), 85-98.
Golby, A., Silverberg, G., Race, E., Gabrieli, S., O'Shea, J.,Knierim, K.,... Gabrieli, J. (2005). Memory encoding in Alzheimer's disease: An fMRI study of explicit and implicit memory.Brain, 128(4), 773–787.
Gozzi, M., Raymont, V., Solomon, J., Koenigs, M., &Grafman, J. (2009). Dissociable effects of prefrontal and anterior temporal cortical lesions on stereotypical gender attitudes.Neuropsychologia, 47(10), 2125–2132.
Greenwald, A. G., & Banaji, M. R. (1995). Implicit social cognition: Attitudes, self-esteem, and stereotypes.Psychological Review, 102(1), 4–27.
Hamilton, D. L., & Trolier, T. K. (1986). Stereotypes and stereotyping: An overview of the cognitive approach. In J. F.Dovidio & S. L. Gaertner (Eds.),Prejudice, discrimination,and racism(pp. 127–163). San Diego, CA, US: Academic Press.
Hilton, J. L., & von Hippel, W. (1996). Stereotypes.Annual Review of Psychology, 47, 237–271.
Jia, L., Dickter, C. L., Luo, J. L., Xiao, X., Yang, Q., Lei,M., … Zhang, Q. L. (2012). Different brain mechanisms between stereotype activation and application: Evidence from an ERP study.International Journal of Psychology,47(1), 58–66.
Knutson, K. M., Mah, L., Manly, C. F., & Grafman, J. (2007).Neural correlates of automatic beliefs about gender and race.Human Brain Mapping, 28(10), 915–930.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network for semantics: [De]constructing the N400.Nature Reviews Neuroscience, 9(12), 920–933.
Lieberman, M. D. (2007). Social cognitive neuroscience: A review of core processes.Annual Review of Psychology,58, 259–289.
Ma, N., Vandekerckhove, M., Baetens, K., van Overwalle, F.,Seurinck, R., & Fias, W. (2012). Inconsistencies in spontaneous and intentional trait inferences.Social Cognitive and Affective Neuroscience, 7(8), 937–950.
Ma, N., Vandekerckhove, M., van Overwalle, F., Seurinck,R., & Fias, W. (2011). Spontaneous and intentional trait inferences recruit a common mentalizing network to a different degree: Spontaneous inferences activate only its core areas.Social Neuroscience, 6(2), 123–138.
Milne, E., & Grafman, J. (2001). Ventromedial prefrontal cortex lesions in humans eliminate implicit gender stereotyping.The Journal of Neuroscience, 21(12), RC150.
Mitchell, J. P., Ames, D. L., Jenkins, A. C., & Banaji, M. R.(2009). Neural correlates of stereotype application.Journal of Cognitive Neuroscience, 21(3), 594–604.
Mitchell, J. P., Macrae, C. N., & Banaji, M. R. (2005).Forming impressions of people versus inanimate objects:Social-cognitive processing in the medial prefrontal cortex.NeuroImage, 26(1), 251–257.
Miyashita, Y. (2004). Cognitive memory: Cellular and network machineries and their top-down control.Science,306(5695), 435–440.
Payne, B. K. (2001). Prejudice and perception: The role of automatic and controlled processes in misperceiving a weapon.Journal of Personality and Social Psychology, 81(2),181–192.
Payne, B. K., & Gawronski, B. (2010). A history of implicit social cognition: Where is it coming from? Where is it now? Where is it going? In B. K. Payne & B. Gawronski(Eds.),Handbook of implicit social cognition: Measurement,theory, and applications(pp. 1–15). New York, NY, US:Guilford Press.
Quadflieg, S., Flannigan, N., Waiter, G. D., Rossion, B., Wig,G. S., Turk, D. J., & Macrae, C. N. (2011). Stereotype-based modulation of person perception.NeuroImage, 57(2), 549–557.
Quadflieg, S., & Macrae, C. N. (2011). Stereotypes and stereotyping: What's the brain got to do with it?European Review of Social Psychology, 22(1), 215–273.
Quadflieg, S., Turk, D. J., Waiter, G. D., Mitchell, J. P.,Jenkins, A. C., & Macrae, C. N. (2009). Exploring the neural correlates of social stereotyping.Journal of Cognitive Neuroscience, 21(8), 1560–1570.
Rissman, J., Eliassen, J. C., & Blumstein, S. E. (2003). An event-related fMRI investigation of implicit semantic priming.Journal of Cognitive Neuroscience, 15(8), 1160– 1175.
Roediger, H. L. (1990). Implicit memory: Retention without remembering.American Psychologist, 45(9), 1043–1056.
Santos, A., Meyer-Lindenberg, A., & Deruelle, C. (2010).Absence of racial, but not gender, stereotyping in Williams syndrome children.Current Biology, 20(7), R307–R308.
Squire, L. R. (2004). Memory systems of the brain: A brief history and current perspective.Neurobiology of Learning and Memory, 82(3), 171–177.
Squire, L. R., & Zola, S. M. (1996). Structure and function of declarative and nondeclarative memory systems.Proceedings of the National Academy of Sciencesof the United States of America, 93(24), 13515–13522.
Tulving, E. (1985). How many memory systems are there?American Psychologist, 40(4), 385-398.
van Overwalle, F. (2009). Social cognition and the brain: A meta-analysis.Human Brain Mapping, 30(3), 829–858.
van Overwalle, F., & Baetens, K. (2009). Understanding others' actions and goals by mirror and mentalizing systems:A meta-analysis.NeuroImage, 48(3), 564–584.
Wang, L., Ma, Q. G., Song, Z. F., Shi, Y. S., Wang, Y., &Pfotenhauer, L. (2011). N400 and the activation of prejudice against rural migrant workers in China.Brain Research,1375, 103–110.
White, K. R., Crites, S. L., Taylor, J. H., & Corral, G. (2009).Wait, what? Assessing stereotype incongruities using the N400 ERP component.Social Cognitive and AffectiveNeuroscience, 4(2), 191–198.
Wong, C. L., Harris, J. A., & Gallate, J. E. (2012). Evidence for a social function of the anterior temporal lobes:Low-frequency rTMS reduces implicit gender stereotypes.Social Neuroscience, 7(1), 90–104.
