黄海水温与沙海蜇丰度年际变化的相关分析*
2016-01-15赵亮李夏张芳
赵 亮 李 夏 张 芳
(1. 天津科技大学海洋与环境学院 天津 300457; 2. 中国科学院海洋研究所 青岛 266071)
近十年来, 在全球海洋的局部海域特别是近海一些重要的渔场和高生产力区出现了水母种群暴发的现象, 水母暴发已经成为重要的生态灾害之一, 对沿海工业、海洋渔业和滨海旅游业等都造成严重危害(孙松, 2012a)。虽然全球诸多海域出现水母数量增多的现象, 然而最近的研究显示, 全球水母的增加是否具有普遍性仍备受争论, 大量研究表明, 最近几十年来全球大约 27%的海域水母丰度出现增加情况(Condon et al, 2013)。无论如何, 水母数量的变化可作为海洋生态系统演变的指示, 探究水母多发的原因对于资源的可持续利用、海洋生态系统的健康发展具有重要意义。水母的数量及生态学角色越来越引起全球海洋生态学家的兴趣(Purcell, 2005; Purcell, 2007;Zhang et al, 2012)。自20世纪90年代末起, 大型水母沙海蜇频繁暴发于东亚海域, 包括中国渤海、黄海、东海北部、日本海及韩国海域(程家骅等, 2005; 丁峰元等, 2005; Kawahara et al, 2006; Uye, 2008; Zhang et al, 2012; Yoon et al, 2014; Sun et al, 2015)。我们根据沙海蜇平均丰度范围人为划分暴发程度, 例如平均丰度为 2—10ind./100m2为暴发年, 1—2ind./100m2为弱暴发年, 0—1ind./100m2为不暴发年。在2003年至 2013年十年间, 水母除 2008、2010、2011、2013年没有暴发外, 其余年份均为暴发年或弱暴发年(Sun et al, 2015)。经过我国科学家多年的研究, 对该种的暴发原因已取得重要进展。研究发现, 我国近海沙海蜇暴发原因和机制主要包括过度捕捞、富营养化、底栖生物的改变、气候变化等多个方面(仲霞铭等, 2004;严利平等, 2004; 周永东等, 2004; 程家骅等, 2004;张芳等, 2009; 孙松等, 2012a, b)。其中, 与研究全球水母与气候变化的关系一样, 研究气候变化与黄海沙海蜇丰度之间的关系需要对它们之间的相关关系做比较分析, 然而我国在此方面的研究尚未进行。在温度对沙海蜇生活史影响的研究中, 根据实验室的研究结果(Fenget al, 2015a, b)可总结出温度的季节变化对黄东海沙海蜇种群补充影响有以下推论: (1) 春季底层海水温度 10—18°C持续的时间长, 有利于水螅体横裂发生, 进而有利于当年水母暴发; (2) 夏季底层海水温度 18—25°C持续时间长, 有利于水螅体的足囊萌发, 有利于下一年的水母暴发。本文借助同化的海洋模式结果及黄海水母总丰度数据, 分析黄海温度自身变化以及特征温度段持续时间与沙海蜇丰度的关系, 讨论了特征温度的年际变化与沙海蜇丰度年际变化的关系, 验证以上室内实验结果的推论, 为黄海沙海蜇将来暴发机制的研究及预测提供基础资料。
1 数据与方法
本文采用的温度数据取自 JCOPE2 (Japan Coastal Predictability Experiment)同化数据。JCOPE2模型使用NCEP (National Centers for Environmental Prediction) /NCAR (The National Center for Atmospheric Research)再分析数据中的风应力、热通量和盐通量作为强迫场, 对多套卫星遥感数据和实测数据进行同化: 包括1992年11月至2002年1月的TOPEX/Poseidon、ERS (European Remote Sensing satellite)-1, 2和2002年1月至2008年1月的Jason-1、GFO (GeosatFollow-On)卫星高度数据同化得到沿轨海平面高度异常(SSHA); AVHRR/MCSST (Very High Resolution Radiometer/Multi-Channel Sea Surface Temperature)数据同化海表面温度(SST); GTSPP(Global Temperature-Salinity Profile Program)的Argo浮标和船舶观测资料同化温盐剖面数据。模型水平网格分辨率为(1/12)°×(1/12)°, 垂向上为 47个不等间距σ层(Miyazawaet al, 2008)。本文选用了1993年1月1日到2013年12月31日期间的日平均温度结果。
本文中所采用的沙海蜇丰度数据为南黄海全区平均丰度, 具体丰度的获得和计算方法参见Zhang等(2012)和Sun等(2015), 具体数据引自Sun等(2015)。本文在处理温度数据时对于 10—18°C持续时间的定义为: 从当年3月1号开始算起, 至9月30号结束,连续 5天温度在 10—18°C之间, 则计入持续时间的累积。对于 18—25°C持续时间的定义为: 从当年 3月1号开始算起, 至当年12月31号结束, 连续5天温度在18—25°C之间, 则计入持续时间的累积。选取的研究区域如图1所示, 包括南黄海和长江口外海域。

图1 研究区域Fig.1 Map of the study area
在利用模式数据分析之前, 首先采用实测数据检验模型数据的可靠性, 观测数据来自海洋局1993—1999年常规断面调查数据, 选取的站位见图1中黑色圆点所示。从模式中选取与观测数据时间和层次相匹配的点进行对比(图2), 模式给出温度数据与实测数据相关系数在95%置信度下达到0.99, 模式数据与实测数据呈现出一致的变化趋势, 两者具有很好的相关性, 说明模式给出的温度数据能较好反映真实温度的变化, 我们可以利用模式数据进行后续研究。

图2 JCOPE2模拟温度与现场实测温度对比Fig.2 Comparison between observation and JCOPE2 simulated temperature
2 结果与讨论
最近十年才有针对黄海海域水母总丰度的系统连续观测, 本文利用 2006—2013年数据, 讨论了温度以及温度持续时间的年际差异和水母多发的关系。为简单起见, 讨论温度影响时, 不考虑其他影响因素的作用。
2.1 沙海蜇的丰度与表底温度的相关性分析
将2006—2013年每年2月至8月黄海沙海蜇整个海区的平均丰度与表底层海水月平均温度做相关性分析, 得出表、底温度和南黄海水母丰度相关系数的分布结果(图 3, 4)。总体来看, 从冬季到夏季表层海水温度与水母丰度均呈正相关关系, 除7月份在朝鲜半岛近岸和 8月份在江苏近岸和长江口部分区域出现负相关关系外, 整个南黄海和长江口外海域, 尤其是南黄海中部海域和黄海潮汐锋区, 均呈现正相关分布。底层海水温度与水母丰度呈现与表层类似的相关分布, 2—7月整个区域呈现正相关趋势, 8月份在江苏近岸和长江口呈弱负相关关系。冬季(12—2月)表底相关系数高值区位于海州湾和苏北沿岸, 春季(3—5月)长江口外海域和苏北沿岸出现相对高值区, 夏季(6—8月)高值区出现在黄海中部, 近岸区域则表现为负相关。整体而言, 底层温度相比表层温度相关性高, 在春夏季, 近岸区域表现尤为明显。温度对沙海蜇生长的各个阶段都有很大的影响, Zhang等(2012)发现, 春夏季节, 黄海中部温度升高, 水母的丰度和生物量也都随之增加。春末夏初, 温度升高,有利于水螅体的横裂, 产生更多碟状幼体, 从而有利于当年水母多发, 而夏季温度继续升高则对足囊的萌发有利(Wanget al, 2015)。
Xu等(2013)通过对黄东海 SST卫星数据分析发现, 高温度有利于沙海蜇暴发。Ruiz等(2012)发现在地中海海域, 温度影响水母的暴发, 表现为夏季温度越高, 越有利于水母的暴发。Sullivan等(2001)发现美国罗得岛纳拉甘塞特海湾 1950—1999年间瓣水母丰度明显增加, 春季水温在此期间增加 2°C。这些研究都表明温度升高与水母增加有明显相关性, 本文从温度与水母丰度的关系进行研究, 也发现温度高的年份水母丰度相对就高的统计特性, 在全球变暖的大背景下, 水母存在增多的潜在性。
2.2 春季底层海水10—18°C持续时间与水母丰度关系

图3 2—8月表层温度与黄海水母丰度相关系数分布Fig.3 Distribution of correlation coefficients between SST and jellyfish abundance in the Yellow Sea from February to August
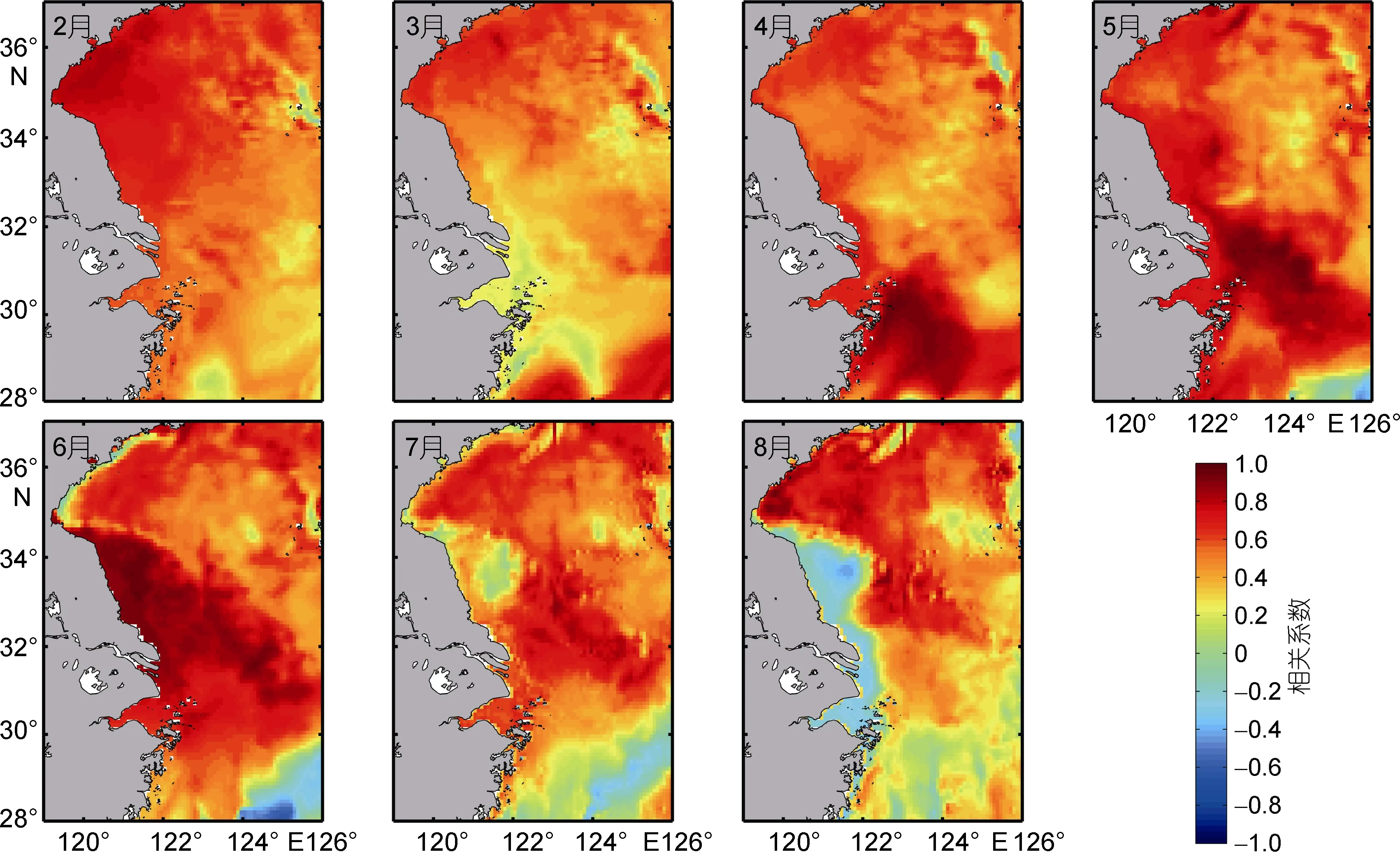
图4 2—8月底层温度与黄海水母丰度相关系数分布Fig.4 Distribution of correlation coefficients between bottom temperature and jellyfish abundance in the Yellow Sea from February to August
通过实验室控制实验发现, 春季底层海水10—18°C持续时间长有利于沙海蜇横裂(Fenget al,2015a)。图 5给出了 2006—2013年南黄海和长江口外海域春季底层 10—18°C持续时间分布, 图中可见大部分研究海区春季底层 10—18°C持续时间在90—140天之内, 黄海东南部及东海东北部持续时间较长, 在 160天以上, 海州湾 10—18°C持续时间在120—130天, 苏北近岸海域持续时间在 80—100天,长江口持续时间在100天左右, 南黄海中部持续时间较短, 约为60—80天。Sun等(2015)通过拖网调查可知长江口及其附近海域为沙海蜇的源地之一, 5月在122°—122.75°E, 31.5°—33°N 海域发现碟状幼体和幼水母。罗晓凡等(2012)利用 POM(Princeton Ocean Model)模型通过质子追踪溯源实验认为水母可能的源地为长江口、苏北沿岸以及海州湾。因此, 本文主要关注黄海近岸海域和长江口海区春季底层10—18°C持续时间分布。在海州湾附近, 2007、2009年10—18°C持续时间较长, 约130天, 而2010、2011年持续时间相对较短, 在 115天左右。春季底层10—18°C 持续时间在 2007、2009年比 2010、2011年多将近15天。在苏北沿岸区域, 2006、2007年持续时间较长, 约110天, 2010、2013年持续时间相对较短, 在98天左右。在长江口海区, 10—18°C持续时间在2006、2007年较长, 约110天, 2010、2011年相对较短, 约95天(图5)。根据2003—2013年的水母观测数据, 黄海水母除 2008、2010、2011、2013年为不暴发年外, 其余年份均为水母暴发年或弱暴发年,春季底层 10—18°C持续时间的年际差异与水母多发表现出较好的对应关系。
根据 Sun等(2015)研究, 2006、2007、2009、2012年是水母相对多年, 2008、2010和2011年较少, 其中2009年是极高值年, 而2011年是极少年。将2009年与2011年做对比, 在长江口, 春季底层10—18°C持续时间2009年比2011年多5天, 在海州湾2009年比2011年多17天。由此可以看出, 黄海沿岸区域及长江口附近海域春季底层10—18°C持续时间越长确实有利于当年水母丰度增多。
将春季底层10—18°C持续时间与水母丰度做相关分析, 结果表明, 长江口外、海州湾近岸海域以及南黄海锋区区域, 春季底层 10—18°C持续时间与水母丰度呈现正相关关系(图 7a), 这个区域春季底层10—18°C持续时间越长, 当年南黄海水母丰度越高。Feng等(2015a)通过实验室实验发现, 在10—18°C的环境中, 沙海蜇水螅体开始进行横裂生殖, 产生碟状幼体, 实验室环境中一个水螅体可产生 1—6个碟状体不等, 并且持续时间越长, 产生的碟状体也越多,此温度范围内水螅体的横裂率达到了80%。本文的统计结果与实验室实验结果一致, 即 10—18°C持续时间越长, 越有利于提高水螅体横裂率, 增加碟状体产量, 从而有利于本年度的水母旺发。
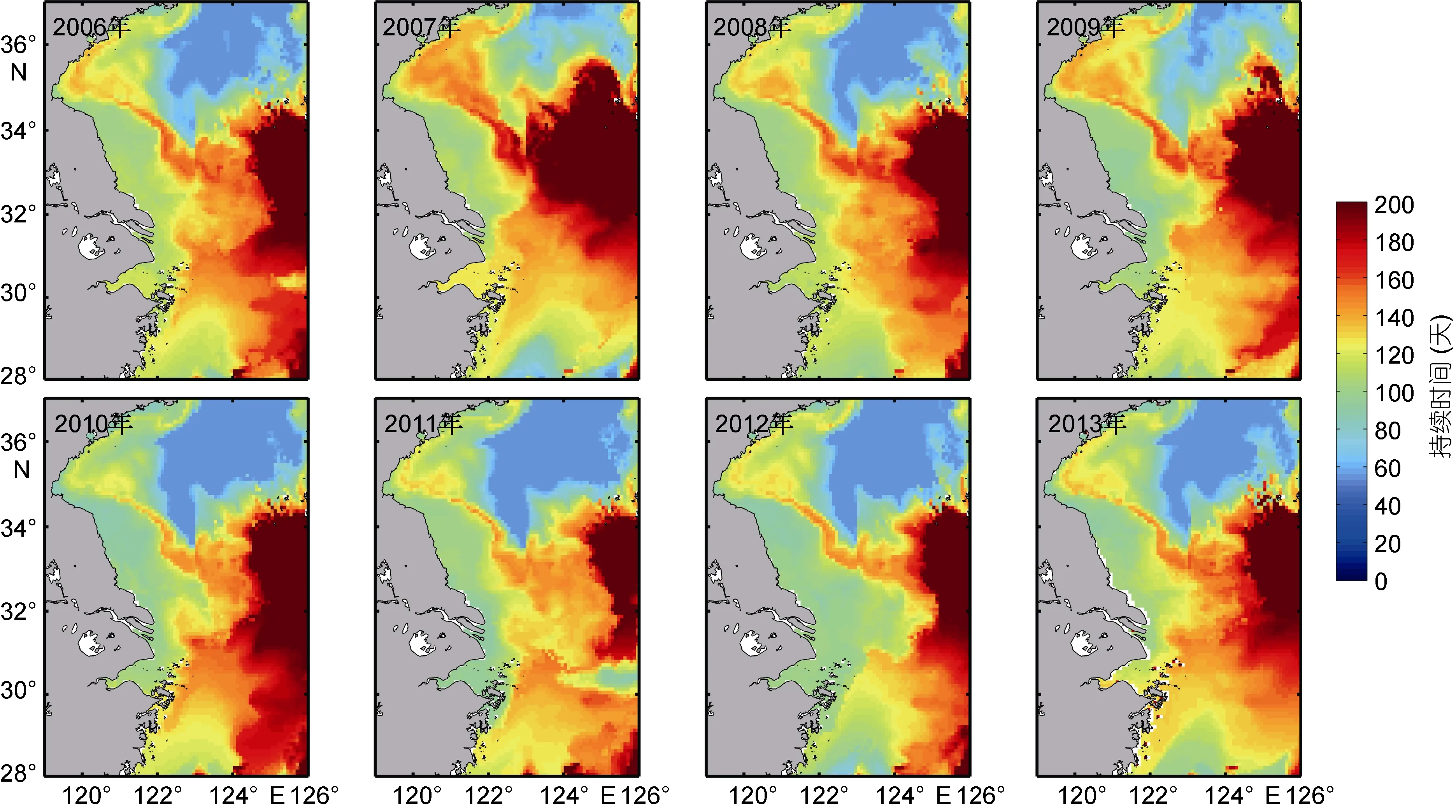
图5 2006—2013年春季底层10—18°C持续时间分布Fig.5 Distribution of 10—18°C duration in the bottom layer in spring from 2006 to 2013
2.3 夏、秋季底层18—25°C的持续时间与水母丰度关系
Feng等(2015b)根据实验室实验指出, 夏季底层海水 18—25°C持续时间长有利于沙海蜇足囊的生成。图6给出夏、秋季底层18—25°C持续时间分布,整个海域持续时间空间差异较大, 研究区域持续时间在 40—110天, 黄东海 30°N以北区域持续时间在100天以内, 海州湾18—25°C持续时间相对较长, 为60—80天。苏北近岸和长江口持续时间相对较长, 为50—80天。黄海中部冷水团区域终年温度低于18°C,不出现该温度段持续时间。在研究海域, 18—25°C持续时间年际变化较大: 在海州湾附近海域, 2005、2008年持续时间较长, 约70天, 2010年持续时间相对较短, 在63天左右; 在苏北近岸区域, 2006、2008、2011年 18—25°C持续时间较长, 约75天, 2007、2010年持续时间相对较短, 在 60天左右, 夏秋季底层18—25°C 持续时间 2006、2008、2011 年比 2007、2010年多将近 15天; 在长江口区域, 2008、2011年18—25°C持续时间较长, 约80天, 2007年持续时间相对较短, 在 60天左右。Sun等(2015)观测表明,2007、2009、2012年为水母多发年, 而2008和2011年水母少发, 可以看出夏秋季底层 18—25°C持续时间越长, 有利于水螅体产生足囊, 从而对第二年的水母多发有利。以2008年和2010年为例, 在苏北沿岸海域, 夏秋季底层 18—25°C持续时间在 2008年比2010年长15天左右(图6), 对应的2009年为水母暴发年和2011年水母丰度较低。夏季底层18—25°C持续时间与来年水母丰度也表现出较好的对应关系。
将夏、秋季底层18—25°C持续时间与年水母丰度做相关分析(图7b)可看出, 夏、秋季底层18—25°C持续时间与水母丰度在黄海近岸都呈现正相关关系,特别是在长江口、苏北沿岸以及海州湾这几个水母补充可能的源地区域, 夏、秋季底层 18—25°C持续时间与水母丰度呈明显正相关关系。
孙松等(2012a)认为水母暴发的关键在于水螅体阶段, 通过对海月水母进行不同温度的实验发现, 在温度为 15°C的环境中, 水螅体的横裂生殖率和碟状体的产量更高, 将温度控制在 15°C的范围内, 水螅体会不断进行横裂繁殖、持续产生碟状幼体。当温度超过15°C之后, 在20—25°C范围内, 水螅体生长活跃, 但主要发展方向是通过足囊、出芽等生殖方式进行无性繁殖。本文研究的沙海蜇生活史与海月水母不同, 但不同生活史阶段对温度的响应表现出相似的规律性。

图6 2005—2012年夏秋季底层18—25°C持续时间分布Fig.6 Distribution of 18—25°Cduration in the bottom layer in summer and autumn from 2005 to 2012

图7 春季底层10—18°C持续时间(a)和夏秋季底层18—25°C持续时间(b)与水母丰度相关系数分布Fig.7 Distribution of correlation coefficient between jellyfish abundance and 10—18°C duration in bottom layer in spring (a) and between jellyfish abundance and 18—25°C duration in the bottom layer in summer and autumn (b)
水母暴发是一个多因素共同影响的结果。Zhang等(2012)发现, 春、夏季节, 黄海中部温度升高, 水母的丰度和生物量也都随之增加。春末夏初, 温度升高,有利于水螅体的横裂, 产生更多碟状幼体, 从而有利于当年水母多发, 而夏季温度继续升高则对足囊的萌发有利(Wanget al, 2015)。Dong等(2015)讨论了光照和盐度对沙海蜇足囊繁殖的影响, 发现光照和盐度对沙海蜇足囊繁殖都有影响, 沙海蜇足囊繁殖的盐度范围为10—40, 对盐度环境的要求不高, 在不同的光照条件下沙海蜇繁殖在统计上有很小的差别。Feng等(2015b)则研究了饵料在水母多发中的贡献,发现春季饵料对水螅体横裂以及碟状幼体的产生作用有限。水母暴发是由多种因素共同影响的, 不同的影响因素在水母不同生活史阶段作用程度不同, 其中温度是一个很重要的因素。今后的一个研究方向则是借助模型综合分析温度、光照、饵料等各种因素对水母暴发的共同影响。
3 结论
在对水母暴发的原因研究中, 渔业过度捕捞导致以水母为食的鱼类(如大马哈鱼)减少, 意味着水母天敌减少, 有利于水母种群的生长, 从而导致水母暴发; 其次, 捕捞与水母饵料重叠的鱼类时, 饵料竞争减少, 给水母提供了暴发机会(Purcell, 2012)。除了富营养化、过度捕捞等因素, 气候变化导致的物理环境的变化也对水母暴发有重要影响, 温度和盐度的变化也会影响水母数量的变化(程家骅等, 2005; Xu et al,2013)。Feng等(2015a, b)通过大量室内实验, 讨论了温度在沙海蜇生活史不同阶段的作用。本文利用模型和现场观测数据采用统计方法对Feng等(2015a, b)的实验推论进行验证。
利用模式同化温度以及黄海水母总丰度, 分析了温度本身以及不同温度段持续时间与黄海沙海蜇丰度的关系, 统计结果表明: 水螅体栖息地的温度对水母暴发有重要作用, 春、夏季高温有利于水母的旺发。在长江口、苏北沿岸以及海州湾区域, 春季底层10—18°C 持续时间越长, 有利于于水螅体横裂, 水母的早期补充增大, 当年水母丰度就越高; 夏、秋季底层 18—25°C持续时间越长, 越有利于水螅体足囊萌发以及第二年水螅体的横裂补充, 导致第二年的水母丰度呈现增多趋势。本文进一步验证了Feng等(2015a, b)根据实验结果提出的两个推论: (1) 春季底层温度10—18°C持续时间长, 有利当年水母暴发; (2)夏季底层温度 18—25°C持续时间长, 有利于下一年的水母旺发。统计结果也进一步表明长江口海域、苏北浅滩和海州湾沿岸是黄海沙海蜇早期补充的重要源地。
研究表明白令海水母暴发主要是由于气候变化引起的区域性的生态体制转换, 而非对人类活动影响的响应(Shimomura, 1959; Brodeur et al, 1999)。温度变化和水母丰度关系的研究, 可以为未来气候变化背景下预测水母丰度的演变提供一种途径和思路。
丁峰元, 程家骅, 2005. 东海区夏、秋季大型水母分布区渔业资源特征分析. 海洋渔业, 27(2): 120—128
仲霞铭, 汤建华, 刘培廷, 2004. 霞水母(Cyanea nozakii Kisninouye)暴发与海洋生态之关联性探讨. 现代渔业信息, 19(3): 15—17
孙 松, 2012a. 对黄、东海水母暴发机理的新认知. 海洋与湖沼, 43(3): 406—410
孙 松, 2012b. 水母暴发研究所面临的挑战. 地球科学进展,27(3): 257—261
孙 松, 于志刚, 李超伦等, 2012. 黄、东海水母暴发机理及其生态环境效应研究进展. 海洋与湖沼, 43(3): 401—405
严利平, 李圣法, 丁峰元, 2004. 东海、黄海大型水母类资源动态及其与渔业关系的初探. 海洋渔业, 26(1): 9—12
张 芳, 孙 松, 李超伦, 2009. 海洋水母类生态学研究进展.自然科学进展, 19(2): 121—130
罗晓凡, 魏 皓, 王玉衡, 2012. 黄、东海水母质点追踪影响因素分析. 海洋与湖沼, 43(3): 635—642
周永东, 王永顺, 黄鸣夏, 2004. 浙江近海海域海蜇的增殖放流. 浙江海洋学院学报(自然科学版), 23(1): 28—30, 36
程家骅, 丁峰元, 李圣法等, 2005. 东海区大型水母数量分布特征及其与温盐度的关系. 生态学报, 25(3): 440—445
程家骅, 李圣法, 丁峰元等, 2004. 东、黄海大型水母暴发现象及其可能成因浅析. 现代渔业信息, 19(5): 10—12
Brodeur R D, Mills C E, Overland J E et al, 1999. Evidence for a substantial increase in gelatinous zooplankton in the Bering Sea, with possible links to climate change. Fisheries Oceanography, 8(4): 296—306
Condon R H, Duarte C M, Pitt K A et al, 2013. Recurrent jellyfish blooms are a consequence of global oscillations.Proceedings of the National Academy of Sciences of the United States of America, 110(3): 1000—1005
Dong J, Sun M, Purcell J E et al, 2015. Effect of salinity and light intensity on somatic growth and podocyst production in polyps of the giant jellyfish Nemopilema nomurai(Scyphozoa: Rhizostomeae). Hydrobiologia, 754(1): 75—83
Feng S, Zhang G T, Sun S et al, 2015a. Effects of temperature regime and food supply on asexual reproduction in Cyanea nozakii and Nemopilema nomurai. Hydrobiologia, 754(1):201—214
Feng S, Zhang F, Sun S et al, 2015b. Effects of duration at low temperature on asexual reproduction in polyps of the scyphozoan Nemopilema nomurai (Scyphozoa: Rhizostomeae).Hydrobiologia, 754(1): 97—111
Kawahara M, Uye S I, Ohtsu K et al, 2006. Unusual population explosion of the giant jellyfish Nemopilema nomurai(Scyphozoa: Rhizostomeae) in East Asian waters. Marine Ecology Progress Series, 307: 161—173
Miyazawa Y, Komatsu K, Setou T, 2008. Nowcast skill of the JCOPE2 ocean forecast system in the Kuroshio-Oyashio mixed water region. Journal of the Marine Meteorological Society of Japan, 84: 85—91 (in Japanese with English abstract)
Purcell J E, 2005. Climate effects on formation of jellyfish and ctenophore blooms: a review. Journal of the Marine Biological Association of the United Kingdom, 85(3):461—476
Purcell J E, 2007. Environmental effects on asexual reproduction rates of the scyphozoan Aurelia labiata. Marine Ecology Progress Series, 348: 183—196
Purcell J E, 2012. Jellyfish and ctenophore blooms coincide with human proliferations and environmental perturbations.Annual Review of Marine Science, 4(1): 209—235
Ruiz J, Prieto L, Astorga D, 2012. A model for temperature control of jellyfish (Cotylorhiza tuberculata) outbreaks: a causal analysis in a Mediterranean coastal lagoon.Ecological Modelling, 233: 59—69
Shimomura T, 1959. On the unprecedented flourishing of“Echizen Kurage” Stomolophus nomurai (Kishinouye), in the Tsushima Current regions in autumn, 1958. Bulletin of the Japan Sea Regional Fisheries Research Laboratory, 7:85—107 (in Japanese with English abstract)
Sullivan B K, Van Keuren D, Clancy M, 2001. Timing and size of blooms of the ctenophore Mnemiopsis leidyi in relation to temperature in Narragansett Bay, RI. Hydrobiologia,451(1—3): 113—120
Sun S, Zhang F, Li C L et al, 2015. Breeding places, population dynamics, and distribution of the giant jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) in the Yellow Sea and the East China Sea. Hydrobiologia, 754(1): 59—74
Uye S I, 2008. Blooms of the giant jellyfish Nemopilema nomurai: a threat to the fisheries sustainability of the East Asian Marginal Seas. Plankton and Benthos Research, 3(S):125—131
Wang Y T, Zheng S, Sun S et al, 2015. Effect of temperature and food type on asexual reproduction in Aurelia sp.1 polyps.Hydrobiologia, 754(1): 169–178
Xu Y J, Ishizaka J, Yamaguchi H et al, 2013. Relationships of interannual variability in SST and phytoplankton blooms with giant jellyfish (Nemopilema nomurai) outbreaks in the Yellow Sea and East China Sea. Journal of Oceanography,69(5): 511—526
Yoon W D, Lee H E, Han C et al, 2014. Abundance and distribution of Nemopilema nomurai (Scyphozoa,Rhizostomeae), in Korean waters in 2005–2013. Ocean Science Journal, 49(3): 183—192
Zhang F, Sun S, Jin X S et al, 2012. Associations of large jellyfish distributions with temperature and salinity in the Yellow Sea and East China Sea. Hydrobiologia, 690(1):81—96
