青南藏区多房棘球绦虫系统发育学分析※
2016-01-22李超群张耀刚曹得萍
李超群,张耀刚,曹得萍
(青海大学医学院)
青南藏区多房棘球绦虫系统发育学分析※
李超群,张耀刚,曹得萍※※
(青海大学医学院)
摘要目的对青南藏区流行的多房棘球绦虫基因多态性及系统发育进行初步分析。方法利用线粒体DNA细胞色素氧化酶亚单位Ⅰ(COX Ⅰ)行分子标记,以COX Ⅰ为基因marker设计引物扩增COX Ⅰ部分基因序列,将得到的11条碱基序列通过Clustal X软件进行多重序列分析比对,与13条来自GenBank的棘球绦虫COX Ⅰ序列共同构建贝叶斯进化树。结果用于分析的标本序列与来源于亚洲的中国(四川,内蒙古)、哈萨克斯坦和欧洲的法国、斯洛伐克的多房棘球绦虫型聚于一枝,位于发育树的冠部,并在这一枝内部形成梳齿状分支。结论流行于青南藏区的多房棘球蚴标本存在基因多态性,其系统发育复杂。
关键词青南藏区多房棘球绦虫细胞色素氧化酶亚单位Ⅰ系统发育
PRELIMINARY PHYLOGENETIC ANALYSIS OF
EchinococcusmultilocularIN QINGHAI SOUTHERN
PLATEAU INFERRED BY CYTOCHROME OXIDASE
SUBUNIT I GENE SEQUENCE※
Li Chaoqun,Zhang Yaogang,Wu Defang,Cao Deping※※
(Department of Joint,Qinghai University Affiliated Hospital,Xining 810001,China)
Abstract ObjectiveTo analysis the phylogeny and genetic polymorphism of Echinococcus multilocular in Qinghai southern plateau.Methods We designed and synthesized the primers of COX Ⅰ gene,and used PCR to cloned 11 sequences from hepatic sample which infected byEchinococcusmultilocular.The sequences were aligned using Clustal X,and then,Bayesian analyses were performed with 13 sequences of COX Ⅰ gene from the GenBank.Results In the phylogenetic tree,all ofEchinococcusmultilocularisolates belongs toEchinococcusmultilocularwhich originate from different places,such as China(Sichuan,Inner Mongolia ),Kazakhstan of Asia,France and Slovakia of Europe,they located on the phylogenetic tree crown,and form a comb shaped branch in this branch of internal.Conclusion There are many gene polymorphism concerningEchinococcusmultilocularisspecimens which specially popular in the southern regions of Qinghai province.
KeywordsThe south region of Qinghai provinceEchinococcusmultilocularisCytochrome oxidase subunitⅠPhylogeny
泡球蚴病(Alveolar Echinococcosis,AE)是多房棘球绦虫(Echinococcus multilocularis,Em)的继绦期幼虫寄生于人体内(多见于肝脏部位)引起的疾患。在利用DNA序列特征对虫株确定研究中,因线粒体DNA具有其母系遗传、突变率高、极少发生重组等特性,所以在分子遗传学和人类学研究上都有重要的意义[1]。本文利用COXⅠ分子标记,通过对收集的11份流行于青海省多房棘球蚴标本和GenBank下载的13条绦虫COXⅠ基因序列行比对、碱基分析,构建系统发育树,进行系统发育学研究。
1材料与方法
1.1标本采集
标本采集于青海大学附属医院和解放军第四陆军医院,共11例,类型为AE手术标本。
1.2 基因组DNA提取
使用细胞/组织提取基因组试剂盒[离心柱型,天根生化科技(北京)有限公司],从标本中提取多房棘球蚴基因组DNA。
1.3PCR扩增、纯化以及测序
引物以细胞色素氧化酶 I(COXⅠ)基因为依据设计,序列[2]为F:5′-TATTGGAGATTATGGATCTG-3′和R:5′-AAAGCGGGCGCGCGGTGCTG-3′。反应体系:50 μL 的2×PCR Master缓冲液,10 pmol/μL的COXⅠ F和COXⅠ R各4 μL,所提取的基因组DNA 6 μL,用ddH2O补足100 μL。置于PCR扩增仪中进行聚合酶链式反应。扩增条件:在温度为94 ℃时行预变性,时长为8 min,同等温度下变性30 Sec,于54 ℃复性30 Sec,72 ℃下延伸1 min,变性、复性和延伸三个过程经历35 个循环后,72 ℃延伸8 min。反应结束后,用琼脂糖凝胶DNA回收试剂盒回收扩增产物。完成纯化后每个样本取5 μL产物在1%琼脂糖凝胶上进行电泳(观察目的片段),将剩余的回收产物送上海生工公司进行DNA单向测序。
1.4系统发育学分析
将扩增得到的COXⅠ基因序列在Genbank上进行Blast比对,选取与标本同源性高,且具有代表性的13条序列下载(表1),将这13条序列与测序所得的11条基因序列,在默认参数下,用Clustal X[3]软件进行比对,后续使用Seqvis v.1.3[4]、PAUP*4.0B10[5]和MEGA v.4.1[6]三个软件对24条序列的异质性、碱基组成和变异位点、遗传距离以及转化和颠换比率进行检验。使用软件DAMBE[7]对COXⅠ序列进行单倍型分析。最后建贝叶斯树:用Modeltest3.7[8]软件依据Akaike信息量准则(AIC)和PAUP*4.0B10软件运行得到最适于贝叶斯算法分析的核酸进化模型之后,用Mrbayes-3.1.2[9]软件构建最后的Bayesian系统发育进化树。
表113株棘球绦虫信息表
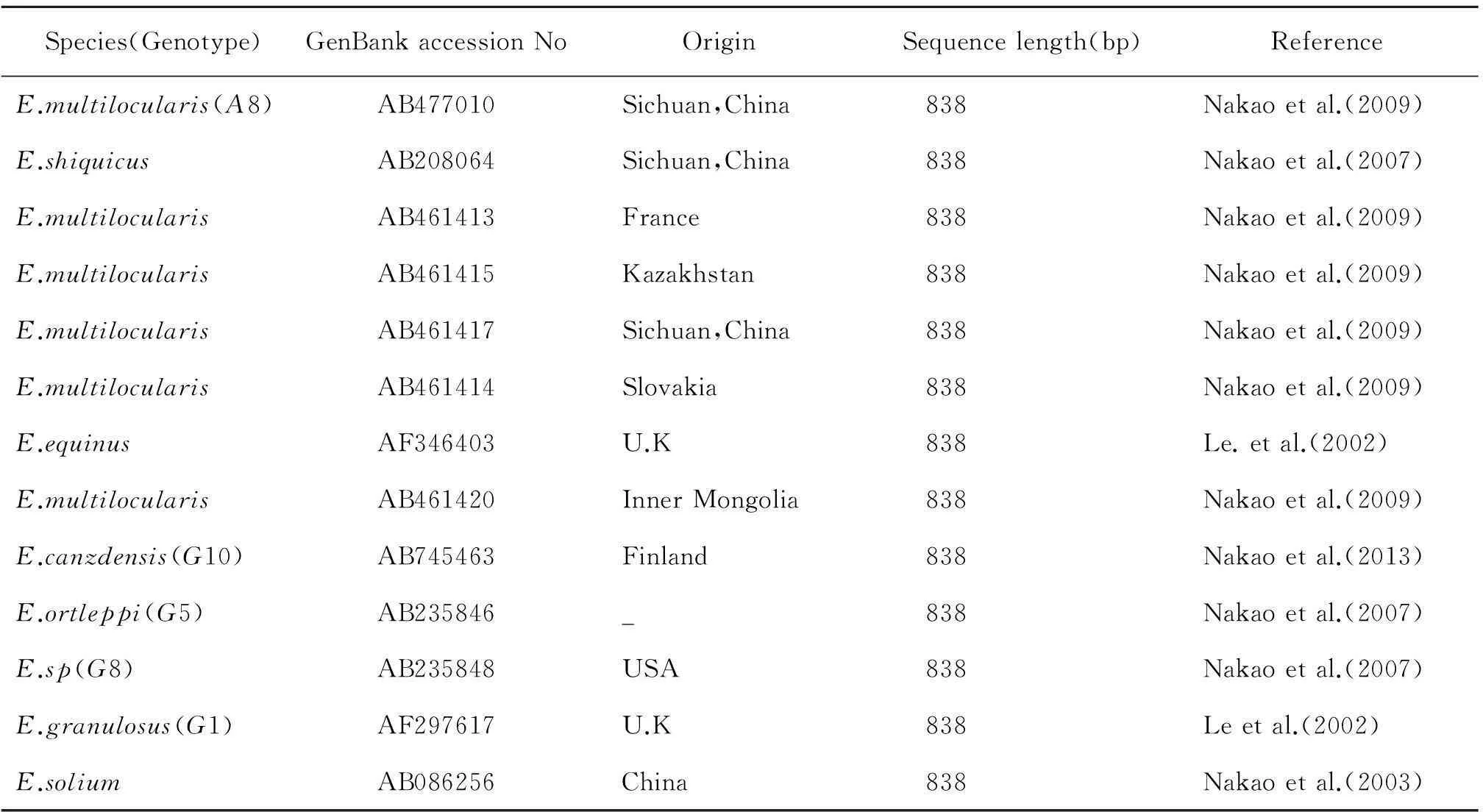
Table 1 Information of 13 Echinococcus spp
1.5统计学处理
使用Win-paup*4.0B10软件对序列碱基的组成进行分析,采用χ2检验对序列所有位点碱基组成的稳定情况进行统计分析,以P<0.05为差异具有统计学意义。
2结果
2.1PCR扩增结果
PCR产物在凝胶电泳检测中得到11例样本PCR扩增片段的位置相同(图1)。

图1 泡球蚴标本COXⅠ基因PCR扩增产物电泳检测图
2.2碱基组成和饱和检验分析
测序得到的11条序列在GeneBank进行BLAST比对,结果显示其与多房棘球绦虫基因的同源性高。利用SeqVis程序对24条序列(11条为样本序列,13条为GenBank下载序列)行异质性分析,显示异质性好(图2)。且经过DAMBE软件的单倍型分析,从24条序列中找出23个单倍型。用于分析的序列长度在778 bp(ZB40)到787 bp(ZB47)之间。在792个位点中共有276个变异位点、137个简约信息位点。4种碱基频率(表2)是A=0.172、C=0.100、G=0.235、T=0.493(碱基总和为1),A+T(0.665)>C+G(0.335)。样本的全部编码位点的碱基频率检测结果(χ2= 26.275df=69P=0.999)具有同质性。经MEGA v.4.1软件分析得出,用于分析的全部序列的转换与颠换比的平均似然估计值(R值)大小为1.611,全部序列平均遗传距离是0.073,两两遗传距离研究:样本虫株(ZB43)与猪带绦虫(AB086256)之间遗传距离最大为0.265。
图2 SeqVis 程序得到的24条序列异质性图
表224条序列碱基组成情况
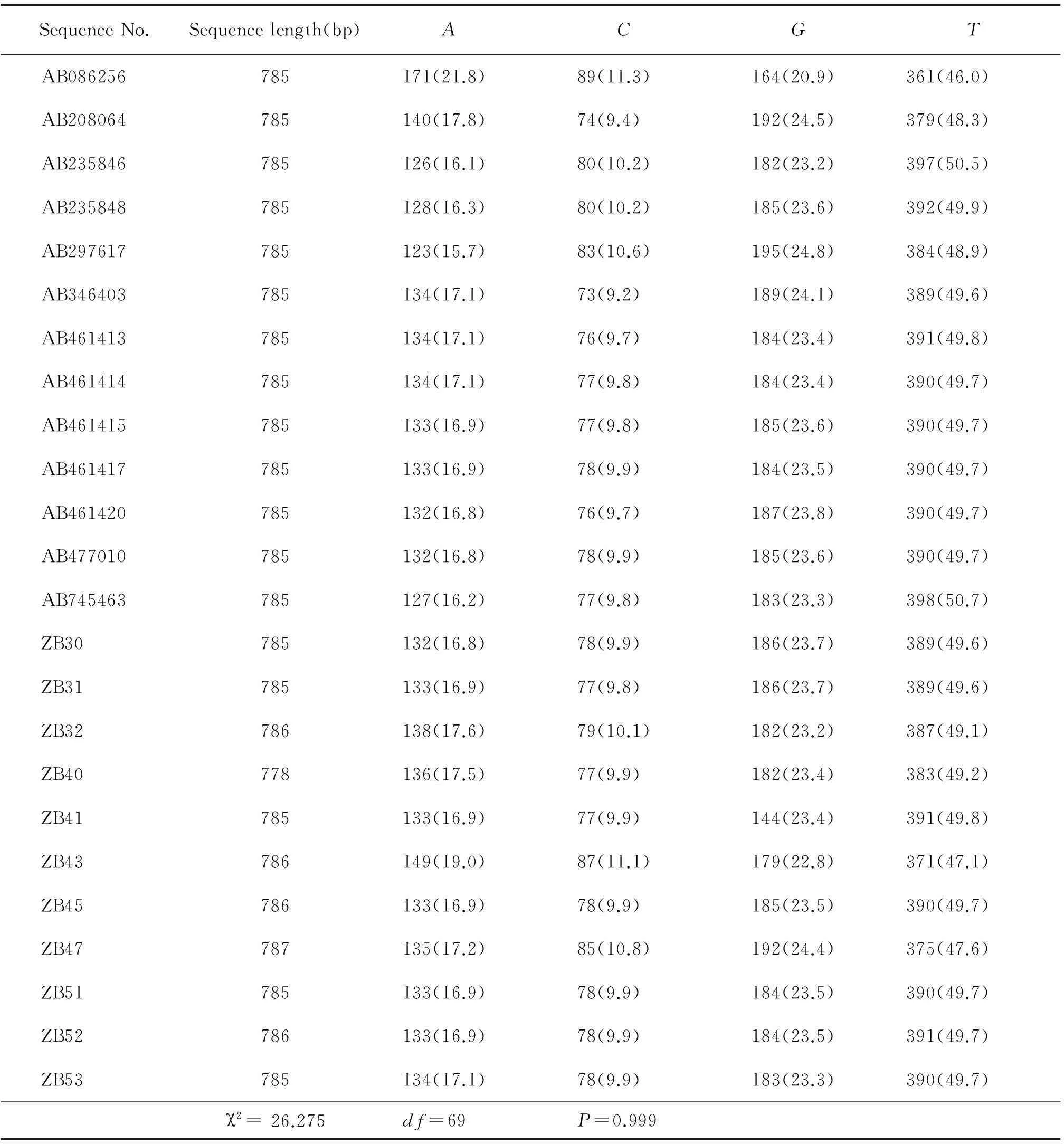
Table 2 the base composition rates in 24 sequence
2.3系统发育分析
把从GenBank下载所得的猪带绦虫(AB086256)的COXⅠ基因序列作为外群,构建Bayesian系统发育树(图3)。系统发育显示:青南地区的11例多房棘球绦虫标本均与来源为亚洲中国(四川,内蒙古)、哈萨克斯坦和欧洲的法国、斯洛伐克的多房棘球绦虫型,即均与AB477010(A8)、AB461417、AB461415、AB461420、AB461413和AB461414聚在一支[后验概率(PP)=1.00],位于发育树的冠部,但因存在较大的基因多态性,故在这一枝内部形成梳齿状分支。且虫株之间形成姐妹枝(PP>0.60)。AB745463(G10)和AB235848(G8)聚在一枝与AB235846(G5)形成姐妹枝。而所收集的青南地区多房棘球绦虫虫株标本与E.shiqucius(AB208064)虫株之间亲缘性较远。
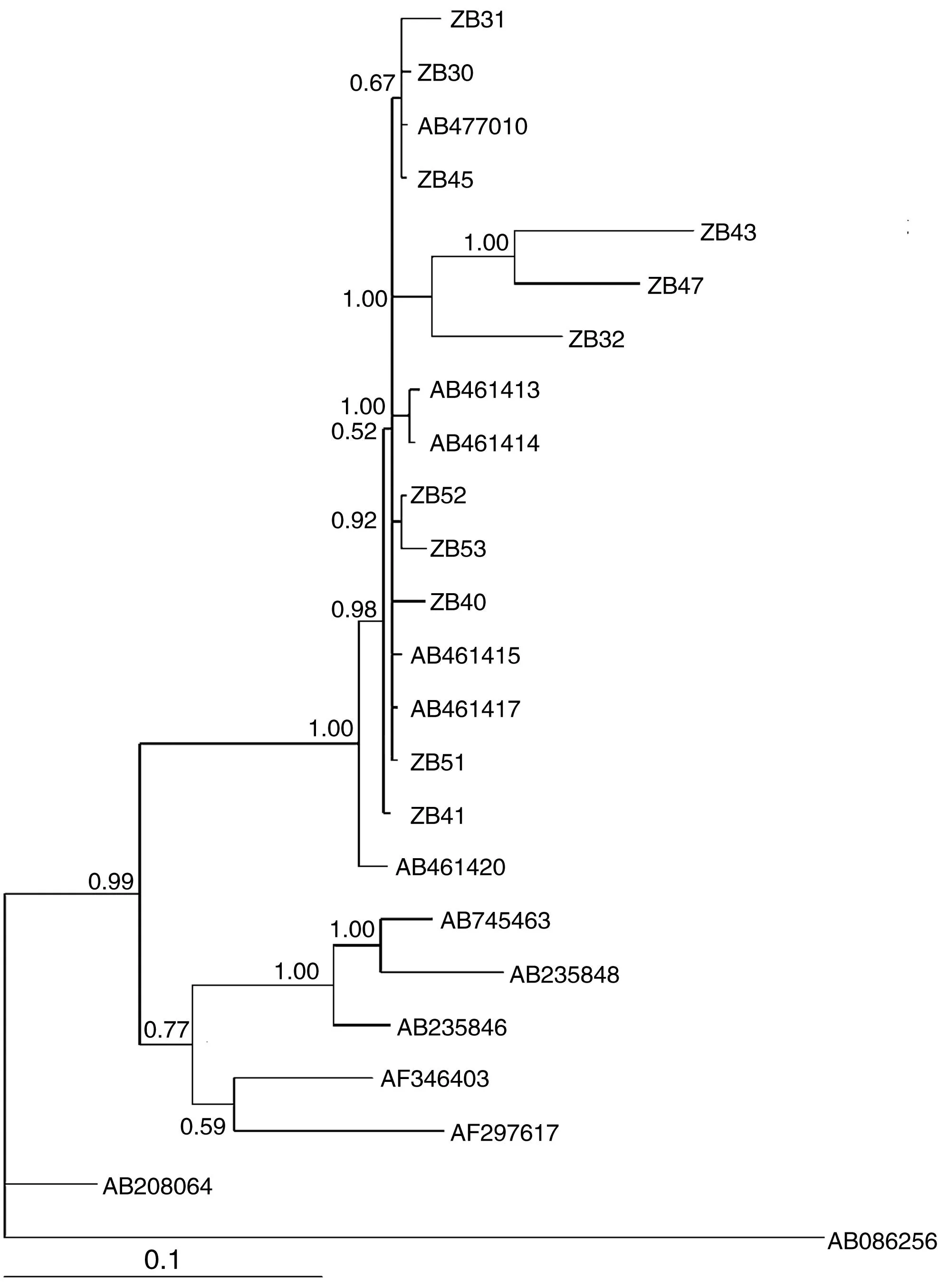
ZB:Sample NumberAB、AF:GenBank accession Number
3讨论
青海位于中国的西北部,因其具有的特殊高原气候环境,代表性的少数民族的生活方式和生活习惯,使当地人和动物棘球绦虫的感染十分普遍,囊型包虫和泡型包虫在该地区广泛流行和扩散[10],人体包虫病在六个牧业州均有较重流行,尤其青南各县,如称多县、甘德县、玛沁县、河南县和泽库县其平均患病率高达6. 81%[11]。故对青南地区棘球绦虫的研究有着重要的意义。付永[13]、朵红[14]等对青南地区狐、犬分离株进行同源虫株的研究中显示,青海省藏犬、野狐中存在有E.G1、Em、Echinococcusshiguicus(E.S)三种绦虫的感染,其中以细粒棘球绦虫(G1)型流行较广,但在野生狐狸中常见的绦虫感染类型是多房棘球绦虫的感染,也有部分病例是石渠棘球绦虫感染。同时由朵红等人在应用COXⅠ基因对所收集的棘球绦虫基因多态性进行分析时,得出32条基因序列,证实了棘球绦虫存在核苷酸变异以及具有基因的多态性。本研究以人体为分离株,利用COXⅠ对多房棘球绦虫的发育学进行研究。多房棘球绦虫主要分布于北半球的高寒地带,遍及北美、欧洲、亚洲这三洲。本研究结果显示,所收集标本与来源为亚洲的中国(四川,内蒙古)、哈萨克斯坦和欧洲的法国、斯洛伐克地方的多房棘球绦虫型聚于一支。说明青南地区的多房棘球绦虫与这些地方的多房棘球绦虫虫株间具有很高的同源性。在这枝内部,青南地区样本虫株ZB30、ZB31、ZB45和中国四川地方的AB477010(A8)虫株间形成姐妹枝(PP=0.67),证明这三个标本虫株与四川的多房棘球绦虫的(A8)型的亲缘性相近。此外,青南地区样本虫株ZB43和ZB47聚于一枝(PP=1.00)与样本ZB32形成姐妹枝(PP=1.00),可看出这3例虫株标本之间具有良好的亲缘性。而其他虫株也形成有不同的小分枝或成梳齿状,这些小分枝的形成证实了虫株间存在有基因的多态性。这种多态性的形成可能是由于青南地区独特的地理环境和气候条件,造成绦虫形成地理隔离和生殖隔离,从而导致多房棘球绦虫核苷酸变异形成。而在发育树下半部分可以看到除猪带绦虫外从Genbank上下载的虫株E.canzdensis(G10)- AB745463、E.ortleppi(G5)- AB235846、E.sp(G8)- AB235848、E.granulosus(G1)- AF297617和E.equinus-AF346403聚于一大枝,同时也根据亲缘性远近形成姐妹枝,但都与我们的标本虫株间具有相对较远的亲缘性。说明他们之间存在着一定的种属差异、基因多态性。拟了解青南地区的多房棘球绦虫虫株标本是否和北美洲或其他州流行的虫株具有同源性或更为特殊的基因多态性,还需要扩大样本量进行更进一步的分析。
总之,由于棘球绦虫有很多中间宿主和终宿主,且生活史复杂,较易受环境影响,种株分离也将越来越细。2005年肖宁等[14]为一个新种系统描述,命名为石渠棘球绦虫(Echinococcusshiquicus),已知青南高原存在藏狐石渠棘球绦虫及高原鼠兔石渠棘球蚴感染,且可观察到石渠棘球蚴具体形态[15]。而在本研究中可以看到下载所得的E.shiquicus(AB208064)虫株在系统发育树中独成一枝,与我们的多房棘球绦虫株、细粒棘球绦虫株和标本虫株间遗传距离相对较远,亲缘性相对不高,所以初步推断其具有自己的系统发育学特点。
参考文献
[1]White DJ,Wolff JN,Pierson M,et al.Revealing the hidden complexities of mtDNA inheritance[J].Mol Ecol.2008,17(23):4925-4942.DOI:10.1111/j.1365-294X.2008.03982.x.
[2]Nakao M,SaKo Y,Yokoyama N,et al.Mitochondrial genetic code in cestodes[J].Mol Biochem Parasitol,2000,111(2):415-24.
[3]Thompson JD,Gibson TJ,Plewniak F,et al.The CLUSTAL-X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J].Nucleic Acids Research,1997,25(24):4876 -82.
[4]Ho JW,Adams CE,Lew JB,et al.SeqVis:visualization of compositional heterogeneity in large alignments of nucleotides[J].Bioinformatics,2006,22(17):2162-2163.
[5]Swofford DL.PAUP*:Phylogenetic Analysis Using Parsimony(and Other Methods)4.0 Beta[M].version 4.Sunderland:Sinauer Associates,2002,1-142.
[6]Tamura K,Dudley J,Nei M,et al.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software version 4.0[J].Mol Bio Evol,2007,24(8):1596-1599.
[7]Xia X,Xie Z.DAMBE:Software Package for Data Analysis in MolecularBiology and Evolution[J].J Hered,2001,92(4):371 -373.DOI:10.1093/jhered/92.4.371.
[8]Posada D,Crandall KA.MODELTEST:testing the model of DNA substitution[J].Bioinformatics,1998,14(9):817 -818.DOI:10.1093/bioinformatics/14.9.817.
[9]Ronquist F,Huelsenbeck JP.MrBayes 3:Bayesian phylogenetic inference under mixed models[J].Bioinformatics,2003,19(12):1572-1574.
[10]王虎,Peter M Schantz,刘凤洁,等.青海省人与动物多房棘球绦虫的感染[J].中国寄生虫病防治杂志,2000,13(2):121-122.DOI:10.3969/j.issn.1673-5234.2000.02.015.
[11]何多龙.青海省包虫流行与防治[J].中国人兽共患病杂志,2000,16(4):101-102.DOI:10.3969/j.issn.1002-2694.2000.04.037.
[12]付永.青海省称多县野生狐狸棘球绦虫COⅠ基因多态性分析[J].青海大学学报(自然科学版),2013,31(5):65-68.
[13]朵红,黄福强,冯凯,等.青海省藏犬和野生狐狸棘球绦虫感染调查及虫种分子鉴定[J].寄生虫与医学昆虫学报,2012,19(4):220-227.
[14]Xiao Ning,Qiu Jiamin,Nakao Minoru,et al.Echinococcus shiquicus n.sp.,a taeniid cestode from Tibetan Fox and plateau pika in China[J].Int J Parasitol,2005,35(6):693-701.DOI:1 0.1016/j.ijpara.2005.01.003.
[15]曾诚.青海省青南高原棘球绦虫新种-石渠棘球绦虫的形态学观察[J].现代预防医学,2013,40(7):1354-1356.
收稿日期2015-01-05
DOI:10.13452/j.cnki.jqmc.2015.04.004
中图分类号R383.3
文献标识码A
※:国家自然基金地区项目(No81360255);春晖计划项目(No2012-Z082)
李超群(1992~),女,汉族,河南籍,青海大学医学院2014级科学班研究生.※※:通讯作者,教授,博士,Email:qhmccdp@163.com
