松材线虫侵害马尾松林后群落谱系多样性和结构动态
2015-12-18王玉玲刘金亮徐学红金毅陆高
王玉玲,刘金亮,徐学红,金毅,陆高
(1.国家林业局森林病虫害防治总站,辽宁 沈阳 110034;2.浙江大学生命科学学院,浙江 杭州 310058;3.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;4.象山县森林病虫防治检疫站,浙江 象山 315700)
物种维持机制一直以来是群落生态学和保护生物学探讨的核心问题之一[1-3]。群落内现有物种组成是长期的进化和生态过程共同作用的结果,随着人类活动影响的加剧,探讨不同干扰方式对群落谱系结构影响的研究日益增多,人为干扰如择伐[4]、自然干扰如冰雪灾害等[5-6]。同时,生物入侵作为一种对本地群落的干扰方式,会影响原有群落内的物种多样性、改变群落内物种的组成以及群落空间结构的变化[7-8]。松材线虫病是由松材线虫Bursaphelenchus xylophilus寄生于松树体内导致松树迅速死亡的一种病害[9]。自1982年在我国南京市中山陵首次发现松材线虫病以来[10],由于入侵种松材线虫缺乏天敌制约等因素的影响,1982—2002年共有约9万hm2的松林被其感染致病[11]。松材线虫侵害马尾松林后,会使群落内松树感病死亡,影响群落内物种组成和演替方向[7,12-13]。本文主要探讨松材线虫侵害马尾松林后群落谱系多样性和结构的动态,旨在更深入地了解松材线虫入侵对本地生态系统的影响,为侵害迹地的生态恢复提供科学依据。
1 研究地概况
浙江省象山县地区(E 121°32'~121°12',N29°14'~29°38')属亚热带季风性湿润气候,年平均气温16.5℃,无霜期248 d,年日照时数1670~2048 h,年均降水量1450 mm[7]。1991年首次记录松材线虫入侵以来,该地区具有很高的松材线虫感染率[7]。
2 研究方法
2.1 样地设置和物种调查根据象山县林业局自1991年以来松材线虫发生情况的统计材料,2003年在象山县内选择海拔高度、土壤、坡向、坡度、坡位等生境相似,经历4个不同发病时间的林地作为研究样地,分别为当年(记为0 a)发病的泗洲头镇、发病4 a的大徐镇、发病8 a的墙头镇、发病12 a的丹城镇,样地内的森林群落在感染松材线虫前均为马尾松林,林龄20~30 a。在象山县附近的天童森林公园选择林龄约60 a未被松材线虫侵害的常绿阔叶林中龄林作为对照林地(图1)[14]。
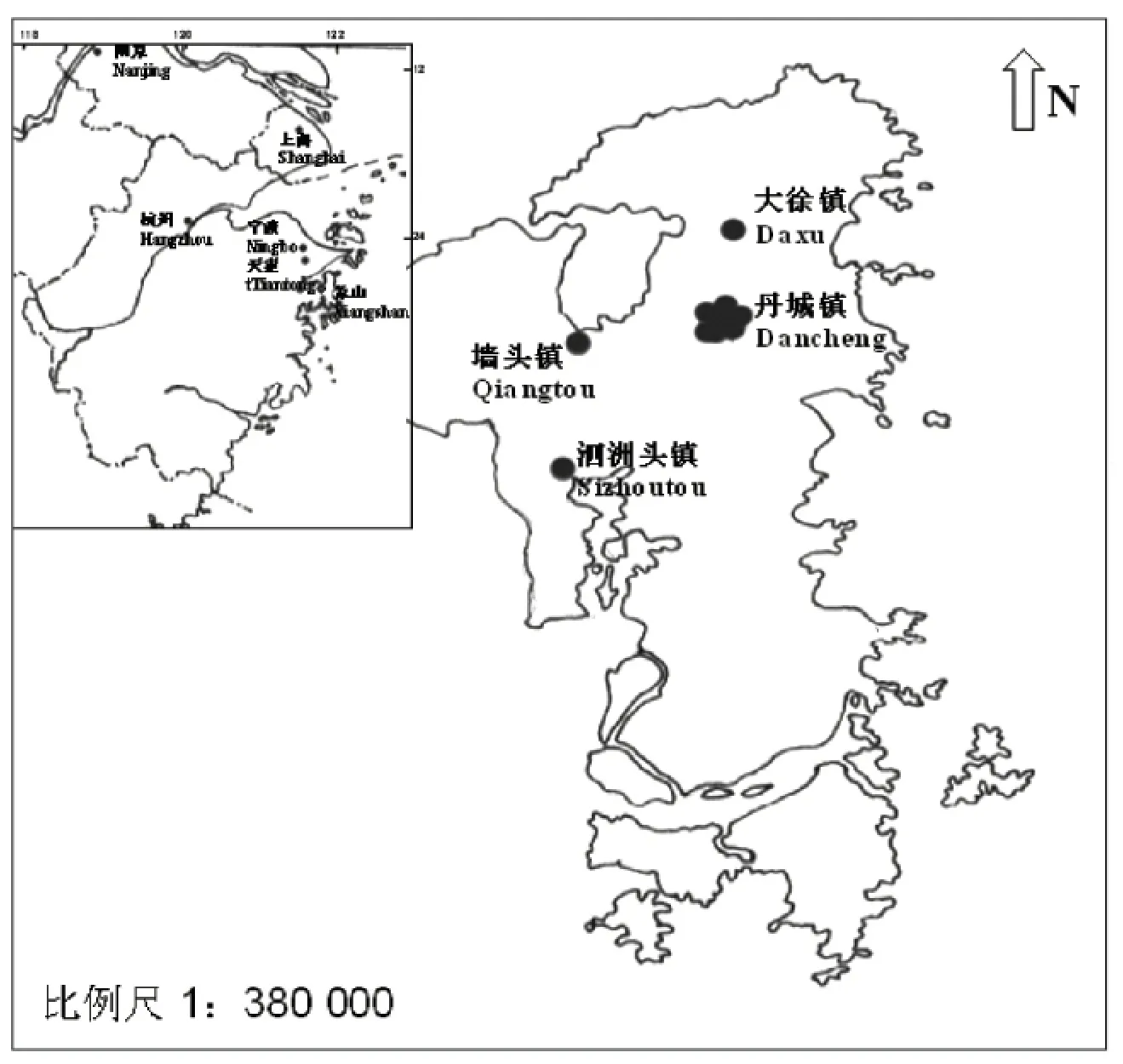
图1 松材线虫侵染不同时间的样地和对照样地分布图
每个样地选取16个10 m×10 m的样方,样方之间至少间隔20 m。每个样方划分为4个5 m×5 m的小样方,对小样方内乔木树种进行每木调查。树高大于1.3 m的记录其胸径、基径、树高、冠幅等,树高小于1.3 m的记录基径、树高、冠幅等;灌木和草本记录其高度、盖度和多度。
2.2 谱系树构建基于Qian和Zhang已发表的谱系树[15],使用Phylomatic V3软件[16]对样方内调查到的148种乔木、灌木以及草本植物(不含蕨类植物和藤本植物)构建谱系树。其中,豆科的小槐花Ohwia caudata和宽卵叶山蚂蝗Podocarpium podocarpum var.fallax无法匹配,均出现在泗洲头样地,个体数均为1,故没放入之后的谱系分析中。
2.3 谱系指数选择利用净谱系亲缘关系指数(Net relatedness index,NRI)和净最近种间亲缘关系指数(Nearest taxon index,NTI)来分析群落的谱系结构[17]。NRI主要从整体上反映群落中物种形成的谱系结构,而NTI主要反映群落内亲缘关系较近物种的谱系结构。假定由已调查的物种组成局域物种库,首先计算出样方中所有物种两两间的平均谱系距离(Mean phylogenetic distance,MPD)和最近种间平均进化距离(Mean nearest taxon distance,MNTD),保持物种数量及物种个体数不变,从物种库中随机抽取样方中物种的物种名999次,获得该样方中物种在随机零模型下MPD和MNTD的分布,之后利用随机分布结果将观察值标准化,从而获得不受物种数影响的NRI和NTI值。计算公式为[17-18]:

其中,n为群落内物种数,δij为物种i和物种j之间的谱系距离,minδij为物种i与群落内其他物种间的最短谱系距离,MPDs和MNTDs代表观察值,MPDr和MNTDr代表物种在谱系树上通过随机后获得的平均值。若NRI>0,NTI>0,样方中的物种在谱系结构上聚集;若NRI<0,NTI<0,样方中的物种在谱系结构上发散;若NRI=0,NTI=0,样方中的物种在谱系结构上是随机的。
所有计算均采用R软件[19]。利用R软件中的Picante包[20]计算谱系多样性和NRI、NTI指数,Vegan包[21]计算群落间物种相似性指数(Sorensen指数)。
3 结果
3.1 群落内物种组成和谱系多样性对于物种多样性而言,松材线虫侵害0 a的泗洲头的物种96种,丰富度最高;所有样地乔木层物种数均高于灌木层和草本层物种数(表1)。对于谱系多样性而言,松材线虫侵害0 a的群落具有最高的谱系多样性,随着侵害时间的增加,谱系多样性降低,且与对照常绿阔叶林接近(图2)。对物种组成而言,松材线虫侵害0 a与侵害4,8,12 a群落内的物种和谱系相似性低于侵害4,8,12 a之间的相似性。对照常绿阔叶林样地与侵害4,8 a群落内的物种和谱系相似性最高,与侵害0 a的相似性最低(表2)。
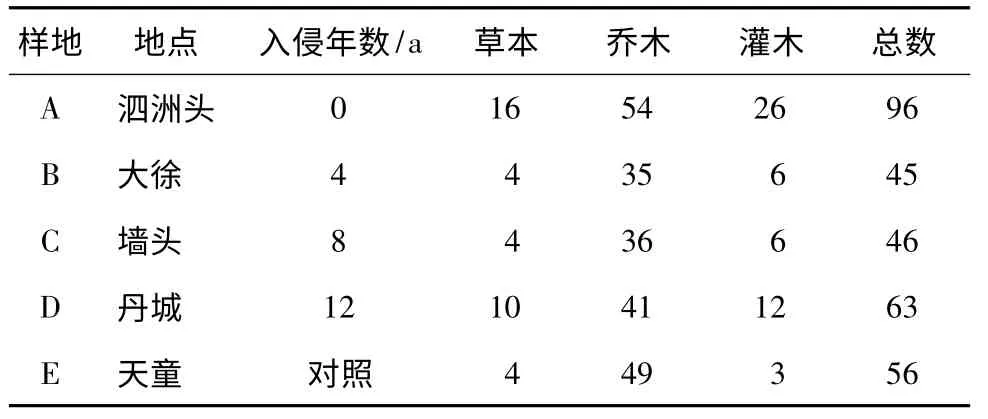
表1 松材线虫侵害不同时间的马尾松林及对照常绿阔叶林群落内的物种数
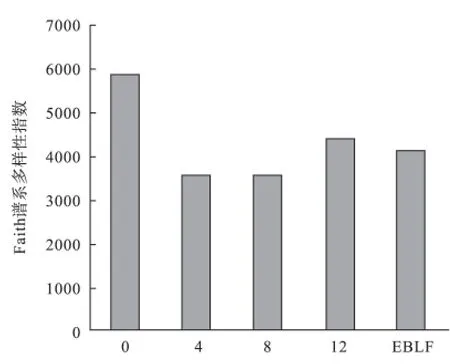
图2 松材线虫侵害马尾松林和对照常绿阔叶林的群落Faith谱系多样性
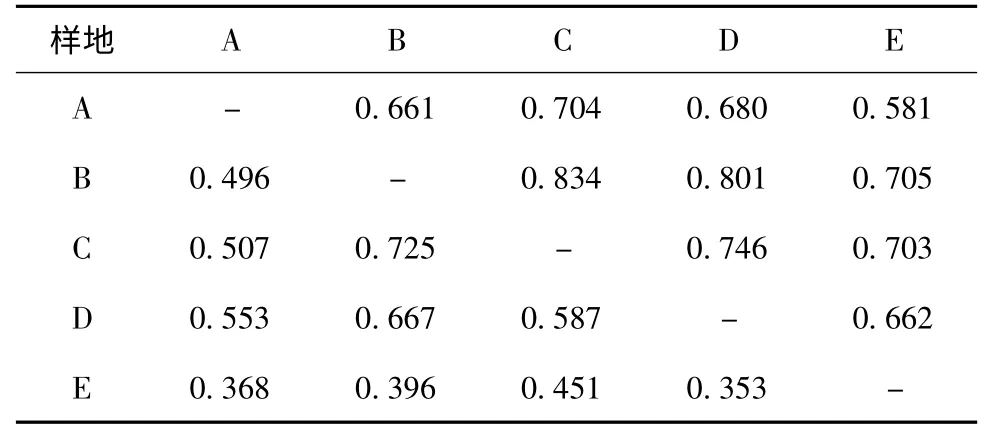
表2 不同样地间的物种(下三角)和谱系(上三角)相似性指数
3.2 群落的谱系结构松材线虫侵害马尾松林0 a和12 a群落内的MPD和MNTD值均低于4 a和8 a的,且受害4 a和8 a的MPD和MNTD值达到最大,受害12 a后群落内的MPD和MNTD值与常绿阔叶林接近(图3)。
松材线虫侵害马尾松林0,4 a群落NRI、NTI均小于0,谱系结构发散,且随侵害时间增加,NRI与NTI值越小;松材线虫侵害马尾松林12 a的与常绿阔叶林的群落NRI、NTI均大于0,谱系结构聚集(图4)。
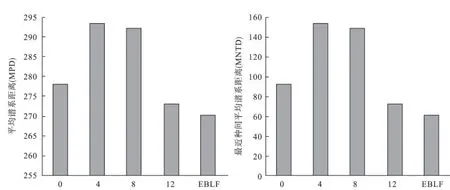
图3 样地和对照地群落内所有物种的平均谱系距离(MPD)和最近种间平均谱系距离(MNTD)
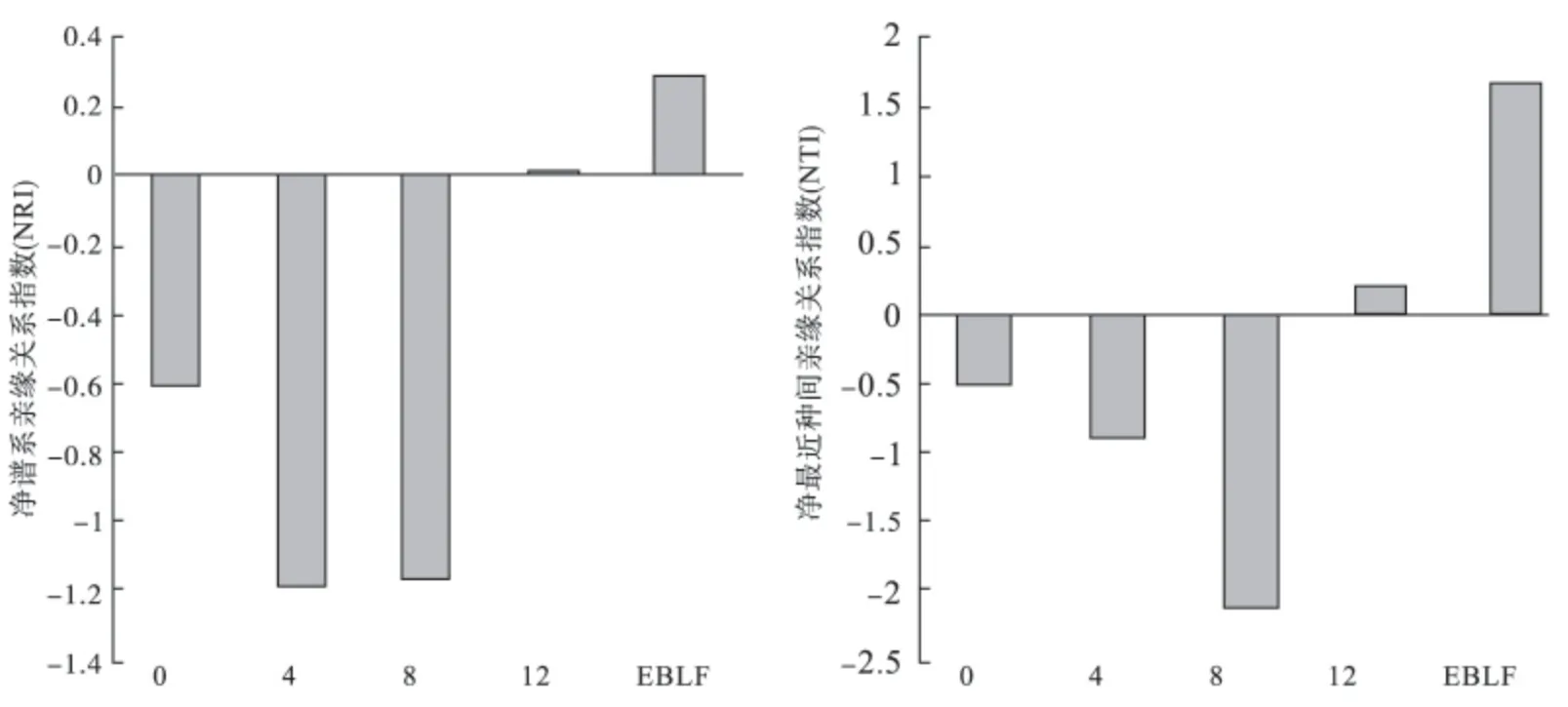
图4 样地和对照地群落内所有物种的净谱系亲缘关系指数(NRI)和净最近种间亲缘关系指数(NTI)
4 讨论
以往研究发现,入侵生物一般会降低本地种的生物多样性,然而入侵种对于生物多样性的影响同时也受到时间、干扰等外界因素的制约。随着时间的推移,入侵态势逐渐趋于稳定,有利于本地生态系统的恢复[8]。与其它生物入侵现象不同的是,松材线虫侵害后导致群落内马尾松等松树死亡,造成物种多样性降低,但会加速森林群落向常绿阔叶林的演替[7,12]。本研究中,松材线虫侵害马尾松林当年(0 a)的群落具有最高的物种和谱系多样性,这是因为该马尾松群落内含有大量与马尾松伴生的先锋物种及喜光物种,如覆盆子Rubus coreanus、美丽胡枝子Lespedeza formosa、三脉紫菀Aster ageratoides等。随着松材线虫侵害时间的增加,会促使群落内主要优势种马尾松以及喜阳物种死亡,造成群落内的物种多样性和谱系多样性均显著下降。但在侵害时间达12 a时,由于马尾松等先锋物种已全部消失,具有耐阴作用的常绿阔叶林物种开始占主要优势,使得物种和谱系多样性水平与常绿阔叶林相近。此外,松材线虫侵害后,马尾松死亡造成群落内环境条件改变,对物种进行筛选,使得群落的物种和谱系组成的相似性增加(如侵害4 a与8 a后的群落具有很高的相似性),且常绿阔叶林样地与松材线虫侵害4,8 a后的群落内物种和谱系相似性最高,与侵害0 a的相似性最低,说明松材线虫侵害后群落的物种组成逐渐向常绿阔叶林过渡。
松材线虫侵害12 a后,群落内的谱系距离和谱系结构均与常绿阔叶林一致,是因为在松材线虫侵害前期,马尾松林尚处于演替前期的早期阶段,此时群落内生境同质性强,且幼苗的新增主要依靠种子传播,使得物种间竞争作用强烈,谱系结构发散[5];但松材线虫侵害后,导致马尾松死亡,而阔叶树并不受其害,随着马尾松死亡时间的延长,在群落内形成大量的林窗,释放出了大量的资源,为阔叶树种新个体的定植和生长提供更多的生态位,此时生境异质性增加,环境过滤作用加剧,形成与常绿阔叶林一致的谱系聚集结构。有研究表明,生态位过程在亚热带常绿阔叶林中群落的维持起重要作用,一般表现为谱系聚集[22]。该结果也能说明,松材线虫侵害后,能够促使群落的谱系结构由谱系发散向谱系聚集转变,促使演替前期的马尾松林向常绿阔叶林转变。
从物种进化历史的角度,能够加深对物种共存机制和群落演替的理解。马尾松林作为中国亚热带地区主要的经济林,在松材线虫侵害后,将使群落的物种组成和谱系结构向稳定的常绿阔叶林转变。对于常绿阔叶林而言,能够提供比马尾松林更多的生态服务,如丰富的动植物多样性[23]、更高的初级生产力和碳储存等[23-24]。虽然马尾松林被松材线虫侵害后短期内会造成木材减产和经济损失,但从生态系统功能和物种保护的角度来看,松材线虫入侵反而具有积极的作用[7]。
因此,关于松材线虫的防治工作,应结合其对群落的长期影响,权衡短期经济损失和长期生态价值的获得,针对具体情况做出相应的管理策略[7]。
志谢:浙江师范大学的南歌在物种名录整理过程中提供了大量帮助,在此表示谢意。
[1] 侯继华,马克平.植物群落物种共存机制的研究进展[J].植物生态学报,2002,26(z1):1-8.
[2] Vellend M.Conceptual synthesis in community ecology[J].The Quarterly Review of Biology,2010,85(2):183-206.
[3] HilleRisLambers J,Adler P B,Harpole W S,et al.Rethinking community assembly through the lens of coexistence theory[J].Annual Review of Ecology,Evolution,and Systematics,2012,43:227-248.
[4] 卜文圣,许涵,臧润国,等.不同采伐干扰方式对热带山地雨林谱系结构的影响[J].林业科学,2014,50(4):15-21.
[5] 宋凯,米湘成,贾琪,等.不同程度人为干扰对古田山森林群落谱系结构的影响[J].生物多样性,2011,19(2):190-196.
[6] Zhang Jian,Mayor S J,He Fangliang.Does disturbance regime change community assembly of angiosperm plant communities in the boreal forest?[J].Journal of Plant Ecology,2014,7(2):188-201.
[7] Yu Mingjian,Xu Xuehong,Ding Ping.Economic loss versus ecological gain:the outbreaks of invaded pinewood nematode in China[J].Biological Invasions.2011,13:1283-1290.
[8] 吴昊,丁建清.入侵生态学最新研究动态[J].科学通报,2014(6):438-448.
[9] Yoshimura A,Kawasaki K,Takasu F,et al.Modeling the spread of pine wilt disease caused by nematodes with pine sawyers as vector[J].Ecology,1999,80(5):1691-1702.
[10]宁眺,方宇凌,汤坚,等.松材线虫及其关键传媒墨天牛的研究进展[J].应用昆虫学报,2004,41(2):97-104.
[11] Shi Juan,Luo Youqing,Song Jiying,et al.Traits of Masson pine affecting attack of pine wood nematode[J].Journal of Integrative Plant Biology,2007,49(12):1763-1771.
[12] Fujihara M.Succession of secondary pine forests after pine wilt disease in San-yo district,Western Japan[J].Natural History Research,1995,3:161-171.
[13] Fujihara M.Structure of Pinus luchuensis forests affected by pine wilt disease in northern Taiwan[J].Natural History Research,1997,Special Issue 4:113-126.
[14]徐学红.松材线虫入侵对植物群落的影响[D].杭州:浙江大学,2005.
[15] Qian Hong,Zhang Jian.Using an updated time-calibrated familylevel phylogeny of seed plants to test for non-random patterns of life forms across the phylogeny[J].Journal of Systematics and Evolution,2014,52(4):423-430.
[16] Webb C O,Donoghue M J.Phylomatic:tree assembly for applied phylogenetics[J].Molecular Ecology Notes,2005,5(1):181-183.
[17] Webb CO.Exploring the phylogenetic structure of ecological communities:An example for rain forest trees[J].American Naturalist,2000,156(2):145-155.
[18] Webb C O,Ackerly D D,McPeek M A,et al.Phylogenies and community ecology[J].Annual Review of Ecology and Systematics,2002,33:475-505.
[19] R Core Team.R:A language and environment for statistical computing[M/OL].R Foundation for Statistical Computing,Vienna,Austria,2014.URL http://www.R-project.org/
[20]Kembel SW,Cowan P D,Helmus M R,et al.Picante:R tools for integrating phylogenies and ecology[J].Bioinformatics,2010,26(11):1463-1464.
[21] Oksanen J,Blanchet F G,Kindt R,et al.Vegan:community ecology package[M/OL].R package version 2.3-0,(2015-05-26).URL http://cran.r-project.org,https://github.com/vegandevs/vegan
[22]黄建雄,郑凤英,米湘成.不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响[J].植物生态学报,2010,34(3):309-315.
[23] 沈琪,张骏,朱锦茹,等.浙江省生态公益林植被恢复过程中物种组成及多样性变化[J].生态学报,2005,25(9):2131-2138.
[24] Chazdon R L.Beyond deforestation:restoring forests and ecosystem services on degraded lands[J].Science,2008,320(5882):1458-1460.
