载畜率对荒漠草原群落及植物功能群生物量的影响
2015-12-13古琛杜宇凡乌力吉陈万杰薛文杰赵萌莉
古琛,杜宇凡,乌力吉,陈万杰,薛文杰,赵萌莉
内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019
载畜率对荒漠草原群落及植物功能群生物量的影响
古琛,杜宇凡,乌力吉,陈万杰,薛文杰,赵萌莉*
内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019
以内蒙古短花针茅(Stipa breviflora)草原为研究对象,设置4个载畜率水平的随机区组实验,根据生活型和不同光合途径划分植物功能群,探讨载畜率对群落地上地下生物量和不同功能群生物量的影响。结果表明:群落地上生物量随载畜率的增大而显著降低(P<0.05),且二者呈负相关关系,回归方程Y=281.54-53.47X;地下生物量呈“T”字型分布,约80%生物量集中分布在0~20 cm土层中,载畜率对地下生物量及分布无显著影响(P>0.05);群落根冠比在7~12之间,随载畜率的增大而显著增大(P<0.05)。载畜率对C4植物地上生物量无显著影响(P>0.05),但其相对生物量随载畜率的增大而增大,C3植物地上生物量与相对生物量均随载畜率的增大而降低(P<0.05)。多年生丛生禾草地上生物量相对稳定,受载畜率的影响差异不显著(P>0.05);其相对生物量在群落中最高,且均随载畜率的增大而增大(P<0.05);灌木、半灌木和小半灌木的地上生物量随载畜率的增大而减小(P<0.05),但其相对生物量除重度放牧显著最低(P<0.05)外,其他处理之间无显著差异(P>0.05);多年生杂类草、多年生根茎禾草和一、二年生植物地上生物量与相对生物量均随载畜率的增大而显著降低(P<0.05),多年生根茎禾草在轻度放牧下就几乎消失,而一、二年生植物在重度放牧下消失。
荒漠草原;载畜率;功能群;生物量
GU Chen, DU Yufan, WU Liji, CHEN Wanjie, XUE Wenjie, ZHAO Mengli. Effects of Stocking Rates on the Biomass of Desert Steppe Community and Plant Functional Group [J]. Ecology and Environmental Sciences, 2015, 24(12): 1962-1968.
荒漠草原是亚洲中部特有的旱生性最强的草地类型,是典型草原与荒漠之间的缓冲带,约占内蒙古草原面积的10.7%(刘钟龄等,1985)。在生境条件、物种组成、群落结构和功能等方面展现了生态学上的独特性(韩国栋等,2007),对自然和人类活动的干扰较为敏感,与其它草原类型相比稳定性较差(李德新,1990)。短花针茅(Stipa breviflora)草原作为荒漠草原的典型代表,占温性荒漠草原总面积的11.2%。因气候干燥、生境严酷、植被低矮稀疏、群落结构简单,家畜放牧是该类型草地最主要、最直接、最经济的利用方式,也几乎是唯一方式(王忠武,2009)。由于气候变化和过度放牧等不合理利用,草地退化加剧。
放牧是草地生态系统中最重要的人为干扰因素之一。家畜是草地生态系统的重要组分,通过采食、践踏和排泄等方式影响草地的形态特征、群落结构和功能等(焦树英,2006;Semmartin et al., 2008;陈万杰等,2015)。植物功能群被认为是与特定环境因子直接相关的物种群体,对外界干扰和环境影响做出的反应具有很大的相似性(Kleyer,2002;Lavorel et al.,2002;王长庭等,2004),且其作为联系草地生态系统中环境、个体、种群以及各生态学过程的桥梁,是响应植被受环境干扰的基本单元(Woodward et al,1996)。有研究表明,植物功能群组成及功能群间的相互作用是影响群落生产力及稳定性的主要因子,对生态系统过程有重要的影响(Tilman et al.,1997;Hooper et al.,1997;Hector et al.,1999;白永飞等,2002)。因此,利用功能群的变化来反映放牧强度对草地群落生产力稳定性的影响不失为一种便捷而有效的方式(孙世贤等,2013;白永飞等,2000)。
以内蒙古短花针茅草原为研究对象,设置4个载畜率水平的随机区组实验,在群落和功能群尺度上研究载畜率对荒漠草原的影响,旨在探讨以下 3个问题:(1)不同载畜率对群落地上地下生物量的影响结果如何?(2)不同生活型和C3/C4植物功能群地上生物量随载畜率如何变化?(3)植物功能群之间的补偿作用对群落稳定性的影响如何?以此阐明不同植物功能群对载畜率的响应,为其生态系统的保护和放牧管理利用提供理论依据。
1 材料与方法
1.1试验地概况
试验地位于阴山北麓,内蒙古自治区乌兰察布市西北部的葛根塔拉草原,四子王旗王府一队,内蒙古农牧科学院草原研究所荒漠草原实验基地(41°47′17″N,111°53′46″E)。地势平坦开阔,海拔1450 m。
该旗地处中温带大陆性季风气候区,具有干旱、少雨、多风和蒸发量大的气候特征。年均气温3.4 ℃,月平均温度最高月为6─8月,分别为21.5、24.0和 23.5 ℃;≥0 ℃积温为 2500~ 3100 ℃,≥10 ℃的年积温为2200~2500 ℃,无霜期90~115 d。平均日照时数为 3117.7 h。年均降水量 299.4 mm,且年际变化较大,降水主要集中在5─9月,占全年总降水总量的 80%以上。年蒸发量约 2300 mm,大大超过了降水量,是降水量的7~10倍。湿润度0.15~0.30干旱严重。
草地植被类型为短花针茅+冷蒿(Artemisia frigida)+无芒隐子草(Cleistogenes songorica)。草层低矮,平均高度8 cm左右,盖度为17%~20%,植被较稀疏。植物群落由 20多种植物组成,主要为多年生草本短花针茅、无芒隐子草、银灰旋花( Convolvulus ammannii)、 阿 尔 泰 狗 娃 花( Heteropappus altaicus)、 冰 草 ( Agropyron cristatum)、羊草(Leymus chinensis)、细叶葱(Allium tenuissimum)、蒙古葱(Allium mongolicum)等;一、二年生草本有猪毛菜(Salsola collina)、猪毛蒿(Artemisia scoparia)、栉叶蒿(Artemisia pectinata)等。还有灌木小叶锦鸡儿(Caragana microphylla)、矮灌木狭叶锦鸡儿(C. stenophylla)、半灌木驼绒藜(Ceratoides latens)、小半灌木木地肤(Kochia prostrata)、冷蒿等。
土壤为淡栗钙土,土层厚度大约为1 m,但在40~50 cm的地方出现了钙积层,土体坚硬且渗透能力差。土壤微生物有好气性细菌、放线菌和霉菌等。
1.2试验设计和测定方法
1.2.1试验设计
放牧试验于2003年6月开始延续至今,在内蒙古乌兰察布市四子王旗的短花针茅草原上,选取地势相对平坦、植被均一的约 50 hm2天然草地围封,进行不同载畜率放牧试验。采用完全随机区组设计,将围封的草地分为12个小区,3个区组(3次重复),每个区组中设4个处理(4个载畜率水平)(图1),分别为对照(CK)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG)。依据王明玖等(1994)、卫志军等(2000)对短花针茅草原载畜率的研究,再结合草地实际调查的结果,载畜率值依次为 0、0.91、1.82和2.71羊单位/(公顷·半年),绵羊只数分别为0、4、8和12只。每个区组内的4个处理完全随机排列,每个处理的面积基本相等,为4.4 hm2。放牧使用当地成年蒙古羯羊。2003—2013年每年放牧期为半年,从6月1日开始,到11月30日结束。每日早晨6点将绵羊赶入各小区让其自由采食,下午6点赶回圈内饮水、休息。
图1 试验小区和区组示意图Fig. 1 Schematic diagram is for experimental plot and block
1.2.2测定方法
2013年5月开始放牧前在每个小区内随机用铁笼围封10个1 m×1 m的样方。试验野外采样于2013年8月中旬群落生物量的最高峰时期进行。在已经围封好的样方内测定地上生物量,分种剪样后带回实验室,在65 ℃下烘干24 h至恒重后称其干重。在样方的正下方用直径为7 cm的根钻,分4层取0~10 cm、10~20、20~30、30~40 cm的地下生物量,每层取2钻混合,带回实验室后分别漂洗干净,在65 ℃下烘干24 h至恒重后称其干重。
1.2.3植物功能群划分
本文采用两种植物功能群的划分方法,第一,将植物按生活型分为5类:灌木、半灌木和小半灌木,多年生丛生禾草,多年生根茎禾草,多年生杂类草和一、二年生草本(白永飞等,2002)。第二,依不同的光合作用途径将植物分成C3植物和C4植物两类(唐海萍等,2001)。
植物功能群的相对生物量(%)=植物功能群地上生物量/群落地上生物量×100%
1.2.4数据分析方法
试验数据用Excel2010软件进行数据整理和简单分析后做图和表;不同载畜率对生物量是否具有显著影响,用SAS.9.2在α=0.05显著性水平下进行单因素方差分析(one-way ANOVA);载畜率与生物量间的关系式应用一元线性回归进行分析。
2 结果与分析
2.1群落地上生物量
短花针茅草原群落地上生物量随载畜率的增加而显著降低(P<0.05),对照最高为276.1 g·m-2,轻度放牧246.7 g·m-2、中度放牧172.7 g·m-2次之、重度放牧139.7 g·m-2最低;且地上生物量与载畜率呈显著负相关(P<0.05),回归方程 Y=281.54-53.47X,决定系数R2=0.66。对照群落地上生物量仅比轻度放牧高12%,但比中度放牧和重度放牧分别高60%和98%;轻度放牧地上生物量比中度放牧和重度放牧分别高43%和77%;中度放牧比重度放牧高24%(图2)。

图2 不同载畜率群落地上生物量Fig. 2 Aboveground biomass of community is under different stocking rates
2.2不同功能群地上生物量
2.2.1C3/C4植物地上生物量
由图3知,在不同载畜率下C4植物地上生物量表现出轻度放牧>中度放牧>对照>重度放牧的趋势,但彼此间无显著差异(P>0.05)。C3植物地上生物量则随载畜率的增加而降低,对照最高为202.1 g·m-2,轻度放牧次之为168.3 g·m-2,中度放牧与重度放牧最低,分别为108.7和89.0 g·m-2,除后两者之间无显著差异(P>0.05)外,其他处理间差异显著(P<0.05)。由载畜率与C3植物地上生物量的回归分析,得出载畜率与地上生物量呈显著负相关性(P<0.05),回归方程 Y=202.1-44.17X,决定系数R2=0.60。

图3 不同载畜率C3/C4植物地上生物量Fig. 3 Aboveground biomass of C3/C4plant is under different stocking rates
C3植物的相对生物量随载畜率的增大而减小,对照的79.0%显著高于中度放牧的65.6%和重度放牧的67.7%(P<0.05),轻度放牧的73.1%与其他处理相比无显著差异(P>0.05)。相反,C4植物的相对生物量随载畜率的增大而增大,中度放牧和重度放牧显著高于对照(P<0.05)(图4)。

图4 不同载畜率C3/C4植物相对生物量Fig. 4 Relative biomass of C3/C4plant is under different stocking rates
2.2.2不同生活型植物地上生物量
由图5、图6知,不同生活型植物地上生物量及相对生物量对载畜率的响应不同。
多年生丛生禾草地上生物量在 100~120 g·m-2之间,表现出随载畜率的增大而上升的趋势,但彼此间无显著差异(P>0.05);但其相对生物量在群落中最高,且随载畜率的增大而显著增加(P<0.05),对照与轻度放牧最小分别为37.5%和46.9%,中度放牧66.3%居中,重度放牧87.9%最高。
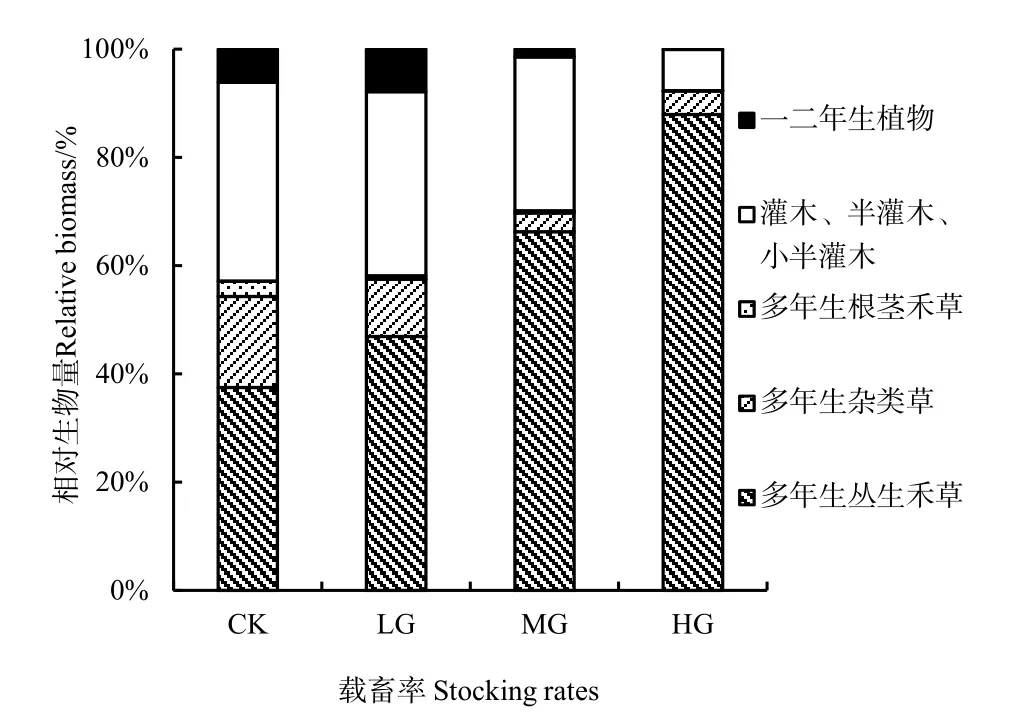
图6 不同载畜率不同生活型植物相对生物量Fig. 6 Relative biomass of plant life forms under different stocking rates
灌木、半灌木和小半灌木的地上生物量随载畜率的增大而减小,对照和轻度放牧最高,分别为94.3和79.1 g·m-2,中度放牧次之为47.2 g·m-2,重度放牧最低10.8 g·m-2,除前两者无差异显著(P>0.05)外,其他处理间差异显著(P<0.05);但其相对生物量除重度放牧的7.6%显著最低(P<0.05)外,其他处理在30%~35%之间,无显著差异(P>0.05)。
多年生杂类草地上生物量随载畜率的增大而显著降低(P<0.05),中度放牧与重度放牧间无显著差异(P>0.05),分别为5.9和5.7 g·m-2,但二者显著低于其他处理(P<0.05);轻度放牧为24.4 g·m-2居中,重度放牧的40.7 g·m-2最高。其相对生物量也有相同的趋势,对照最高为 16.8%,轻度放牧10.6%次之,中度放牧与重度放牧最低分别为3.5%和4.4%,除后两者无显著差异外(P>0.05),其他处理间差异显著(P<0.05)。
一、二年生植物地上生物量重度放牧仅为 0.1 g·m-2最少,中度放牧2.7 g·m-2居中,对照与轻度放牧的16.6和18.0 g·m-2最高,除后二者之间差异不显著(P>0.05)外,其他处理间差异显著(P<0.05)。其相对生物量对照与轻度放牧的6.1%和7.9%显著高于中度放牧与重度放牧的1.5%和0.1%(P<0.05)。
多年生根茎禾草地上生物量和相对生物量随载畜率的增大而降低,对照最高为 6.12 g·m-2和 2.8%,显著高于其他处理(P<0.05);轻度放牧与中度放牧次之,分别为 1.70 g·m-2、0.6%和 0.67 g·m-2、0.3%,二者之间差异不显著(P>0.05)。重度放牧中多年生根茎禾草消失。
2.3群落地下生物量及其分布
群落地下生物量呈随载畜率的增大而减少的趋势,对照1892.52 g·m-2、轻度放牧1842.53 g·m-2、中度放牧1591.17 g·m-2和重度放牧1493.57 g·m-2,但彼此间均无显著差异(P>0.05)(图7)。

图7 不同载畜率群落地下生物量Fig. 7 Belowground biomass of community is under different stocking rates
放牧对地下0~10、20~30、30~40 cm的生物量无显著影响(P>0.05)。10~20 cm的生物量轻度放牧显著高于中度和重度放牧(P<0.05),其他处理间均无显著差异(P>0.05)。群落地下生物量的垂直分布均是自上而下递减,0~10 cm的生物量最高,占0~40 cm生物量50%以上。0~20 cm的生物量约占0~40 cm生物量的80%(表1)。
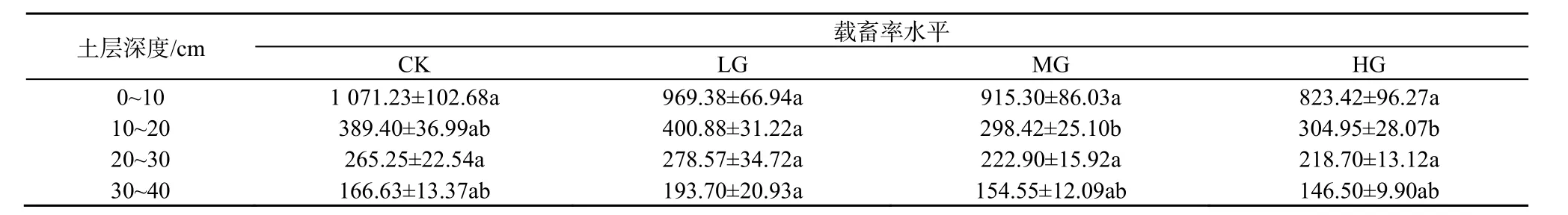
表1 不同载畜率水平地下生物量分布Table 1 The distribution of underground biomass under different stocking rates g·m-2
2.4群落根冠比
群落根冠比随载畜率的增大而增大,重度放牧最高11.8,中度放牧次之为9.7,轻度放牧与对照最小,分别为7.7和7.2,除后两者间差异不显著(P>0.05)外,其他处理间差异显著(P<0.05,图8)。
3 讨论
3.1载畜率对群落地上地下生物量及分配的影响
放牧是天然草原的主要影响因子和重要的进化驱动力,可以改变草地的形态特征、生产力以及群落结构(李永宏等,1999)。生物量作为草地生态系统最基本的数量特征,对草原生态系统结构和功能起着关键性作用(陈佐忠等,2000)。

图8 不同载畜率群落根冠比Fig. 8 Root shoot ratios of community are under different stocking rates
放牧优化假说认为适度放牧有利于草地生物量的增加,即生物量随载畜率的增大呈先上升后下降的非线性关系(Mcnaughton,1979);适度放牧通过加速营养循环、改善冠幅状况、增大光合面积、促进资源再分配等方式使牧草补偿或超补偿性生长,从而使草地生产力达到较高水平(Oesterheld et al.,1991);Semmartin et al.(2008)认为在放牧条件下植物体现出一种明显的响应机制来提高生长率。但是许多对内蒙古典型草原和荒漠草原的研究表明,随着放牧率的增大地上生物量呈现线性下降(王艳芬等,1999a;安渊等,2001;方楷等,2012)。本试验的结论与后者一致,因为随着载畜率的增大,家畜的采食和践踏对草地的破坏程度增强,减少光合面积的同时,光合产物的积累也随之降低;而且载畜率的增大造成植物的再生能力下降,最终导致群落地上生物量与载畜率呈显著负相关,随载畜率的增大而直线下降(图2)。
不同研究区域的不同草地类型关于放牧对群落地下生物量影响的结论略有差异。Pucheta et al.(2004)、梁燕等(2008)研究结果表明,适度放牧可使根系生物量增加,但重度放牧使之显著降低;李怡等(2011)、王艳芬等(1999b)、锡林图雅等(2009)的研究表明,0~30 cm地下生物量均随着载畜率的增大而呈线性下降。放牧通过影响土壤理化性质影响地下根系的生长发育。随着放牧强度的增加,家畜对草地的践踏增强,土壤的紧实度增大,土壤渗透性、含水量、孔隙度降低,土壤表层容重逐渐增大(戎郁萍等,2001;周丽艳等,2005;安慧等,2013),影响了根系伸长与扩展,再加上放牧使其光合作用减弱,输入根部的养分减少,极大地减弱了根系生物量(董亭等,2011)。本试验结果虽然表现出地下生物量随着载畜率的增加而降低的趋势,但无显著性差异(图7),可能是由降水及其季节分布导致,还要结合多年的地下生物量数据及气象数据进一步讨论。
国内外大量研究表明,各草地类型地下生物量主要分布在 0~30 cm土层中,约占地下生物量的80%~85%,且主要集中于0~10 cm,沿土层深度增加急剧下降,呈明显“T”字型或倒金字塔型分布(Richards et al.,1988;Mcnaughton et al.,1998;Greenwood et al.,1999;孙熙麟等,2011;宇万太等,2001)。范国艳(2010)、王长庭等(2008)指出放牧条件下,0~10 cm的根系生物量占0~40 cm的65%左右。而本试验得出0~10 cm根系生物量占0~40 cm总根系生物量的50%以上(表1)。但不同载畜率未对地下生物量的分布造成影响,未表现出放牧使地下生物量向土壤表层集中的趋势(Coupland,1979;Greenwood et al.,1998;王艳芬,1999b)。
植物在不同环境下的地上、地下生物量分配反映了其生长对环境的响应规律。放牧能够改变植物及群落生物量固有的物质与能量分配模式,而根冠比作为重要的参数之一,在草地生态系统中有重要的意义。马文红等(2006)、王亮等(2010)认为荒漠草原群落的根冠比在 2.0~32.2之间,平均为8.7,本试验的结果与以上结论保持一致(图 8)。由于家畜的采食和践踏使得地上部分不能充分生长,地上生物量减少的幅度常大于地下部分,造成根冠比随载畜率的增大而增大(李旭东等,2012)。
3.2载畜率对不同功能群地上生物量的影响
植物功能群常作为相对统一的整体对环境的波动或外界干扰做出响应。不同光合途径的植物,无论是地理分布还是叶片组织结构和生理功能,均体现了对光、水、热的响应与适应,是理想的植物功能群分类(韩梅等,2006)。与 C3植物相比,C4植物具有低CO2补偿点、高光饱和点和净光合速率等特点,对水分、光和CO2的利用效率高于C3植物。
本试验得出 C3植物地上生物量及相对生物量均随载畜率的增大而减少,C4植物地上生物量基本不受载畜率的影响,但其相对生物量随载畜率的增大而上升(图3,图4)。这意味着C3植物的作用随载畜率的增大而减弱,C4植物的作用随载畜率的增大而增强(王国杰等,2005)。放牧使群落地上生物量中C3植物所占的比例显著降低,而C4植物的比例明显增加,说明C4植物在放牧干扰下表现出较强的适应性,对C3植物生物量的减少和群落生产力水平起到了一定的补偿作用。短花针茅草原的 C4植物主要有木地肤、无芒隐子草和猪毛菜。放牧造成植被覆盖度降低,土壤水分减少,地表温度升高,土壤进一步干旱化和盐碱化,而C4植物对干旱、高温、盐碱等环境胁迫具有特殊适应性。因此放牧使耐旱性更强的无芒隐子草和木地肤以及既耐旱又耐盐碱的猪毛菜在群落中的作用增强。通过植物不同光合途径功能群的调节是维持草地生态系统稳定性的重要机制(韩梅等,2006)。
植物生活型是植物对环境长期适应而具有一定的形态外貌、结构和习性的植物类型,而植物的生活型功能群也是环境因子的综合反映(江小蕾等,2003)。本试验得出多年生丛生禾草地上生物量相对稳定,受载畜率的影响差异不显著;其相对生物量在群落中最高,且均随载畜率的增大而增加大,灌木、半灌木和小半灌木的地上生物量随载畜率的增大而减小,但其相对生物量除重度放牧显著最低外,其他处理之间无显著差异;多年生杂类草、多年生根茎禾草和一、二年生植物地上生物量与相对生物量均随载畜率的增大而显著降低,多年生根茎禾草在轻度放牧下就几乎消失,而一、二年生植物在重度放牧下消失(图5,图6)。
短花针茅草原的多年生丛生禾草主要有短花针茅和无芒隐子草,二者分别是该草原的建群种和优势种,有较强的耐牧机制。随着载畜率的增大,多年生丛生禾草的株丛破碎化加重、丛幅变小,只能通过增大株丛密度以进一步适应环境;再加上其他功能群随载畜率的增大而逐渐消失,最终导致该功能群相对生物量逐渐升高,在群落中的作用增大。随载畜率的增大,冷蒿的优势地位是灌木、半灌木和小半灌木虽然生物量降低但相对生物量保持稳定的重要原因。随载畜率的增大,多年生杂类草、多年生根茎禾草和一、二年生植物的生物量与相对生物量逐渐减少甚至消失,说明在干旱的荒漠草原地区放牧不利于它们的生长,使之在群落中的作用减弱,造成群落的组成、结构和功能趋于简单化。羊草是多年生根茎禾草中的主要物种,是中旱生植物,靠根茎繁殖,在干旱的荒漠草原原本就不占优势,再加上随着载畜率的增加,土壤容重、紧实度增大,而土壤含水量下降,严重影响了多年生根茎禾草和一、二年生植物的生长和繁殖,导致它们随载畜率的增大而减少甚至消失(王国杰等,2005;焦树英等,2006a,2006b)。
4 结论
(1)群落地上生物量随载畜率的增大而显著降低,且二者间呈显著负相关;载畜率对地下生物量及分布无显著影响,不同载畜率地下生物量呈“T”字型分布,约80%生物量集中分布在0~20 cm土层中;不同处理根冠比在 7~12之间,随载畜率的增大而增大。
(2)载畜率对C4植物地上生物量无显著的影响,但其相对生物量随载畜率的增大而增大,中度放牧和重度放牧显著高于对照;C3植物地上生物量与相对生物量均随载畜率的增加而降低。
(3)多年生丛生禾草地上生物量与相对生物量在群落中最高,且均随载畜率的增大而增加;灌木、半灌木和小半灌木的地上生物量随载畜率的增大而减小,但其相对生物量除重度放牧显著最低外,其他处理之间无显著差异;多年生杂类草、多年生根茎禾草和一、二年生植物地上生物量与相对生物量均随载畜率的增大而显著降低。
COUPLAND R T. 1979. Grassland ecosystems of the world: analysis of grasslands and their uses [M]. Cambridge, Cambridge University Press.
GREENWOOD K L, HUTCHINSON K J. 1998. Root characteristics of temperate pasture in New South Wales after grazing at three stocking rates for 30years [J]. Grass and Forage Science, 53(2): 120-128.
HECTOR A, SCHMID B, BEIERKUHNLEIN C, et al. 1999. Plant diversity and productivity experiments in European grasslands [J]. Science, 286(5442): 1123-1127.
HOOPER D U, VITOUSEK P M. 1997. The effects of plant composition and diversity on ecosystem processes [J]. Science, 277(5330): 1302-1305.
KLEYER M. 2002. Validation of plant functional types across two contrasting landscapes [J]. Journal of Vegetation Science, 13(2): 167-178.
LAVOREL S, GARNIER E. 2002. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail [J]. Functional Ecology, 16(2): 545-556.
MCNAUGHTON S J, BANYIKWA F F, MCNAUGHTON M M. 1998. Root biomass and productivity in a grazing ecosystem: the Serengeti [J]. Ecology, 79(2): 587-592.
MCNAUGHTON S J. 1979. Grazing as an optimization process grass-ungulate relationships in the Serengeti [J]. The American Naturalis, 113(5): 691-703.
OESTERHELD M, MCNAUGHTON S J. 1991. Effect of stress and time for recovery on the amount of compensatory growth after grazing [J]. Oecologia, 85(3): 305-313.
PUCHETA E, BONAMICI I, CABIDO M, et al. 2004. Below-ground biomass and productivity of grazed site and a neighbouring ungrazed enclosure in a grassland in central Argentina [J]. Austral Ecology, 29(2): 201-208.
RICHARDS J H, MUELLER R J, MOTT J J. 1988. Tillering in tussock grasses in relation to defoliation and apical bud removal [J]. Annals of botany, 62(2): 173-179.
SEMMARTIN M, GARIBALDI L A., CHANETON E J. 2008. Grazing history effects on above-and below-ground litter decomposition and nutrient cycling in two co-occurring grasses [J]. Plant and Soil, 303(1): 177-189.
TILMAN D, KNOPS J, WEDIN D, et al. 1997. The influence of functional diversity and composition on ecosystem processes [J]. Science Magazine, 277(5330): 1300-1302.
WOODWARD F I, CRAMER W. 1996. Plant functional types and climatic changes: introduction [J]. Journal of Vegetation Science, 7(3): 306-308.
安慧, 徐坤. 2013. 放牧干扰对荒漠草原土壤性状的影响[J]. 草业学报, 22(4): 35-42.
安渊, 李博, 杨持, 等. 2001. 内蒙古大针茅草原草地生产力及其可持续利用研究: 放牧系统植物地上现存量动态研究[J]. 草业学报, 10(2): 22-27.
白永飞, 陈佐忠. 2000. 锡林河流域羊草草原植物种群和功能群的长期变异性及其对群落稳定性的影响[J]. 植物生态学报, 24(6): 641-647.
白永飞, 张丽霞, 张焱, 等. 2002. 内蒙古锡林河流域草原群落植物功能群组成沿水热梯度变化的样带研究[J]. 植物生态学报, 26(3):308-316.
陈万杰, 董亭, 古琛, 等. 2015. 不同放牧强度对大针茅根系特征的影响[J]. 中国草地学报, 37(4): 86-91.
陈佐忠, 汪诗平. 2000. 中国典型草原生态系统[M]. 北京: 科学出版社.
董亭, 李群, 赵萌莉, 等. 2011. 放牧对大针茅根系生物量影响的研究[J].草地学报, 19(2): 237-241.
范国艳. 2010. 放牧对贝加尔针茅草原群落地上生物量和根系分布特征的影响[D]. 兰州: 甘肃农业大学.
方楷, 宋乃平, 魏乐, 等. 2012. 不同放牧制度对荒漠草原地上生物量及种间关系的影响[J]. 草业学报, 21(5): 12-22.
韩国栋, 焦树英, 毕力格图, 等. 2007. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响[J]. 生态学报, 27(1): 184-190.
韩梅, 杨利民, 张永刚, 等. 2006. 中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应[J]. 生态学报, 20(6): 1825-1832.
江小蕾, 张卫国, 杨振宇, 等. 2003. 不同干扰类型对高寒草甸群落结构和植物多样性的影响[J]. 西北植物学报, 23(9): 1479-1485.
焦树英, 韩国栋, 李永强, 等. 2006a. 不同载畜率对荒漠草原群落结构和功能群生产力的影响[J]. 西北植物学报, 26(3): 564-571.
焦树英, 韩国栋, 赵萌莉, 等. 2006b. 荒漠草原地区不同载畜率对功能群特征及其多样性的影响[J]. 干旱区资源与环境, 20(1): 161-165.
焦树英. 2006. 短花针茅草原生态系统对不同载畜率水平绵羊放牧的响应[D]. 呼和浩特: 内蒙古农业大学.
李德新. 1990. 短花针茅荒漠草原动态规律及其生态稳定性[J]. 中国草地, (4): 1-5.
李旭东, 张春平, 傅华. 2012. 黄土高原典型草原草地根冠比的季节动态及其影响因素[J]. 草业学报, 21(4): 307-312.
李怡, 韩国栋. 2011. 放牧强度对内蒙古大针茅典型草原地下生物量及其垂直分布的影响[J]. 内蒙古农业大学学报, 32(2): 89-92.
李永宏, 汪诗平. 1999. 放牧对草原植物的影响[J]. 中国草地, (3): 11-19.
梁燕, 韩国栋, 周禾. 2008. 羊草草原不同退化程度植物群落内地上部分变化对群落根系的影响[J]. 草业科学, 25(4): 110-115.
刘钟龄, 雍世鹏, 王义风. 1985. 内蒙古植被[M]. 北京: 科学出版社.
马文红, 方精云. 2006 .内蒙古温带草原的根冠比及其影响因素[J]. 北京大学学报:自然科学版, 42(6): 774-778.
戎郁萍, 韩建国, 王培, 等. 2001. 放牧强度对草地土壤理化性质的影响[J]. 中国草地, 23(4): 42-48.
孙世贤, 卫智军, 吕世杰, 等. 2013. 放牧强度季节调控下荒漠草原植物群落与功能群特征[J]. 生态学杂志, 32(10): 2703-2710.
孙熙麟, 王明玖, 陈海军, 等. 2011. 短花针茅荒漠草原地下生物量对不同强度放牧的响应[J]. 内蒙古农业大学学报, 31(4): 101-104.
唐海萍, 刘书润. 2001. 内蒙古地区的C4植物名录[J]. 内蒙古大学学报:自然科学版, 32(4): 431-438.
王国杰, 汪诗平, 郝彦宾, 等. 2005. 水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响[J]. 生态学报, 25(7): 1650-1656.
王亮, 牛克昌, 杨元合, 等. 2010. 中国草地生物量地上—地下分配格局:基于个体水平的研究[J]. 中国科学C辑: 生命科学, 40(7): 642-649.
王明玖, 马长升. 1994. 两种方法估算草地载畜量的研究[J]. 中国草地, (5): 19-22.
王艳芬, 汪诗平. 1999a. 不同放牧率对内蒙古典型草原牧草地上现存量和净初级生产力及品质的影响[J]. 草业学报, 8(1): 15-20.
王艳芬, 汪诗平. 1999b. 不同放牧率对典型草原地下生物量的影响[J].草地学报, 7(3): 198-203.
王长庭, 龙瑞军, 丁路明. 2004. 高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响[J]. 生物多样性, 12(4): 403-409.
王长庭, 王启兰, 景增春, 等. 2008. 不同放牧梯度下高寒小嵩草草甸植被根系和土壤理化特征的变化[J]. 草业学报, 17(5): 9-15.
王忠武. 2009. 载畜率对短花针茅荒漠草原生态系统稳定性的影响[D].呼和浩特: 内蒙古农业大学.
卫智军, 韩国栋, 杨静, 等. 2000. 短花针茅荒漠草原植物群落特征对不同载畜率水平的响应[J]. 中国草地, (6): 1-5.
锡林图雅, 徐柱, 郑阳. 2009. 不同放牧率对内蒙古克氏针茅草原地下生物量及地上净初级生产量的影响[J]. 中国草地学报, 31(3): 26-29.
宇万太, 于永强. 2001. 植物地下生物量研究进展[J]. 应用生态学报, 12(6): 927-932.
周丽艳, 王明玖, 韩国栋. 2005. 不同强度放牧对贝加尔针茅草原群落和土壤理化性质的影响[J]. 干旱区资源与环境, 19(7): 182-187.
Effects of Stocking Rates on the Biomass of Desert Steppe Community and Plant Functional Group
GU Chen, DU Yufan, WU Liji, CHEN Wanjie, XUE Wenjie, ZHAO Mengli*
Collage of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China
Desert steppe of Stipa breviflora in Inner Mongolia was taken as the object, and randomized block experiment was set based on 4 stocking rates, as well as plant functional groups were divided according to different life forms and photosynthetic pathways to study the effects of different stocking rates on above-and-underground biomass of community and the biomass of different functional groups. The results indicated that with stocking rates raised, aboveground biomass of community was dropping (P<0.05), and they were negative correlated as well, the regression equation was Y=281.54-53.47X. Belowground biomass distributed like “T”, and about 80 percent of it concentrated on 0~20 cm soil, meanwhile, it was not effected significantly by stocking rates (P>0.05). However, the root shoot ratio which between 7 and 12 was increasing with the stocking rates added. Aboveground biomass of C4was also not influenced remarkably, whereas its relative biomass was upgrading and both of aboveground and relative biomass of C3were reducing markedly by stocking rates increased (P<0.05). Aboveground biomass of perennial bunchgrass was not impacted (P>0.05) but its relative biomass, the highest one in community, was advancing with stocking rates ascended (P<0.05). On the contrary, the aboveground biomass of shrub, subshrub and semishrub was declining (P<0.05), whereas its relative biomass was stable except it was the lowest in heavy grazing (P>0.05). Both aboveground and relative biomass of perennial forbs, perennial rhizome grasses, annuals and biennials were decreasing respectively by stocking rates enlarged (P<0.05), and even perennial rhizome grasses practically disappeared under light grazing as well as annuals and biennials vanished under heavy grazing.
desert steppe; stocking rates; functional group; biomass
10.16258/j.cnki.1674-5906.2015.12.006
Q948; X171
A
1674-5906(2015)12-1962-07
国家自然科学基金项目(31460110);内蒙古农业大学草业与草地资源重点实验室
古琛(1988年生),男,博士研究生,主要从事草地生态学方向研究。E-mail: guchen0706@sina.com *通信作者。E-mail: nmgmlzh@126.com
2015-08-26
引用格式:古琛, 杜宇凡, 乌力吉, 陈万杰, 薛文杰, 赵萌莉. 载畜率对荒漠草原群落及植物功能群生物量的影响[J]. 生态环境学报, 2015, 24(12): 1962-1968.
