大豆灰斑病发生特点及抗病遗传育种研究进展
2015-12-25常芳国赵团结孔杰杰
程 伟,常芳国,赵团结,孔杰杰
(南京农业大学大豆研究所/国家大豆改良中心/农业部大豆生物学与遗传育种重点实验室(综合)/作物遗传与种质创新国家重点实验室,江苏南京 210095)
大豆灰斑病发生特点及抗病遗传育种研究进展
程 伟,常芳国,赵团结,孔杰杰*
(南京农业大学大豆研究所/国家大豆改良中心/农业部大豆生物学与遗传育种重点实验室(综合)/作物遗传与种质创新国家重点实验室,江苏南京 210095)
摘要从抗病育种角度出发,综述了大豆灰斑病病原菌生物学特点、生理小种分化与鉴别、病害流行规律与防控、大豆对灰斑病抗性的鉴定方法与抗病种质(基因)发掘、抗病常规及分子育种等方面研究进展,并提出了大豆灰斑病抗性育种研究应加强病菌致病机理及生理小种划分、抗病资源(基因)发掘、常规与分子育种相结合等对策。
关键词大豆;大豆灰斑病;生理小种分化;抗病育种;分子标记
大豆灰斑病又称褐斑病或蛙眼病,病原菌为(CercosporasojinaHara),称大豆短胖胞,属半知菌亚门真菌,其异名为Cercosporidiumsofinum(Hara)Liu & Guo,是世界性病害。1915年首次在日本报道,很多年后陆续在其他国家也有发现,如美国、中国、英国、德国等[1-3]。在我国黑龙江、吉林、辽宁、山东、河北等省灰斑病严重,少量在湖北、安徽、江苏等也有发生。其中黑龙江省发病最严重。历史上大豆灰斑病的发生与危害出现4次大流行。另外,大豆灰斑病的危害症状是侵染大豆的叶、茎、荚和子粒,使品质变劣,降低光合速率从而降低产量,造成轻者减产10%~30%,重者减产50%~100%,严重影响商品价值。笔者综述了大豆灰斑病发生特点及抗病遗传育种研究进展,提出了目前该方面研究中存在的问题,并展望了今后的研究方向,以期为大豆灰斑病防治提供参考。
1大豆灰斑病的生理小种与流行规律
1.1大豆灰斑病病原菌及其生理小种分化大豆灰斑病病原CercosporasojinaHara的分生孢子梗多为5~12根,不分枝,呈淡褐色。孢子有膝状屈曲,孢痕可见,具1~9个隔,大小(24~108)×(3~9) μm,形状、大小因培养条件不同略有差异。分生孢子萌发时从两端细胞长出芽管,有时也从中部细胞长出[4]。黑暗条件下有利于病菌的生长与产孢萌发,阳光直射对孢子萌发有明显抑制作用。在大豆叶葡萄糖培养基上该病菌生长最快,在小白菜琼脂培养基和番茄琼脂培养基上产孢量最大[5]。
大豆灰斑病菌具有明显的生理分化,存在高度的遗传变异。如Kim等[6]研究表明来自美国6个不同地区的132个大豆灰斑病原菌的标准化基因型多样性值在26%~79%,还发现阿肯色州的灰斑病菌存在有性繁殖现象,可见,生理小种划分工作重要。目前国内外多个国家和地区已对大豆灰斑病生理小种进行了划分(表1)。国内多个研究应用钢5151、九农1号、双跃4号、合交69-231、Ogden、桦南绿大豆(现改用合丰 22)为鉴别寄主,对黑龙江省为主的灰斑病菌样进行鉴定,分别鉴定出11~18个生理小种,其中1号小种是东北春大豆产区的优势小种[7-10]。马淑梅[11]对2006~2010年黑龙江省大豆灰斑病生理小种的监测结果表明,1号生理小种在各大豆产区出现频率最高,为50.5%,平均为40.1%;其次是7号,出现频率为35.9%,平均为26.2%。出现频率较高的还有6、10和11号生理小种,平均出现频率分别为18.8%、12.0%和9.6%。近期研究表明15号生理小种有增加的趋势[10-12]。
国外研究中,巴西采用10个鉴别寄主鉴别出22个生理小种[13],美国早期用一套由16个品种组成的鉴别寄主鉴别出11个生理小种[12]。近期,Mian等[14]研究了来自美国、巴西、中国的93个灰斑病病原菌对 38个大豆品种的致病性差异,结果67个美国菌株可归为13类,其中54%属于第1、2类群,15个巴西菌株则属于其中8个类群,中国的7个菌株分别属于7个类群。最终筛选出Davis、Peking、Kent、CNS、Palmetto、Tracy、Hood、Lincoln、Lee、Richland、S100、Blackhawk共12个鉴别寄主,鉴定出5~15号共11个生理小种。Scandiani等[15]首次在阿根廷发现11和12号生理小种(表1)。
1.2病菌的分子鉴定利用分子标记能较好地反映出病原菌之间的遗传关系。张俊华等[16]用引物对ITS1和ITS4PCR扩增我国东北大豆灰斑病菌16个生理小种的DNA,均得到600 bp片段,继而用HinfI、RsaI和HaeIII 3种限制性内切酶将扩增产物消化,得到15个多态性片段,可将16个生理小种区分开来。孙洪利等[17]利用10对多态性高且稳定性好的AFLP引物组合,对黑龙江省121个菌株进行分析,共获得148个多态性条带,以遗传距离小于0.81为界,将111个单元型划分为6个类群。丁俊杰等[18]利用11对SSR引物对黑龙江省的24个灰斑病菌菌株进行EST-SSR基因型分析,结果共检测出等位变异46.0个,平均每个位点为4.2个,相似系数范围为0.091~0.956,平均相似性系数达到0.589;聚类分析结果与大豆灰斑病生理小种间呈现出较高的相关性。

表1 大豆灰斑病生理小种鉴定结果汇总
Bradley等[19]对来自美国(44个大豆灰斑病生理小种)、巴西(10个)、中国(7个)、尼日利亚(1个)等地共62个菌株进行AFLP标记分析,结果发现小种之间平均遗传多样性值高达0.56。聚类分析发现2个来自乔治亚州的生理小种被聚集到一起,2个来自中国的生理小种被聚集到一起。在美国QoI杀菌剂可以有效控制大豆灰斑病的发生与流行,近期发现抗该类杀菌剂的大豆灰斑病菌,研究发现QoI-抗病包含G143A突变的细胞色素b基因,导致一种氨基酸替代丙氨酸甘氨酸。其聚合酶链反应和TaqMan探针开发可以有效辨别QoI-抗病和QoI感病[20-21]。
1.3病害流行规律与防控一般在苗期和花期只能见到少数病斑,但是在大豆鼓粒期后病情发展迅速。气象条件、感病品种布局及田间菌源量等条件都会影响大豆灰斑病的发生和流行。孢子萌发最适温度为28~30 ℃,温度范围为10~40 ℃,生长最适温度为25~28 ℃,萌发和侵染需要较高的湿度。pH为3~9时均可生长,最适温度下分生孢子在水中1 h即可萌发,pH为5~6时最适宜。影响病害发生程度的主要气象因子是湿度和降雨量,研究发现,东北地区大豆灰斑病发生与7月上旬、下旬降雨量呈显著正相关;降雨量大、周期长,相对湿度较大,病害发生会较重[22-23]。
刘惕若等[24]认为大豆灰斑病是一个多循环病害,叶、荚病情田间流行曲线均呈“S”型。还根据病粒率与气象因子的相关关系建立了早熟材料和中晚熟材料的灰斑病粒率预测多元回归方程,预测病粒率与实际调查的病粒率十分接近。顾鑫等[25]利用2005~2013年黑龙江省东部地区大豆灰斑病发生情况和相应的气象因子数据,采用逐步回归方法建立包括的回归预测方程,可对该地区大豆灰斑病发生情况进行中短期预测。
灰斑病流行与大豆耕种方式、病原菌、品种等因素有关。研究发现在美国大豆生产体系中,不施用杀菌剂条件下发病程度与耕作或免耕方式之间无显著差异。杀菌剂的应用显著减少了发病严重程度,产量涨幅为1%~17%。杀菌剂应用时,免耕耕作的大豆发病严重程度没有显著降低[26]。
2大豆抗灰斑病种质鉴定
2.1抗病鉴定方法与指标大豆灰斑病主要侵染幼苗、叶、茎、荚和种子[27]。带菌种子长出幼苗,子叶上病斑明显,多是半圆形、圆形、稍凹陷,深褐色。叶片发病,初为褪绿小圆斑,后逐渐扩展为边缘褐色,中央灰褐色或灰色、直径为1~5 mm的病斑,叶背中央有时密生灰色霉层,即为分生孢子梗和分生孢子。严重时导致叶片枯死并脱落。茎部发病形成椭圆形病斑,褐色。病轻时仅产生褐色小点。豆粒上病斑与叶斑相似,多圆形蛙眼状,也有的呈不规则形,边缘暗褐色,中央灰白色。
目前田间鉴定大豆灰斑病等级多分为5级。曹越平等[28]先按病斑的大小和病斑的有无将病斑分为5种:B 斑、M 斑、N 斑、S 斑、O 斑(直径> 3 mm、2~3 mm、1~2 mm、<1 mm、0 mm) ,并对上述5种病斑分别赋予 5、4、2、1、0 的权重系数;再按病斑数量的多少将病情分为5个级别:1、2、3、4、5 级。将病斑数量的级别数与病斑种类的权重系数相乘,其乘积即为病斑型级数。在进行病情调查时,以每个植株感染灰斑病较重的叶片为代表,表示该大豆植株的病情。而Sharma等[29]进行数量抗性鉴定时,根据整个植株叶片枯萎或坏死所占比例分0~9级(枯萎/坏死):枯萎、坏死均为0级;1级是 1%~10%/1%~5%;2级是10%~20%/6%~10%;3级是20%~40%/10%~20%;4级是40%~60%/20%~40%;5级是枯萎>60%,坏死>40%;6级为落叶达到33%;7级是落叶>66%; 8级是落叶>66%;9级为整个植株死亡。
2.2抗病种质筛选不同地区和来源的大豆材料对灰斑病菌的抗性存在差异,同一品种对不同的病原菌的抗性也有差异,所以选育抗病材料是抗病育种的基础。国内外均进行了大量研究。我国从5 000余份国内外大豆资源中鉴定出一大批优异抗源(表2)。如陈庆山[30]从50个主栽品种中筛选出抗8个以上生理小种的品种系17份,其中抗10个生理小种的代表品种有1572、巴24、农大13699等。研究发现进化程度低的原始材料中抗源较多,对166份野生大豆鉴定发现表现免疫和高抗的分别有57和38份,占58.30%,表明野生大豆中抗源丰富[31-32]。而对东北地区的916份大豆品种进行抗性鉴定,其中来自黑龙江省、吉林省表现免疫的分别有0.31%、1.55%,表现高抗分别有9.09%和11.46%[33];对我国南北方大豆品种系进行抗灰斑病鉴定,结果发现不同地区和来源的品种、品系均有抗灰斑病的材料,北方高抗材料比例较高,育成品种的比例要高于品系[34]。
国外也开展类似抗源筛选工作,如美国对世界各地1 686份种质资源对灰斑病强致病菌株C-32抗性鉴定,发现有 660 份为抗性(0~3级)材料,韩国资源抗源较多[35]。Mengistu等[36]在2009~2010年通过田间接种大豆灰斑病11号生理小种鉴定发现,Davis、PI532465、PI567302、PI56748160、PI490766、PI567330、PI567333A、PI567432A、PI567432B、PI567438、PI532463A、PI567477、PI567478、PI567490、PI63945、PI538383、PI538386B、PI567291、PI567319B、PI567316A、PI567333B品种都表现出抗性。
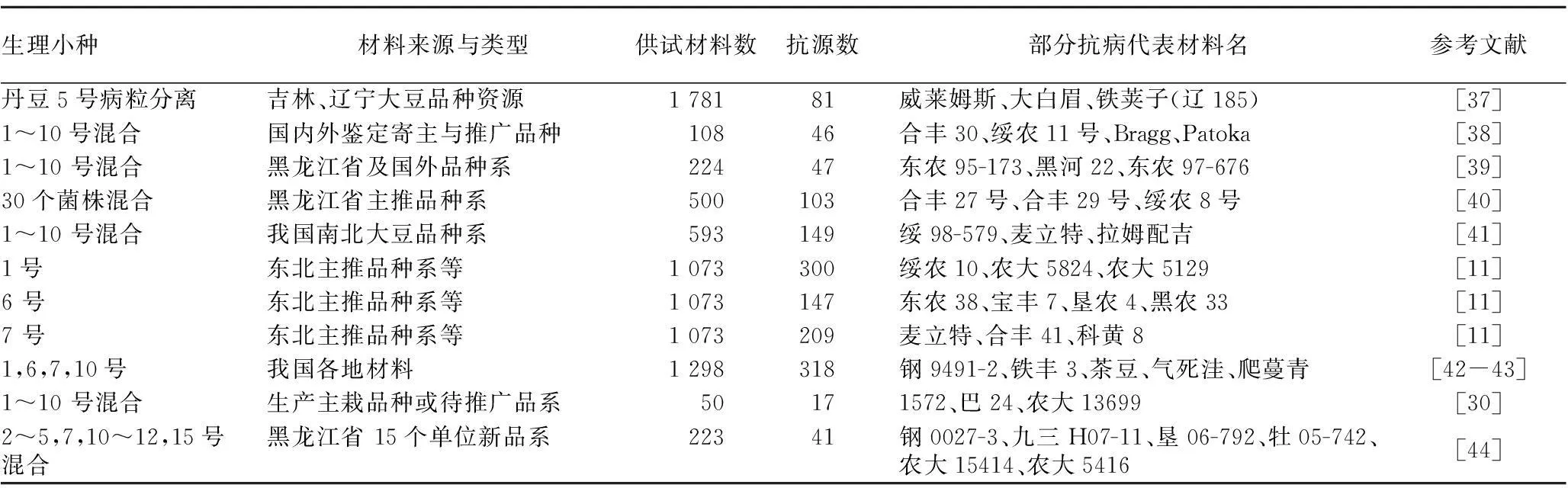
表2 大豆种质对我国灰斑病小种抗性鉴定结果汇总
3抗病育种研究
3.1常规育种美国开展大豆抗灰斑病育种工作较早,早期选育出的抗病品种有Wabash、Clark等。1986年美国西北部受到灰斑病的严重危害,对P746、Shillageet、PK719、Bragg、VLsoya2、Himso507、Himso10 7个品种进行抗病鉴定,发现其病情指数达到了50%。其后育成一批抗病品种,如Doles、Maxcy、Holladay和VLsoya2等。近年来美国公共研究机构的育种家开展了较全面的新品种抗性鉴定工作,又选育出一大批抗病品种[45]。
我国在20世纪80年代就开始了大豆抗灰斑病育种工作。刘忠堂[46]采用一次杂交,连续接种,定向选择的方法培育了合丰28、合丰29、合丰33、合丰34号等抗灰斑病大豆品种。迄今已育成推广合丰号抗病品种20多个。其他单位也育成一大批抗病品种(系),如东农42、东农43、1574、9674、593、垦农18、合辐93154-2、黑生101、建农1号、建丰2、北豆30、北豆35等。东农9674、合丰29和合丰34能抗黑龙江省8个以上生理小种[47-48]。由于东北地区大豆新品种审定要求对灰斑病的抗性达到中抗以上,目前大豆审定品种的抗性水平不断提高。如通过杂交育种获得的合农64和合农59均抗大豆灰斑病品种[49-50]。
3.2抗病基因发掘与分子标记辅助育种大豆对灰斑病生理小种的抗性是由单显性基因控制,国内发现控制1号、7号和15号小种的Hrcs1、Hrcs7和Rcs15抗病基因[51]。利用东农9674(高抗)×东农87-104(高感)F2群体发现3个与抗病基因Rf1的连锁的RAPD标记;为利于分子标记分析,还将抗病基因连锁的RAPD标记转化为共显性的SCAR标记SCS3620&580[52]。张文慧等[53-54]用东农40566(抗)和东农410(感)杂交衍生群体接种1号生理小种,发现位于C1连锁群上的3个SSR标记可能抗性基因连锁。Dong等[55]用Gang 95144-1和Gongjiao 9723-6 2个亲本杂交获得的F2代,接种大豆灰斑病7号生理小种鉴定其抗病基因,并命名为Hrcs7,将抗病基因定位在E连锁群(15号染色体),与Satt411的遗传距离为7.9 cM。姜翠兰等[56]研究发现,垦丰16对15号小种的抗性受1对显性基因控制,定名为Rcs1,抗病基因位于J连锁群Satt 529和Satt 547之间,遗传距离分别为18.5和-6.7 cM。对108份大豆新品系进行标记检测,Satt 547和Sat_224的检测准确率达到85%以上,可用于分子标记辅助选择育种和抗源筛选。丁俊杰等[18]以黑龙江省103份已鉴定抗大豆灰斑病3个生理小种的品种(系)为材料,同时利用7个标记(Satt565、Satt547、Satt431、Sct_186、SOYGPATR、Satt244、Sat_151)构建了供试品种(系)的分子身份证。
美国通过经典遗传学方法已发现控制1号、2号和5号小种的Rcs1、Rcs2、Rcs3、Rcs5等抗病基因。利用Blackhawk(S)和 Davis(R)2个亲本杂交获得的F2群体以及F2∶3,Wright和Wright6-Rcs3的近等基因系接种3号生理小种,发现位于J连锁群(16号染色体)2个标记Satt244、Satt547与Rcs3连锁[57];进一步加密标记,在Rcs3周围0.3 cM范围内11个SNP,其中AZ573TA150和AZ573CA393可以直接应用于分子标记辅助育种[58]。近期又将PI 594891、PI 594774对灰斑病的显性抗病基因精细定位在13染色体72.6 kb区间,包含5个注释基因,其中3个基因(Glyma13g25320、Glyma13g25340、Glyma13g25350)的序列在抗感材料间有差异。已对Glyma13g25340、Glyma13g25350基因SNP变异开发出KASP标记用于标记辅助育种[13]。
Sharma等[29]利用Essex×Forrest RIL群体的94个家系接种灰斑病2号生理小种,分别在21和42 d调查家系抗病反应,结果定位到2个对灰斑病表现部分抗性的主效QTL,1个在6号染色体(C2连锁群)Satt319标记附近,LOD=3.8,R2=52%,1个位于8号染色体Satt632附近,LOD=3.6,R2=15%。通过单标记分析,还在A1、B1、F、G、H、I、J、K、L、M 和O分子连锁群上检测到13个微效QTL。
4问题与展望
大豆灰斑病生理小种分化明显,国内外对小种划分都进行了许多研究,国内目前采用6个鉴别寄主材料,鉴别出多达18个小种[10];美国则提名了12个鉴别寄主材料,鉴别出11个小种[14]。由于二者没有相同鉴别寄主,中美鉴定出的小种间还难以一一对应,其致病性等也难以比较,制约了进一步的研究工作。因此,建立一套标准鉴别寄主品种十分必要,有待有关研究者合作完成。此外,大豆对灰斑病抗性鉴定体系也需要统一和完善,包括小种选用(混合或单用)、处理时期、调查鉴定标准等。如我国大豆灰斑病病情分级一般按病斑数量分4~6级[27],国外有将灰斑病病情分为9级,还有数量抗性鉴定则更复杂[28]。
大豆对灰斑病抗性育种工作已取得很大进展。分子标记辅助选择技术可弥补常规育种的不足,实现基因型的直接选择。目前抗大豆灰斑病基因发掘工作相对落后,虽然已发现多个抗病基因,但只有Rcs3和位于13染色体的抗病基因被精细定位[13,58],可用于分子育种的分子标记很少。高通量测序技术不仅使对大豆转录组和基因组进行细致全貌的分析成为可能,更使挖掘高通量多态性 DNA分子标记成为现实,该方面工作需要加强。目前,分子辅助选择育种只能对主基因控制性状的选择,对大量微效多基因控制的性状的选择方法与技术仍待进一步研发。另一方面,大豆灰斑病菌侵染大豆致病机理研究也有待加强,可发掘相关基因通过转基因育种提高大豆抗病性。
化学防治大豆灰斑病的效果好,但对环境和土壤都可能产生一定的污染,不利于农业的可持续发展,近期在美国还发现对防控灰斑病的QoI杀菌剂产生抗性的灰斑病菌[20]。因此,种植抗病品种是防治大豆灰斑病理想方法,并且依赖于抗病品种选育的速度和水平。由于灰斑病原菌小种构成复杂,优势小种变化快,从大豆生产实践看,仍需在考虑抗病品种的基础上,充分发挥耕作栽培方式、化学防治的作用。
参考文献
[1] YORINORI J T,SINCLAIR J B.Cercosporasojina: A set of differentiated cultivars races for identification[J].Phytopathology,1982,72:173.
[2] 刘忠堂.解决我省东部地区大豆灰斑病的途径[J].黑龙江农业科学,1985(1):20-30.
[3] 廖林.大豆灰斑病研究概况及展望[J].中国农学通报,1992,8(1):6-9.
[4] 曹越平,李海英,刘学敏,等.大豆灰斑病菌(CercosporasojinaHara)及其对寄主作用的研究[J].植物病理学报,2003,33(2):116-120.
[5] 钟兆西,王伟,张桂荣.大豆灰斑病菌(Cercosporasojina)生物学特性的研究[J].大豆科学,1989,8(3):288-294.
[6] KIM H,NEWELL A D,COTA-SIECKMEYER R G,et al.Mating-type distribution and genetic diversity ofCercosporasojinapopulations on soybean from Arkansas:Evidence for potential sexual reproduction[J].Phytopathology,2013,103(10):1045-1051.
[7] 黄桂潮,霍虹,张再兴,等.大豆灰斑病菌(CercosporasojinaHara)生理小种鉴定结果初报[J].大豆科学,1984(3):7.
[8] 霍虹,马淑梅,卢官仲,等.黑龙江省大豆灰斑病菌(CercosporasojinaHara)生理小种的研究[J].大豆科学,1988,7(4):315-320.
[9] 马淑梅,李宝英.东北春大豆灰斑病菌生理小种鉴定结果初报[J].植物病理学报,1997,27(2):180.
[10] 顾鑫,丁俊杰,杨晓贺,等.2008~2009年黑龙江省大豆灰斑病生理小种的监测[J].大豆科学,2010,29(3):540-542.
[11] 马淑梅.2006~2010年黑龙江省大豆灰斑病菌生理小种监测及部分主栽品种抗性鉴定[J].大豆科学,2011,30(3):450-454.
[12] 丁俊杰,顾鑫,杨晓贺,等.黑龙江省大豆灰斑病菌生理小种及遗传关系分析[J].中国农业科学,2012,45(21):4377-4387.
[13] PHAM A T,HARRIS D K,BUCK J,et al.Fine mapping and characterization of candidate genes that control resistance toCercosporasojinaK.Hara in two soybean germplasm accessions[J].PLoS ONE,2015,10(5):126753.
[14] MIAN M A R,MISSAOUI A M,WALKER D R,et al.Frogeye leaf spot of soybean:A review and proposed race designations for isolates ofCercosporasojinaHara[J].Crop science,2008,48(1):14-24.
[15] SCANDIANI M,FERRI M,FERRARI B,et al.First report of races 11 and 12 ofCercosporasojina, the causal agent of soybean frogeye leaf spot,in Argentina[J].Plant disease,2012,96(7):1067.
[16] 张俊华,张明,韩英鹏,等.大豆灰斑病菌生理小种PCR-RFLP分子检测[J].中国油料作物学报,2010,32(1):128-131.
[17] 孙洪利,张俊华,韩英鹏,等.黑龙江省大豆灰斑病菌遗传多样性[J].中国油料作物学报,2011,33(1):57-61.
[18] 丁俊杰,姜翠兰,顾鑫,等.利用与大豆灰斑病抗性基因连锁的SSR标记构建大豆品种(系)的分子身份证[J].作物学报,2012,38(12):2206-2216.
[19] BRADLEY C A,WOOD A,ZHANG G R,et al.Genetic diversity ofCercosporasojinarevealed by amplified fragment length polymorphism markers[J].Canadian journal of plant pathology,2012,34(3):410-416.
[20] ZHANG G R,NEWMAN M A,BRADLEY C A.First report of the soybean frogeye leaf spot fungus (Cercosporasojina) resistant to quinone outside inhibitor fungicides in North America[J].Plant disease,2012,96(5):767-767.
[21] ZENG F,ARNAO E,ZHANG G,et al.Characterization of quinone outside inhibitor fungicide resistance inCercosporasojinaand development of diagnostic tools for its identification[J].Plant disease,2015,99(4):544-550.
[22] 刘学敏,李本宁.大豆灰斑病流行动态预测[J].大豆科学,1996,15(3):222-227.
[23] 丁俊杰.三江平原地区降水量变化与大豆灰斑病相关性分析[J].东北农业大学学报,2013,44(7):1-5.
[24] 刘惕若,李海燕,甄鸿杰.大豆灰斑病流行强度预测模型研究[J].中国油料作物学报,2006,27(3):54-57.
[25] 顾鑫,杨国珍,丁俊杰,等.黑龙江省东部地区大豆灰斑病短期预测模型的建立[J].黑龙江农业科学,2014(12):67-69.
[26] MENGISTU A,KELLY H M,BELLALOUI N,et al.Tillage,fungicide,and cultivar effects on frogeye leaf spot severity and yield in soybean[J].Plant disease,2014,98(11):1476-1484.
[27] 方中达.中国农业植物病害[M].北京:中国农业出版社,1996.
[28] 曹越平,杨庆凯.大豆灰斑病抗感标准划分的研究[J].大豆科学,2002,21(2):113-116.
[29] SHARMA H,LIGHTFOOT D A.Quantitative trait loci underlying partial resistance toCercosporasojinarace 2 detected in soybean seedlings in green-house assays[J].Atlas journal of biology,2014,3(1):175-182.
[30] 陈庆山.大豆灰斑病种质资源遗传多样性的RAPD和SSR分析[D].哈尔滨:东北农业大学,2001.
[31] 万学臣.大豆灰斑病抗源筛选及其与若干性状的关系[J].作物品种资源,1987(2):9.
[32] 姚振纯,张玉华.野生大豆田间感染大豆灰斑病简报[J].大豆科学,1986,5(4):349-350.
[33] 张文慧,杜吉到,陈庆山,等.抗大豆灰斑病育种研究进展[J].黑龙江八一农垦大学学报,2006,18(1):22-26.
[34] 马淑梅,韩新华,邵红涛.大豆主要病害多抗性资源筛选鉴定[J].中国农学通报,2014(27):12.
[35] MENGISTU A,BOND J,MIAN R,et al.Resistance to frogeye leafspot in selected soybean accessions in MG I through MG VI[J].Plant health progress,2012,10.doi:10.1094/PHP-2012-0521-02-RS.
[36] YANG W,WEAVER D B.Resistance to frogeye leaf spot in maturity groups VI and VII of soybean germplasm[J].Crop science,2001,41(2):549-552.
[37] 朱希敏,王利财,邹桂珍.大豆品种资源抗大豆花叶病 (SMV),灰斑病 (Cercosporasojina) 和霜霉病 (Peronosporamanschurica) 的鉴定和评价[J].大豆科学,1988(3):9.
[38] 张丽娟,杨庆凯.大豆抗灰斑病菌多个生理小种资源的筛选[J].大豆科学,1997,16(1):38-41.
[39] 吴秀红.大豆抗灰斑病菌多个生理小种资源的筛选[J].植物遗传资源学报,2003,4(4):341-344.
[40] 马淑梅,李宝英.黑龙江省大豆品种和资源与本省大豆斑病菌相互作用的研究[J].大豆科学,1996,15(4):322-325.
[41] 马淑梅.大豆种质资源对灰斑病抗性鉴定评价[J].植物遗传资源学报,2011,12(5):820-824.
[42]马淑梅,韩新华.大豆品种资源对灰斑病的抗性鉴定[J].大豆科学,2014(6):957-960.
[43]马淑梅,韩新华,邵红涛.大豆主要病害多抗性资源筛选鉴定[J].中国农学通报,2014,30(27):58-65.
[44]丁俊杰,顾鑫,杨晓贺,等.黑龙江省大豆新品系抗灰斑病鉴定[J].东北农业大学学报,2011,42(10):120-124.
[45]CHAWLAS,BOWENCR,SLAMINKOTL,etal.Apublicprogramtoevaluatecommercialsoybeancultivarsforpathogenandpestresistance[J].Plantdisease,2013,97(5):568-578.
[46]刘忠堂.抗灰斑病大豆育种技术的探讨[J].大豆科学,1986,5(2):147-152.
[47]齐宁,郭泰,刘忠堂,等.高抗灰斑病高产大豆新品种合丰34号的选育[J].中国油料,1994,16(4):65-66.
[48]郭泰,刘忠堂,齐宁,等.大豆高抗灰斑病品种合丰29号的选育及利用[J].作物品种资源,1997(3):25-26.
[49]刘成贵,刘秀芝,郑伟,等.抗灰斑病高油高产大豆新品种合农64选育与推广[J].黑龙江农业科学,2015(3):1-4.
[50]刘秀芝,刘成贵,王志新,等.早熟高产优质抗病大豆新品种合农59选育与推广[J].黑龙江农业科学,2015(2):1-5.
[51]顾鑫,丁俊杰.大豆灰斑病的研究现状[J].中国农学通报,2010,26(9):303-306.
[52]邹继军,杨庆凯.大豆灰斑病抗病基因RAPD标记的分子特征及抗,感种质的SCAR标记鉴定[J].科学通报,1999,44(23):2544-2550.
[53]张文慧,陈庆山,杨庆凯,等.大豆灰斑病1号生理小种抗性基因的SSR标记分析[J].大豆科学,2004,23(3):169-173.
[54]陈立君,郭强,刘迎雪,等.大豆灰斑病1号生理小种抗性基因的SSR标记[J].中国农学通报,2009,25(9):43-46.
[55]DONGZ,WANGS,LIUJ,etal.PreliminarymappingofsoybeandominantlocusHrcs7conferringresistancetoCercospora sojinarace7[J].Molecularplantbreeding,2011,2(6):37-40.
[56]姜翠兰,丁俊杰,文景芝,等.大豆对灰斑病菌 15 号小种的抗病基因定位及标记检测[J].植物保护学报,2011,38(2):116-120.
[57]MIANMA,WANGT,PHILLIPSDV.MolecularmappingoftheRcs3geneforresistancetofrogeyeleafspotinsoybean[J].Cropscience,1999,39(6):1687-1691.
[58]MISSAOUIAM,HABK,PHILLIPSDV,etal.SinglenucleotidepolymorphismdetectionoftheRcs3geneforresistancetofrogeyeleafspotinsoybean[J].Cropscience,2007,47(4):1681-1690.
中图分类号S435.651
文献标识码A
文章编号0517-6611(2015)30-22-04
基金项目国家高技术研究发展计划(863计划)项目(2012AA101106);长江学者和创新团队发展计划项目(PCSIRT13073);江苏省现代作物生产协同创新中心项目(JCIC-MCP)。
作者简介程伟(1988-),男,安徽亳州人,硕士研究生,研究方向:大豆遗传育种。*通讯作者,助教,硕士,从事大豆遗传育种研究。
收稿日期2015-09-10
Advance in the Disease Occurrence and Resistance Breeding of Frogeye Leaf Spot in Soybean
CHENG Wei, CHANG Fang-guo, ZHAO Tuan-jie,KONG Jie-jie*(Soybean Research Institute of Nanjing Agricultural University/National Center for Soybean Improvement/ Key Laboratory of Soybean Biology and Genetic Improvement, Ministry of Agriculture/National Key Laboratory for Crop Genetics and Germplasm Enhancement, Nanjing, Jiangsu 210095)
AbstractFrom the viewpoint of resistance breeding, some aspects including the pathogen’s characteristics, its physiological race differential and classification, the disease epidemiology and its control, identification methods for resistance and screening of elite resistant germplasm (genes), genetic improvement for resistance to frogeye leaf spot were reviewed. Some discussions involved. It is necessary to strengthen the research on the mechanism of disease occurrence, race identification system, discovery of elite resistant lines (genes) and combination of convention and molecular breeding for resistance to frogeye leaf spot.
Key wordsSoybean; Frogeye leaf spot; Race differentiation; Resistance breeding; Molecular marker
