人工林下与林缘紫茎泽兰种子形态与萌发特征比较研究
2016-01-15朱晓媛王崇云周新茂
潘 燕, 朱晓媛, 王崇云, 周新茂, 高 杰
(1.云南大学生态学与地植物学研究所, 昆明650091;2.贵州省关岭自治县水利局, 贵州 安顺561300; 3.云南大学生命科学学院, 昆明650091)
紫茎泽兰(Ageratina adenophora R.M.King & H.Rob)是一种危害严重的世界性恶性杂草[1-2],种子小而轻,成年植株年产种子10 000~100 000枚,种子具冠毛,适于长距离的风力传播[3-4],与同属本地种相比,紫茎泽兰产生较多的种子,具有较高的发芽率[5]。紫茎泽兰种子萌发需要光照[6],同时光质和光强影响其种子发芽[7],干扰很可能是通过改善光环境促进种子萌发从而导致紫茎泽兰入侵[8]。此外,储藏条件、地面覆盖物、pH值及温度等也影响着紫茎泽兰的种子萌发[9-10]。紫茎泽兰种子寿命较长[10],能形成长效土壤种子库[11]。
种子繁殖是紫茎泽兰种群定居及远距离传播的主要途径[9]。在同地分布于干香柏(Cupressus duclouxiana)人工林的种群中,比较分析林下和林缘紫茎泽兰个体的种子形态与萌发特征,进一步认识紫茎泽兰在不同环境中的入侵扩散特征,为入侵防治提供科学依据。
1 材料与方法
1.1 种子采集
在昆明金殿后山干香柏人工林中,选择林下和林缘2种生境,林下郁闭度85%,林缘郁闭度<15%,于2013年4月间,野外随机采集紫茎泽兰30株植株顶部的成熟花序置于纸杯中编号保存,带回实验室备用。室内随机挑选黑色饱满且冠毛完好的种子(瘦果)备用。
1.2 种子形态特征测定
1.2.1 千粒重
按4分法随机选取采自2种生境中饱满的紫茎泽兰种子各3 000粒,1 000粒为一组,用精度为0.000 1g的电子天平(ME 54)称重,重复3次。
1.2.2 种子长度、宽度、冠毛数量和冠毛长度
2种生境紫茎泽兰种子混合样品中,随机选取饱满且冠毛完好的种子各50粒。双目体视显微镜下对种子进行形态观察,利用尼康XM-1200F数码相机对种子进行拍照,统计种子的冠毛数量。采用Image J 1.43软件对种子(瘦果)长度和宽度进行测量,从每粒种子的冠毛中随机挑选中等长度的2条并对其进行测定,记录长度,计算种子长宽比和种子长/冠毛长。具体方法参照戴志聪等[12]的研究。
1.3 种子萌发实验
分别随机选取采自林下和林缘2种生境的适量黑色饱满种子,先用0.1%的次氯酸钠(NaClO)溶液消毒15min,然后再在25℃温箱中用双蒸水浸泡24h后备用。在已浸泡好的种子中随机选取50粒为一组,共3个重复。将每50粒种子均匀放入垫有2层中速定性滤纸(已浸湿)且直径为10cm的玻璃培养皿中。培养皿用封口膜封住以减少水分蒸发。将培养皿分别放入设定为25℃的恒温培养箱中进行种子的萌发试验,12 h光照和12h黑暗交替培养。种子的萌发以种子长出的胚根达到2mm为标准。种子萌发过程中每天对种子萌发情况进行观察记录,并将已萌发的种子从培养皿中移除,每天适量补充一次双蒸水。试验周期为30d,连续7d未观测到萌发的种子视为该组萌发结束[13]。种子的萌发率(GC)表示在实验观察期间种子的萌发能力,根据下式计算[14-15]:
GC=∑Gt/N×100%(式中:Gt为萌发终期所有正常萌发的种子数量,N 为供试种子总数)。
萌发指数(GI)表示种子的萌发速度。
GI=∑Gt/Di(式中:Gi表示在第ti天(ti=1,2,3…)萌发的种子数,Di表示不同的萌发天数)。
1.4 数据分析
采用SPSS和Excel进行统计分析。
2 结果与分析
2.1 林下与林缘紫茎泽兰种子形态特征比较
2.1.1 种子千粒重
2种生境中紫茎泽兰种子重量有差异显著。林缘生境紫茎泽兰种子的千粒重为0.052 5g,林下为0.047 6g,林缘生境种子千粒重极显著高于林下生境(表1)。林缘生境中种子千粒重的变异系数(0.48%)大于林下生境(0.42%)。
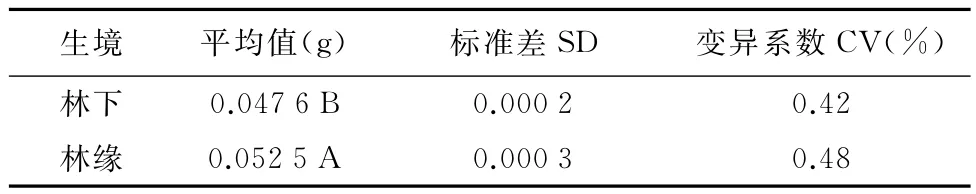
表1 林下与林缘紫茎泽兰种子千粒重比较
2.1.2 种子大小和冠毛特征
不同生境紫茎泽兰种子大小比较见表2。林下和林缘2种生境中紫茎泽兰种子长度均值分别为1.382 mm和1.378mm,种子长宽比均值分别为4.874和4.531,二者差异均不显著(p>0.050);林缘紫茎泽兰种子宽度均值(0.306mm)极显著高于(p<0.01)林下紫茎泽兰种子宽度(0.285mm)。
林下和林缘2种生境中紫茎泽兰种子冠毛数量均为10,冠毛长度均值分别为3.290mm和3.343mm,种子长/冠毛长均值分别为0.420和0.415,它们之间的差异均不显著(p>0.05)。
2.2 林下与林缘紫茎泽兰种子萌发特征比较
对不同生境中紫茎泽兰种子萌发率和萌发指数进行比较(图1)。林缘紫茎泽兰种子萌发率(92.00%)高于林下紫茎泽兰种子萌发率(84.67%),但是二者差异不显著(p>0.05)(图1a)。林缘和林下2种生境中的紫茎泽兰种子的萌发指数分别为25.95和17.93,前者极显著高于后者(p<0.01)(图1b)。
3 讨论与结论
林缘开阔生境中的紫茎泽兰种子重量和种子宽度均极显著高于林下郁闭生境中的紫茎泽兰种子重量和种子宽度,二者其余种子形态特征之间差异不显著。林缘开阔生境中的紫茎泽兰种子萌发率高于林下郁闭生境中的紫茎泽兰种子萌发率,但差异不显著;但前者种子萌发指数极显著高于后者的种子萌发指数。
为适应不同干扰程度的生境,紫茎泽兰种子表现出不同的形态特征,种子形态特征影响着种子的萌发和传播[16]。大的种子能够保证种子的萌发和幼苗的生长,而小的种子易于传播扩散[17]。在林缘干扰较强的生境中,紫茎泽兰的种子更大,且萌发能力更强,有利于种子繁殖定居。在林下更稳定的生境中,紫茎泽兰小种子质量影响了种子的生活力,但是却提高了种子的传播速度和范围,使种子的风力传播能力更强,为紫茎泽兰提供入侵可能性。

表2 林下与林缘紫茎泽兰种子形态特征比较(平均值±标准差)
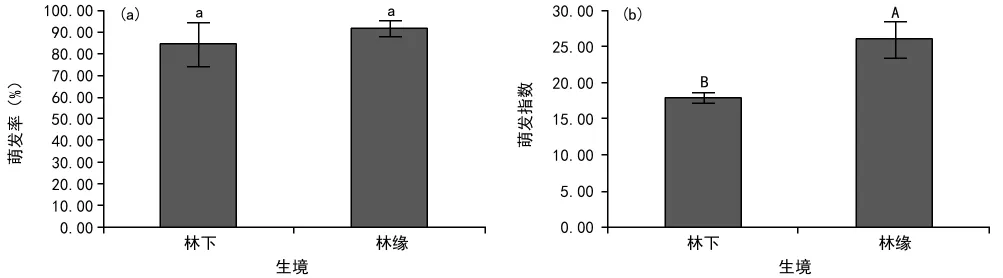
图1 林下与林缘紫茎泽兰种子萌发率(a)和萌发指数(b)比较
[1]尹芳,黄梅,徐锐,等.紫茎泽兰的危害及其综合利用进展分析[J].灾害学,2009,24(4):63-67.
[2]Muniappan R,Raman A,Reddy GVP.Ageratina adenophora (Sprengel)King and Robinson(Asteraceae)[M].In:Biological Control of Tropical Weeds using Arthropods.Cambridge University Press,2009:63-73.
[3]杨逢建,张衷华,王文杰,等.八种菊科外来植物种子形态与生理生化特征的差异[J].生态学报,2007,27(2):442-449.
[4]Sang W,Zhu L,Axmacher JC.Invasion pattern of Eupatorium adenophorum Spreng.in southern China[J].Biological Invasions,2010,12:1 721-1 730.
[5]韩利红,刘潮,郑玉龙.紫茎泽兰与3种同属本地植物种子特性比较[J].种子,2010,29(2):73-76.
[6]Auld BA,Martin PM.The autecology of E.upatorium adenophorum Spreng.in Australia[J].Weed Research,1975,15:27-31.
[7]倪文.环境因子对杂草紫茎泽兰种子发芽的影响[J].生态学报,1983,3(4):327-331.
[8]牛燕芬,冯玉龙,谢建磊,等.干扰强度对群落中紫茎泽兰种子萌发、幼苗定居和生长的影响[J].广西植物,2011,31(6):795-800.
[9]王文琪,王进军,赵志模.环境因素对紫茎泽兰种子萌发的影响[J].中国农学通报,2007,23(2):346-349.
[11]沈有信,刘文耀.长久性紫茎泽兰土壤种子库[J].植物生态学报,2004,28(6):768-772.
[12]戴志聪,杜道林,司春灿,等.用扫描仪及ImageJ软件精确测量叶片形态数量特征的方法[J].广西植物,2009,29(3):342-347.
[13]闫兴富,曹敏.光照对绒毛番龙眼种子萌发的影响[J].云南植物研究,2008,30(2):183-189.
[14]苏秀红,宋小玲,强胜,等.不同地理种群紫茎泽兰种子萌发对干旱胁迫的响应[J].应用与环境生物学报,2005,11(3):308-311.
[15]杨立学.俄罗斯大果沙棘(Hippophae rhamnoides L.)种子萌发特性[J].生态学报,2007,27(6):2 215-2 222.
[16]Fowler N.The role of competition in plant communities in arid and semi-arid regions[J].Annual Review of Ecological Systematics,1986,17:89-110.
[17]Chambers JC,Macmahon JA,Haefner JH.Seed entrapment in alpine ecosystems:effects of soilparticle size and diasporemorphology[J].Ecology,1991,72:1 668-1 677.
