上海市4种主要乔木的光合生理特征研究
2016-01-16顾志明
顾志明
(上海久盛建设监理咨询有限公司,上海 200011)
上海市4种主要乔木的光合生理特征研究
顾志明
(上海久盛建设监理咨询有限公司,上海 200011)
摘要:利用LI-6400红外气体分析仪测定了上海城市森林中常见的4种落叶阔叶树种的光合生理日变化参数,对其光合生理参数与环境因子的相关显著性等进行了比较分析。结果表明:4个树种夏季净光合速率(Pn)日变化均为双峰型,具有明显的“光合午休”现象,峰值分别出现在11:30和15:30,白玉兰具有较高的日均净光合速率,栾树日均净光合速率较低,白玉兰和榉树具有较高的蒸腾速率,栾树蒸腾速率相对较低;4个树种气孔导度日变化在测定时段均为双峰型,榉树、栾树、白玉兰的午间Pn的下降因子为气孔限制,悬铃木在午间的Pn下降为非气孔限制。影响光合速率的主要因子是光合有效辐射、气孔导度和胞间CO2浓度等。
关键词:城市森林;光合速率;生理生态
中图分类号:S718.43
文献标识码:A
文章编号:编号:1674-9944(2015)09-0050-03
收稿日期:2015-07-08
作者简介:顾志明(1963—),男,上海人,工程师,主要从事园林植物景观生态方面的研究工作。
1引言
城市森林是改善城市环境、提高人居环境质量的重要资源。近年来,国内外许多学者们对城市植物的研究主要集中在植物群落的配置方面,针对生态功能定量化的研究还很少[1-3]。植物通过光合作用积累有机物质,光合作用强弱直接影响着生态系统水热平衡和物质循环过程。由于实验条件的限制,以往的研究多集中于农作物、花卉、草本植物和天然林等方面,对人类密集的城市区域内绿地植被光合生理生态特性及影响因子的研究报道较少[4]。本文以上海常见4种落叶树种为对象,对城市森林主要乔木绿化树种的光合特性进行测定,揭示它们光合作用的基本生理生态特性,找出影响其作用发挥的主要因子,旨在为城市森林固碳释氧能力的定量化研究和城市森林绿化树种的合理配置养护提供科学依据。
2研究区域与研究方法
2.1 自然概况
上海市位于30°40′~31°53′N,120°51′~122°12′E,地处长江三角洲的东缘,南北海岸线的中点,长江入海口,属于长江三角洲以太湖为中心的碟形洼地的东缘,属北亚热带季风性气候,雨热同期,日照充分,雨量充沛,年均降水量1048~1138 mm。上海市夏季酷热,冬季湿冷,年均温15.2~15.9 ℃,极端最高气温40.8 ℃,极端最低气温-2.1 ℃,春秋较短,冬夏较长。
2.2 研究方法
2.2.1供试材料
在上海市建城区中选择典型城市森林功能类型样地(延中绿地和中山公园)内主要落叶阔叶树种:白玉兰(Magnoliadenudata)、榉树(ZelkovaschneiderianaHand-Mazz)、栾树(Koelreuteriapaniculata)和悬铃木(PlatanusLinn) 4种树木作为研究材料,测定树种在上海城市森林乔木绿化树种中占绝对优势。
2.2.2光合生理生态指标的测定
连续2年在7~8月的晴天,利用Li-6400便携式红外气体分析仪,于每日7:30~17:30每隔2 h测定1次,同一样地内选择健康成熟大小一致的植株5株,随机选取树中上部的叶片进行测定,每株取3-7片叶片,待系统稳定后,测定净光合速率(Pn,μmol CO2/ m2s)、蒸腾速率(Tr, mmol H2O/ m2)、气孔导度(Gs,mol H2O/ m2s)、胞间CO2浓度(Ci,μmol CO2/mol)、叶片温度(Tl, ℃)及主要环境因子,包括水汽压亏缺(VPD,kPa)、大气CO2浓度(Ca,μmol CO2/mol)、光合有效辐射(PAR,μmol/ m2s)、气温(Ta, ℃)和相对湿度(RH,%)等参数。
2.3 数据分析
使用Excel 2007 软件进行数据处理,Origin 8.5绘图,采用SPSS 17.0 软件进行数据统计与分析。
3结果与分析
3.1 4种乔木树种的净光合速率(Pn)日变化
植物生长在特定环境中,并与环境之间不断地进行着能量和物质交换。由于光照、温度和水分等环境因子在一天中发生明显的变化,因此,植物的光合速率也呈现出相应的变化规律。由图1可知,4个树种净光合速率(Pn)日变化曲线在测定时段均为双峰型,上午随着气温(Ta)和光合有效辐射(PAR)的增大,Pn随之升高,之后随着温度和有效辐射的继续增大而降低,下午光强逐渐减弱,Pn有一定程度的增大,最后随着PAR和Ta的降低,Pn随之减小,第一个峰值均出现在11:30,第二个峰值出现在15:30,并在13:30均具有明显的“光合午休”现象[5],可能是由于午间强烈的太阳辐射使气温升高,相对湿度湿度降低,蒸腾强烈,植物体内水分不足引起气孔关闭导致Pn下降[2],榉树的光合速率日变化曲线变幅最大,栾树的光合速率日变化曲线变幅相对较小,Pn午间降低的程度不同说明4个树种受环境因子的抑制各不相同;白玉兰具有较高的日均净光合速率,栾树日均净光合速率较低。
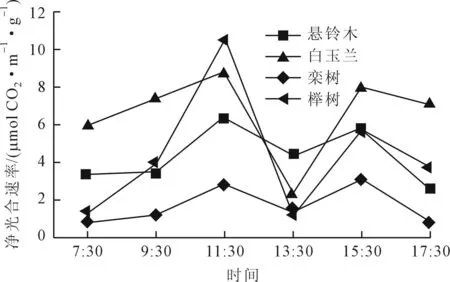
图1 4种树木树种净光合速率(Pn)日变化曲线
3.2 4种乔木树种的蒸腾速率(Tr)和气孔导度(Gs)日变化
白玉兰、榉树、栾树、悬铃木4种树木的蒸腾速率(Tr)日变化均为双峰曲线(图2a),白玉兰、栾树第一个峰值出现在9:30,悬铃木、榉树的第一个峰值出现在11:30,4个树种第二个峰值均出现在15:30。从蒸腾速率日变化来看,悬铃木、白玉兰在中午前后(9:30~15:30)数值较高,上午和傍晚较低,而榉树在午间(11:30~13:30)数值较高。榉树和栾树在午间呈现蒸腾速率的轻微下降,可能是由于中午温度高,失水过多,导致气孔部分关闭引起[5];其它3个树种在中午前后Tr维持在较高的水平,应该是植物通过提高蒸腾速率来降低叶温实现叶片各器官的正常功能。蒸腾速率在一定程度上反映了植物适应逆境调节水分损失的能力和适应干旱环境的能力[6],白玉兰和榉树具有较高的蒸腾速率,而栾树蒸腾速率较低,推测相同环境下,白玉兰、榉树比另2种树单位叶面积要消耗更多的水分,因而白玉兰和榉树在单位面积里失水量更大,必须及时浇水,才能保证植株水分的充足供应。
4个树种的气孔导度(Gs)日变化曲线均为双峰型(图2b),Gs与其蒸腾速率(Tr)日变化规律基本相似,早上可能由于气温较低,致使CO2扩散能力降低,叶片的Gs值相对较小。随着光强的增大,气孔缓慢张开[6],Gs随之上升,在Pn和Tr增大到一定程度后,Gs显现不同程度的下降。4个树种峰值出现的时间和其对应的Tr峰值出现的时间相同(图2A、2B)。
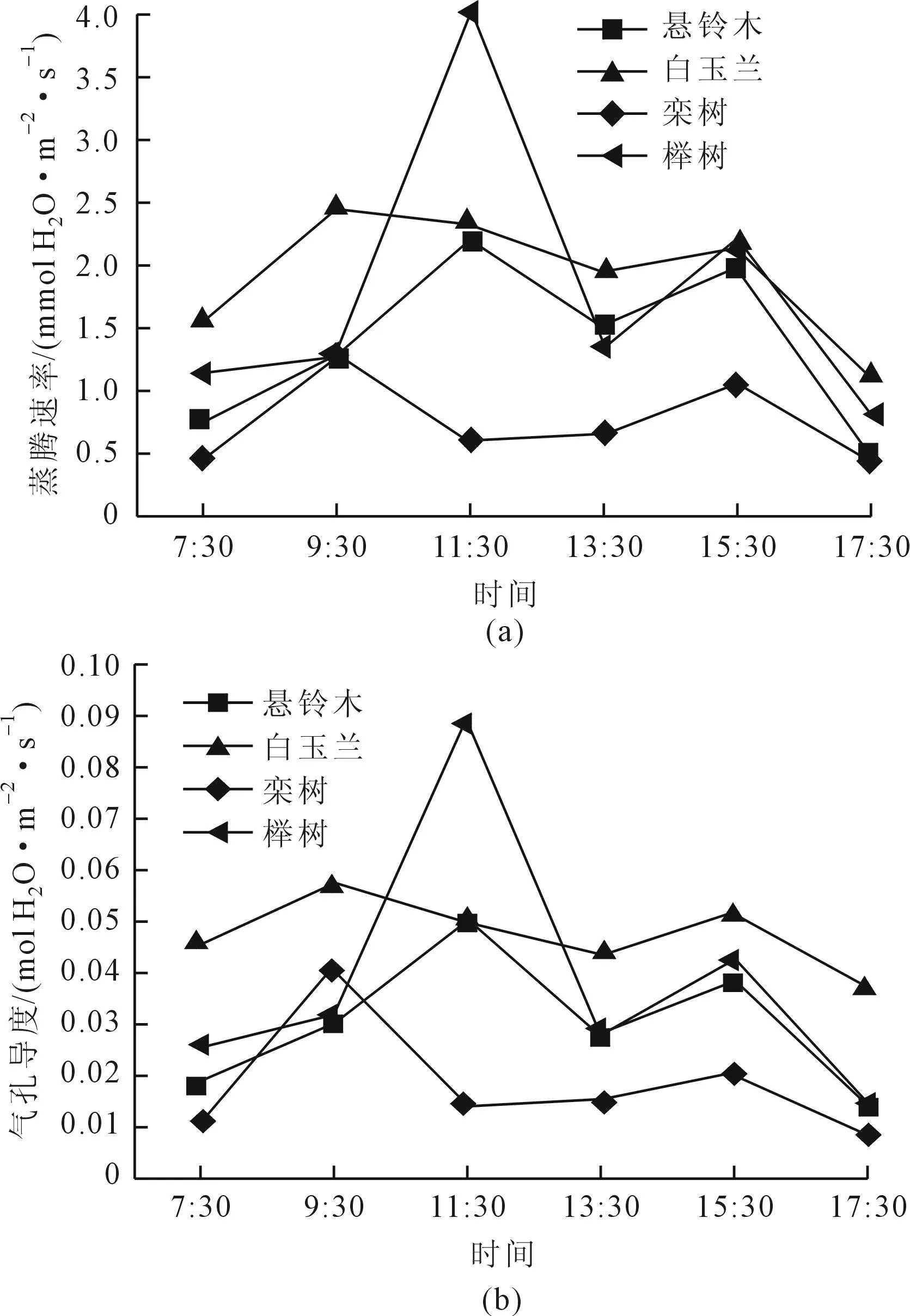
图2 不同树种蒸腾速率(Tr)和气孔导度(Gs)日变化曲线
3.3 4个树种光合作用的环境因子相关性分析
3.3.1净光合速率(Pn)和环境因子的相关性分析
如表1所示,除白玉兰外,其他3个树种Pn与PAR、Ta呈正相关,说明光合有效辐射和气温是这3个树种光合速率的主要影响因子。3个树种均与RH、Ca呈负相关,说明一定程度上大气CO2浓度和相对湿度阻碍了植物光合作用的进行[7]。另一方面,除白玉兰外,另3个树种Pn与Vpd、T1均为正相关,但相关性不明显。
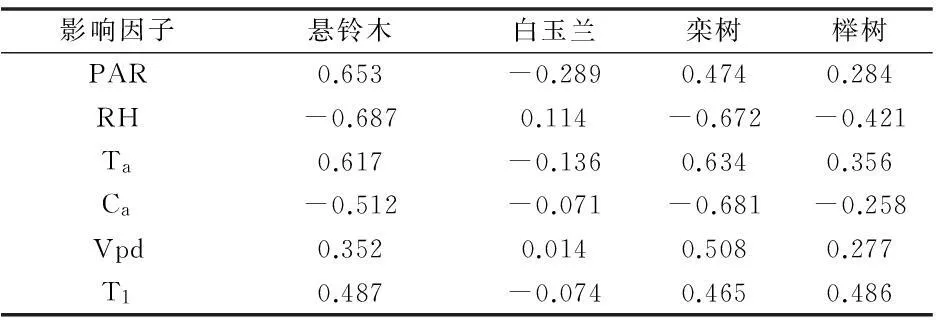
表1 不同树种净光合速率(Pn)与光合参数相关性
注:** 在0.01 水平(双侧)上显著相关。* 在 0.05 水平(双侧)上显著相关
3.3.2蒸腾速率(Tr)与环境因子相关性分析
由表2可知,4个树种Tr与PAR、Ta、Vpd、T1呈正相关,而与RH、Ca均为负相关,说明空气相对湿度越大、大气CO2浓度越高则植物蒸腾速率越小。
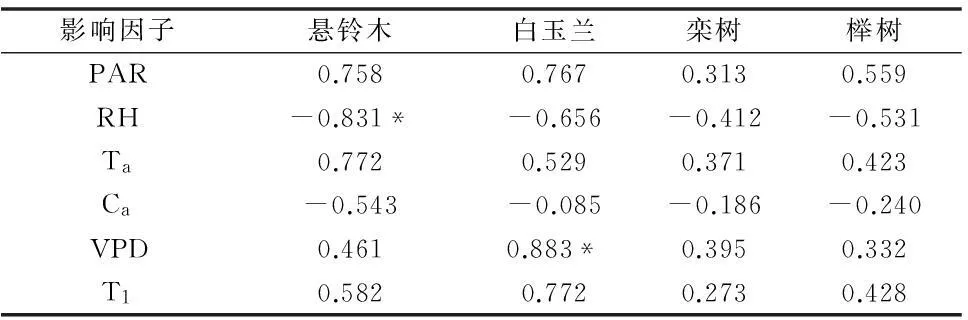
表2 不同树种蒸腾速率(Tr)与光合参数相关性
注:** 在0.01 水平(双侧)上显著相关。* 在 0.05 水平(双侧)上显著相关
4结论与讨论
4个树种净光合速率(Pn)日变化均为双峰型,存在明显的“光合午休”现象,强光、高温、低湿等环境因子引起的气孔部分关闭或光抑制的发生和叶片光合器官功能受到损害都有可能是引起这种现象的主要原因[5-7],“光合午休”现象虽然在夏季比较普遍,但并不是每种植株每个部位的叶片都存在这种现象的,且有的植物只有夏季才会出现“午休”现象,冬季则Pn日变化为单峰曲线[8]。根据Farquhar和Sharkey气孔限值分析的方法和判断标准[9],榉树、栾树、白玉兰午间Pn的下降为气孔限制,悬铃木在午间的Pn下降为非气孔限制,说明在一定光强和气温下,悬铃木净光合速率的下降是由于光合组织的功能受到了损害,而榉树、栾树和白玉兰下降是由于植株蒸腾作用强烈、叶片失水过多导致气孔导度(Gs)降低引起的,因此,适当控制环境因子是提高植物光合作用产量的重要举措,如通过遮光来降低光照强度,适当的灌溉来提高植物水分的补充和调节周边环境空气的湿度等。植物主要通过蒸腾作用消耗水分,蒸腾速率决定植物体内水分的平衡,影响着植物的水分利用效率,因而叶片的蒸腾速率一定程度上反映出植物适应干旱环境并调节水分损失的能力[9]。相同环境下,白玉兰、榉树单位叶面积要消耗更多的水分,夏季高温期间的园林绿化养护中应结合植物的冠层结构等特性进行合理的灌溉管理,以保证植株有充足的水分供应。
植物的水分利用效率(WUE)反映了植物的耗水和耐旱特性[9],榉树、栾树对环境因子的响应比另2种树种更为敏感,白玉兰能够更有效地利用土壤水分。另外,分析植物水分利用效率还需要考虑植物的灌溉状况,在干旱半干旱条件下,植物的水分利用效率受到土壤水分供给的影响。光饱和后植物对阳光的利用效率随着光强的升高而降低,白玉兰具有较高的LUE,在相同的环境条件下,白玉兰单位叶面积能够固定更多的碳量,对太阳能具有更强的光能利用率。
对4种树木净光合速率(Pn)、蒸腾速率(Tr)与环境因子的相关性分析认为,影响树种光合速率的主要因子为光合有效辐射、气孔导度和胞间CO2浓度等,而各因子对不同树种的影响机理还有待于进一步研究。
参考文献:
[1] 何春霞,李吉跃,郭明,等.4种乔木叶片光合特性和水分利用效率随树高的变化[J].生态学报,2008(7):3008~3016.
[2] 刘磊,李旖旎,夏磊,等.重庆地区黄葛树夏季光合日变化与主要环境因子的关系[J].西南师范大学学报:自然科学版,2013(3):120~126.
[3] 张小全,徐德应.18年生杉木不同部位和叶龄针叶光响应研究[J].生态学报,2001,21(3):409~414.
[4] 王得祥,刘建军,王翼龙,等.四种城区绿化树种生理特性比较研究[J].西北林学院学报,2002,17(3):5~7.
[5] 常杰,葛滢,陈增鸿.青冈常绿阔叶林主要植物种叶片的光合特性及其群落学意义[J].植物生态学报,1999,23(5):393~400.
[6] 陈志辉,张良诚.温州蜜桔叶片光合速率的日变化[J].植物生理学报,1994,20(3):263~267.
[7] 姜小文,张秋明,易于军,等.四季柚的光合特性研究[J].湖南农业大学学报,2003,29(5):397~401.
[8] 李海梅,何兴元,陈玮.沈阳城市森林主要绿化树种-丁香的光合特性研究[J].应用生态学报,2004,15(12):2245~2249.
[9] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982(33):317~345.
