西藏色季拉山两种忍冬属植物点格局分析
2015-12-20金增涛解传奇次仁卓嘎郑维列王国严扈晶晶
金增涛,解传奇,次仁卓嘎,罗 建,郑维列,王国严,扈晶晶
(1.西藏自治区林业调查规划研究院,拉萨 850000; 2.西藏大学农牧学院,西藏 林芝 860000)
西藏色季拉山两种忍冬属植物点格局分析
金增涛1,解传奇2,次仁卓嘎2,罗 建2,郑维列2,王国严2,扈晶晶1
(1.西藏自治区林业调查规划研究院,拉萨 850000; 2.西藏大学农牧学院,西藏 林芝 860000)
基于色季拉山长期4 hm2监测样地的调查资料,采用成对相关函数(pair-correlation function)点格局方法,分析了色季拉山观测样地灌木层优势种杯萼忍冬和柳叶忍冬的空间格局及关联性。结果表明:发现两种忍冬都在小尺度上呈显著聚集分布,随着龄级和研究尺度的增加种群聚集强度逐渐减弱,最终趋向于随机分布格局;杯萼忍冬种群和柳叶忍冬种群在小尺度上具有正的关联性。
杯萼忍冬;柳叶忍冬;空间格局;种间关联
物种的空间格局及其形成机制是生态学研究的核心内容,植物种群空间格局的研究一直是生态学中的研究热点[1-4]。植物种群空间格局的研究在揭示森林结构、动态及其生态学过程、种内和种间关系以及种群与环境关系、正确评价、合理经营森林群落等方面有着重要意义[5-6]。
西藏亚高山原始暗针叶林的典型代表是急尖长苞冷杉(Abiesgeorgeivar.smithii)林,群落内乔、灌、草层次分明,群落郁闭度较高(0.6~0.8)[7]。杯萼忍冬和柳叶忍冬是色季拉山冷杉林群落灌木层的主要优势物种,是色季拉山区植被垂直带谱中的重要组成部分。杯萼忍冬(Lonicerainconspicua)是忍冬科忍冬属植物,落叶灌木,树高1~3 m,为中国特有。主要分布于甘肃南部、四川西部、西藏东南部,海拔1700~3400 m的山坡或谷地、沟边的林下、林缘和灌丛等处。柳叶忍冬(L.lanceolata)为忍冬科忍冬属植物,落叶灌木,高达4 m。主要分布于尼泊尔、不丹以及中国大陆的四川、西藏、云南等地,海拔2000~3900 m的林缘或灌丛中。
点格局分析法最大限度地利用了点与点之间的距离,能够提供较为全面的空间尺度信息[8]。因此,它可以分析任意尺度上种群空间格局和种间关系,同时还能够给出空间格局的最大聚集强度及其对应的尺度,为群落内种群空间格局的比较提供了方便[4-9]。近年来成对相关函数(pair-correlationfunction)因能避免尺度累积效应而被广泛应用于分析植物种群多尺度空间分布格局和种群间空间关联性[10-11]。
1 研究地概况及研究方法
1.1 研究地自然概况
色季拉山位于西藏东南部雅鲁藏布江大峡谷西北侧,系念青唐古拉山向南延伸的余脉,与喜马拉雅山向东发展的山系相连,山脉走向大致西北至东南,主峰海拔高度5200 m。属于湿润山地暖温带和半湿润山地温带气候,冬温夏凉、干湿季分明。土壤为漂灰化山地暗棕壤,土层平均厚度60 cm,腐殖质化程度明显[12-13]。主要植被类型有高山稀疏垫状植被、以云冷杉为主的暗针叶林、以川滇高山栎为主的常绿硬阔叶林和以杨桦林为主的落叶阔叶林等。试验地位于西藏林芝地区森林生态系统色季拉山长期定位站对面大阳坡(29°41′N,94°23′E),海拔3600~3800 m,样地内主要冠层植被是以急尖长苞冷杉为主的暗针叶林,并伴有少量红毛樱(Prunusserrula)、西南花楸(Sorbusrehderiana)、褐背柳(Salixdaltoniana)等低矮小乔木;林下灌木层主要优势种有白蓓紫斑杜鹃(Rhodendronvellereum)、杯萼忍冬(Lonicerainconspicua)、柳叶忍冬(Loniceralanceolata)、峨眉蔷薇(Rosaomeiensis)、冰川茶藨子(Ribesglaciale)等。
1.2 研究方法
1.2.1 样地设置与调查
2013年7月在试验区设定4 hm2固定监测样地一个,样地南高北低,呈折页状,水平距离南北长200 m,东西长200 m,海拔3790~3875 m。用全站仪 (拓普康GTS—102N)选定样地中线,然后由中线向两侧分散布置,将样地划分为100个20 m×20 m 的小样方。于2013年7—8月对样地内植物群落进行全面调查,对所有主胸径≥1 cm的乔、灌木进行编号并挂牌标记,并识别到种,测量每株植物坐标、胸径、分枝数并记录样方号、种名、生境特征等数据。
1.2.2 数据分析方法
点格局分析方法中的成对相关函数(pair-correlationfunction)包括单变量的g(r)分析和双变量的g12(r)分析。本文采用单变量的g(r)函数分析不同种群的空间格局,用双变量的g12(r)函数分析两种忍冬间的关联性。
为获得有效分析结果,数据分析应仔细筛选零假设模型[14-15],在本次研究中,对于单变量统计,使用异质性Poisson过程零假设,以考虑可能的环境异质性的存在。对于双变量统计,由于种群之间的竞争是非对称性的,采用完全随机过程零假设[16]。空间格局和空间关联性分析统一采用0~100 m的研究尺度,并经过99次Monte Carlo模拟得到95%的置信区间。整个数据分析过程使用Programita软件(Wiegand,2008版)完成。
2 结果与分析
2.1 空间格局
经实地调查,样地内共有杯萼忍冬2172株,柳叶忍冬1852株,占样地内所有灌木总个体数的63.1%,为群落内灌木层的主要优势种。由图1可以看出,两种忍冬在样地内分布较为均匀,并没有明显的地域差异性。
对杯萼忍冬种群进行g(r)单变量统计分析发现(图2),整个种群在0~3 m尺度内呈显著聚集分布,在3~100 m尺度上为随机分布;由图3可以看出,柳叶忍冬在0~4 m尺度上呈聚集分布格局,在4~100 m尺度上为随机分布格局。说明两种忍冬都在空间小尺度上呈聚集分布,而随着空间尺度的增大,聚集度逐渐减低,表现出两个种群在种内资源利用方式和空间分布格局上的相似性。而大尺度的随机分布则可能与大环境的空间异质性有关。
2.2 空间关联性
对色季拉山杯萼忍冬种群和柳叶忍冬种群进行g12(r)分析发现(图4):杯萼忍冬和柳叶忍冬在0~4 m尺度内呈正的关联性,随着空间尺度的增大关联性逐渐减弱,逐渐表现为无关联。验证了两个种群各自种内空间分布格局的相近性,说明两个亲缘关系较近的同属植物不仅在形态结构上具有相似性,其生活习性、种子散布等生活史特征以及对环境的适应方式都具有相似性。
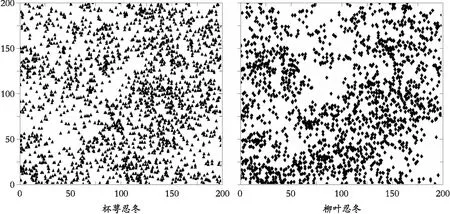
图1 样地内两种忍冬分布图
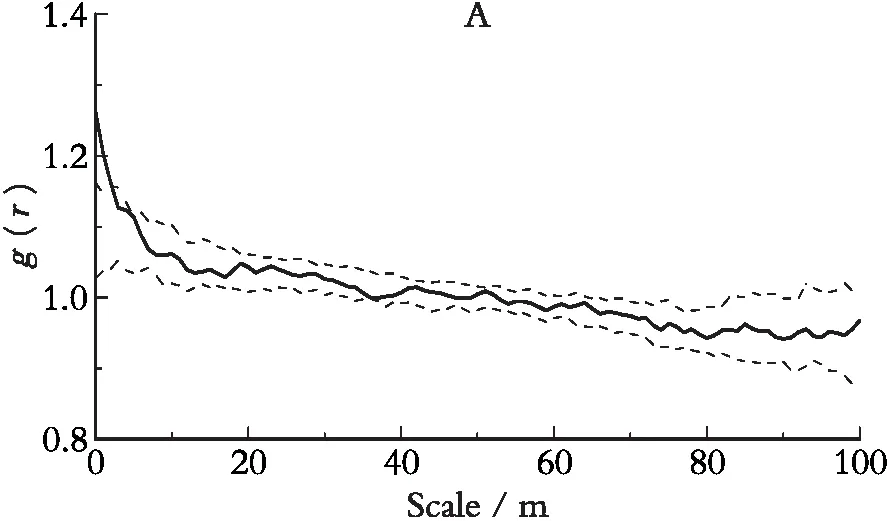
图2 色季拉山4 hm2样地杯萼忍冬空间分布格局
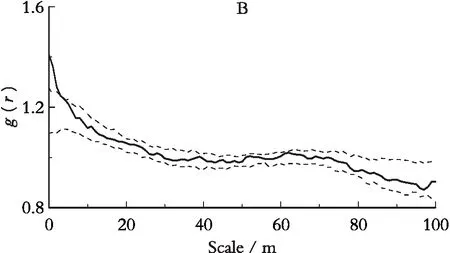
图3 色季拉山4 hm2样地柳叶忍冬空间分布格局
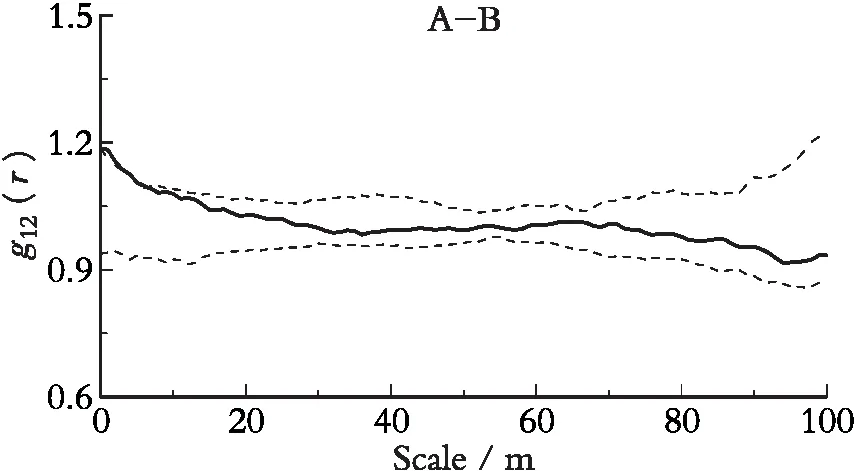
图4 杯萼忍冬与柳叶忍冬间的空间关联性
3 讨论
研究尺度是影响种群空间分布格局的重要因素,因此对格局的识别往往是从尺度辩识开始的[17]。种群分布格局具有尺度依赖性,不同尺度下往往呈现不同的分布格局。本研究表明两种忍冬都在小尺度上呈聚集分布格局,大尺度上呈随机分布格局,这与张金屯等[18]的研究一致。而造成这种分布格局的原因是多方面的,一是个体以集群形式占据和利用资源,有利于维持种群自身的稳定;二是种子扩散也会影响种群分布格局,种子散布的限制会导致种子萌发不均匀和幼苗分布的不均匀,最终表现为聚集型分布格局;三是生境异质性也是促成种群聚集格局的重要因素,种子萌发和幼苗生长受地形、光斑、林隙、土壤有机质等的影响很大[19-21]。
种群之间的空间关联性体现的是物种之间的竞争关系或互惠关系以及不同物种对生境分化的响应。空间关联性通常是由群落生境的差异影响物种分布引起的[22]。大量研究表明物种间的关联性也受空间尺度的影响,两个物种的关联关系在一种尺度下可能是正相关,而在另一种尺度下则可能是负关联或者没有显著的相关性[23]。本研究表明杯萼忍冬和柳叶忍冬在小尺度内呈正的关联性,随着空间尺度的增大关联性逐渐减弱至无关联。有研究认为资源限制导致在生态学尺度上相似的近缘种通常不可能共存,种间竞争在这类物种间变得更加强烈,尤其在小尺度上[24]。
但在本研究中同属种杯萼忍冬与柳叶忍冬却在小尺度上正关联,这可能是由两方面的原因造成的。首先,由于该群落内近缘种之间生态位重叠水平较低,在群落内利用相同资源或占有同一资源而产生的种间竞争并不激烈,对资源的共享比较充分;其次,由于近缘种间长期的竞争和适应,近缘种生态位趋于特化[25]。
综上所述,本研究利用点格局方法分析了色季拉山4 hm2永久观测样地灌木层优势种杯萼忍冬和柳叶忍冬的空间格局及关联性。发现两种忍冬都在小尺度上呈显著聚集分布,随着研究尺度的增加种群聚集强度逐渐减弱,最终趋向于随机分布格局;杯萼忍冬种群和柳叶忍冬种群在小尺度上具有正的关联性。物种的空间格局及其相关性分析仅仅是探讨森林格局与生态过程之间关系、物种多样性维持机制的第一步,其复杂性需要继续深入研究。
[1] Pielou EC. An introduction to mathematical ecology[M]. NewYork: Wiley Interscience Publication,1969.
[2] Greig-Smith P. Quantitative plant ecology[M]. Oxford: Blackwell,1983:54-104.
[3] Dale,MRT. Spatial Pattern Analysis in Plant Ecology[M]. London: Cambridge university press,1999.
[4] Wiegand T,Moloney KA. Rings,circles,and null-models for point pattern analysis in ecology[J]. Oikos,2004,104: 209-229.
[5] Wiegand T,Gunatilleke S,Gunatilleke N. Species associations in a heterogeneous Sri Lankan dipterocarp forest[J]. The American Naturalist,2007,170: 67-95.
[6] Getzin S,Wiegand T,Wiegand K,etal. Heterogeneity influences spatial patterns and demographics in forest stands[J]. J Ecol,2008,96: 807-820.
[7] 任德智,罗大庆. 基于GIS的林隙特征测算方法——以西藏林芝地区色季拉山急尖长苞冷杉原始林为例[J]. 东北林业大学学报,2013,41(1): 17-20.
[8] Wang YF,J. Julio C,Luo TX,etal. Spatial patterns of Smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns[J]. Plant Ecology & Diversity,2012,311-321.
[9] Stoyan D,Stoyan H. Fractals,random shapes and point fields methods of geometrical statistics[J]. John Wiley&Sons,1994.
[10] Condit R,Ashton PS,Baker P,etal. Spatial patterns in the distribution of tropical tree species[J]. Science,2000,288: 1414-1418.
[11] Zhu Y,Mi XC,Ren HB,etal. Density dependence is prevalent in a heterogeneous subtropical forest[J]. Oikos,2010,119: 109-119.
[12] 孙鸿烈,徐阿生,罗大庆,等. 森林生态系统卷:西藏林芝卷2001—2007[M]. 中国农业出版社,2010.
[13] 王国严,罗建,徐阿生. 西藏色季拉山冷杉林下优势种群种间关联[J]. 东北林业大学学报,2009,37(3): 21-23.
[14] 张健,郝占庆,宋波,等. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J]. 应用生态学报,2007,18(8): 1681-1687.
[15] 闫海冰,韩有志,杨秀清,等. 华北山地典型天然次生林群落的树种空间分布格局及其关联性[J]. 生态学报,2010,30(9): 2311-2321.
[16] 张金屯. 数量生态学[M]. 北京: 科学出版社,2004.
[17]刘振国,李镇清.不同放牧强度下冷蒿种群小尺度空间格局[J]. 生态学报,2004,24(2):1020-1028.
[18] 张金屯,孟东平. 卢芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报,2004,24(1): 35-40.
[19] 张毓涛,李吉玫,常顺利,等. 天山中部天山云杉种群空间分布格局及其与地形因子的关系[J]. 应用生态学报,2011,2(11): 2799-2806.
[20] 韩有志,王政权,谷加存. 林分光照空间异质性对水曲柳更新的影响[J]. 植物生态学报,2004,28(4): 468-475.
[21] 杨秀清,韩有志,李乐,等. 华北山地典型天然次生林土壤氮素空间异质性对落叶松幼苗更新的影响[J]. 生态学报,2009,29(9): 656-664.
[22] 奇凯. 黑里河天然油松林主要植物种空间格局和空间关联[D]. 北京林业大学,2011.
[23] 缪宁,刘世荣,史作民,等. 青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局[J]. 生态学报,2011,31(1): 0001-0009.
[24] Webb C O,Ackerly,D D,McPeek,M A,Donoghue,M J. Phylogenies and community ecology[J]. Annual Review of Ecology and Systematics,2002,33: 475-505.
[25] 胡正华,钱海源,于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报,2009,29(7):3670-3677.
PointPatternAnalysisofTwoLoniceraLinn.SpeciesinSegeraMountainsofTibet
JIN Zengtao1,XIE Chuanqi2,Ciren Zhuoga2,LUO Jian2,ZHENG Weilie2,WANG Guoyan2,HU Jingjing1
(1. Forest Inventory and Planning Institute of Tibet Autonomous Region,Lhasa 850000,Tibet,China; 2.Tibet Agriculture and Animal Husbandry College,Linzhi 860000,Tibet,China)
In this study,based on the investigating materials of a 4 hm2stem-mapping plot,we analyzed spatial distributions ofLonicerainconspicuaandL.lanceolate,the dominant species in forest of Segera mountains. Pair-correlation function was used to characterize spatial point patterns of the population and spatial realations between these two species. TwoLoniceraLinn. Species of the population were spatially obviously aggregated at the small scales(0~4 m),and the distribution were more and more random and uniform with the increases of the age and research scale,finally tending to random distribution pattern.LonicerainconspicuaandL.lanceolatewere positive related at the small scales.
Lonicerainconspicua;L.lanceolate;spatial pattern;interspecific association
2015—08—11
2015—08—27
国家自然科学基金项目(31260147);西藏林芝高山森林生态系统定位研究项目。
金增涛(1980—),男,汉族,山东临清人,硕士,工程师,主要从事林业调查、规划方面的工作。
S 718.54+2
A
1003—6075(2015)03—0043—04
10.16166/j.cnki.cn43—1095.2015.03.010
