贡嘎山海螺沟冰川退缩区原生演替序列植被生物量动态
2015-12-08杨丹丹罗辑佘佳唐荣贵
杨丹丹,罗辑,佘佳,唐荣贵
1. 中国科学院成都山地灾害与环境研究所,四川 成都 610041;2. 中国科学院大学,北京 100049;3. 成都市环境保护科学研究院,四川 成都 610031
贡嘎山海螺沟冰川退缩区原生演替序列植被生物量动态
杨丹丹1,2,罗辑1*,佘佳3,唐荣贵1
1. 中国科学院成都山地灾害与环境研究所,四川 成都 610041;2. 中国科学院大学,北京 100049;3. 成都市环境保护科学研究院,四川 成都 610031
基于对贡嘎山海螺沟冰川退缩后形成的125 a的原生演替序列上不同森林群落类型的调查,以空间代替时间的方法,选取了7个典型样地(S0~S6),分别代表冰川退缩后第0、17、35、49、56、85和125年后的演替群落,探讨了不同演替阶段生态系统各组分生物量变化规律及分配特征。结果表明,群落生物量与演替阶段和乔木层优势种的组成密切相关。乔木层生物量与活植物体总生物量均随演替的进行呈显著的指数增长的趋势,分别从 10.195 Mg·hm-2增至 366.122 Mg·hm-2,从9.162 Mg·hm-2增至332.461 Mg·hm-2;不同演替阶段乔木层生物量在各个层次分配中占绝对优势(>89.871%),其他各层所占比例较小,总趋势为:灌木层>地被层>草本层,林下各层生物量分配受到群落环境影响较大。粗木质物残体量和年叶凋落物量也随着演替的进行不断积累,其中粗木质物残体量在针阔混交林阶段(S5)达到最高,年叶凋落物量则随演替的进行呈显著的指数增加的趋势。演替前60年(S0~S4),柳树(Salix rehderana)、沙棘(Hippophae rhamnoides)和冬瓜杨(Populus purdomii)等落叶阔叶树种对乔木层生物量贡献最大,演替后60年(S5~S6),乔木层生物量则主要来自冷杉(Abies fabri)和云杉(Picea brachytyla)等针叶树种(>93.070%);乔木层生物量的器官分配以树干所占比例最高,为56.388%~72.658%,枝和根的比例次之,叶所占比例则最小。经过了125 a的演替,海螺沟冰川退缩区生态系统植被生物量已达到成熟林水平,生态系统结构与功能相对稳定,植被演替发展至顶级群落。
海螺沟;冰川退缩区;原生演替;生物量
YANG Dandan, LUO Ji, SHE Jia, TANG Ronggui. Dynamics of Vegetation Biomass Along the Chronosequence in Hailuogou Glacier Retreated Area, Mt. Gongga [J]. Ecology and Environmental Sciences, 2015, 24(11): 1843-1850.
极地或高山冰川消退区所形成的冰川裸地作为研究植被原生演替的理想场所,引起了很多生态学家的关注(Crocker et al.,1955;Whittaker,1993;Chapin et al.,1994;Vetaas,1994;Ohtonen et al.,1999;Hodkinson et al.,2002;Jones et al.,2003;许中旗等,2005)。主要采用空间代替时间的方法,假设不同地段冰川退缩区植被与环境演替总是朝着固定方向有序进行,前一个样地植被往往起到改善土壤环境条件的作用,从而为后一个样地植被的定居提供一定的环境基础(Cole et al.,1981;Kimmis,1987),生态学家通过对不同时期冰川退缩区典型样地的调查,就可以反演历史上完整的植被演替系列的动态(许中旗等,2005;Johnson et al.,2008)。最著名的是Crocker et al.(1955)在阿拉斯加Glacier Bay冰川所做的工作,他们发现在Glacier Bay冰川消退不到200 a的时间里,植被经历了由先锋草本群落到针阔混交林的演变,同时土壤条件也发生了明显的变化,而这些变化与演替年龄和植被发育有着密切的关系。然而,也有一些生态学家对这一方法提出了质疑(Collins et al.,1983;Pickett,1989;Johnson et al.,2008)。科学家通过对沙丘、冰川以及采矿迹地原生演替过程的研究发现,由于受到不同程度的干扰以及微地形等因素的影响,在年龄相同的地段上出现了原本被认为是处于不同阶段的群落类型(Cowles,1899;Olson,1958;Maun,1981;Wiegleb et al.,2001)。Chapin et al.(1994)通过对阿拉斯加Glacier Bay冰川退缩区的研究表明演替早期的物种总是抑制而不是促进后期物种的生长。Fastie(1995)在同一地点利用树木年轮法也得出了与之一致的结论。由此可见,原生演替过程中植被变化由于受到原生裸地的环境特征以及植物的生物学和生态学特性的影响而变得十分复杂。
海螺沟冰川自小冰期以来开始退缩,冰川退缩后形成了大量原生裸地,由于此处受到的人为干扰少,在其长达2 km的序列范围内生态因子变化小,因此形成了一个完整的从裸地到先锋群落再到顶级群落的连续植被原生演替序列,为我们研究原生演替过程与机制提供了一个天然试验场地。此前的科学家们主要从植被原生演替与气候变化的关系(李逊等,1995;霍常富等,2010)、植被原生演替的种间协变(罗辑,1996)和土壤的演替序列(何磊等,2007)等角度对海螺沟冰川裸地的原生演替系列进行了研究,而关于海螺沟冰川退缩区植被原生演替序列生物量动态变化的研究仍是一片空白。生物量作为生态系统在一定时期内积累的有机质总量,是整个生态系统运行的能量基础和营养物质来源,也是衡量生态系统生产力和元素循环的基础,对研究各生态系统的结构和功能具有重要作用(Overman et al.,1994;冯宗炜,1999)。一方面,森林生物量通常随林龄的增加而增加,在成熟林时接近一个稳定值(Saatchi et al.,2007),另一方面,生物量也会随不同群落类型、不同生境条件等的变化而变化(Hoshizaki et al.,2004;郑晓翾等,2007),同时会受到湿度、温度等环境条件及干扰等的变化的影响(Houghton,2005)。
本文以海螺沟冰川退缩区作为研究区域,以空间代替时间的方法,基于前人对冰川退缩区域植被演替完整序列的研究,对冰川退缩后形成的原生演替序列不同阶段植物群落的生物量在生态系统各个层次、种类组成和器官的变化及其分配进行了比较研究。这对于重建小冰期以来的环境变化过程,预测未来在全球气候变化背景下冰川退缩区域环境变化和植被演替的趋势具有重要意义,也可为揭示原生演替序列生物量变化的一般规律提供基础数据,同时对于植被恢复也具有一定的指导作用。
1 研究区概况与研究方法
1.1研究区概况
海螺沟冰川退缩区(29°34′21″N,102°59′42″E)位于贡嘎山东坡,青藏高原东南缘(图 1a)。该地区气候湿冷,属于山地寒温带气候类型,年平均气温3.8 ℃,1月平均气温最低(-4.38 ℃),7月平均气温最高(11.9 ℃),年降水日数在260 d以上,年平均降水量约1960 mm,年平均相对湿度约90%(Li et al.,2008)。海螺沟冰川属于季风海洋性冰川,水热条件好,冰川消融速度快,近百年来没有冰进过程,土壤有连续成土过程。海螺沟是贡嘎山东坡最主要冰川河,也是我国最具代表性的季风海洋性山谷冰川之一,自小冰期开始退缩(李宗省等,2009),20世纪30年代退缩加速(图1b)形成了大量原生裸地,在其长达2 km的序列范围内形成了一个完整的从裸地到先锋群落再到顶级群落的连续植被原生演替序列。

图1 海螺沟冰川退缩区采样点分布图Fig. 1 Map of sampling plots on Hailuogou Glacier retreat area
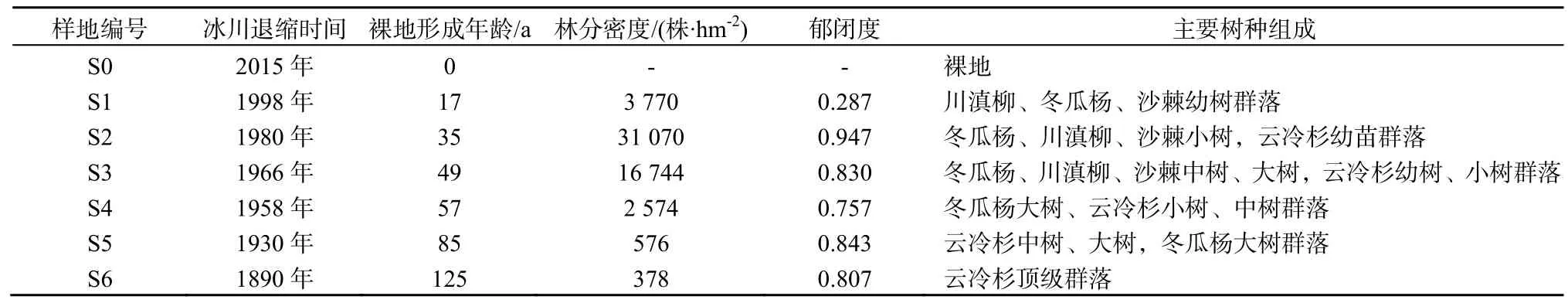
表1 不同演替阶段样地植被特征Table 1 species characteristics of plots at different stand ages
1.2研究方法
1.2.1 样品采集
本研究以120多年形成的冰川退缩区作为研究区,基于前人对海螺沟冰川退缩区植被演替不同阶段划分的研究上(陈富斌等,1998;钟祥浩等,1997),选取了7个典型样地(图1c),各样地冰川退缩时间、环境条件以及主要树种组成见表 1。于2013年10月和2014年10月在每个样地随机布设20 m×20 m样方,调查其中的乔木,记录其树种、胸径、树高、存活状态等,并在样地内随机布设 3个1 m×1 m样方,调查样方内所有灌木、草本和地被物,记录其种类、数量等,并取部分样品带回实验室烘干、称重;同时,设置3个1 m×1 m的小样方,收集样方内所有凋落叶,带回实验室烘干后称重。具体采集方法如下:
乔木层:采集优势种。样品按生物量等比例混合。叶与枝在同一时期采集。枝和叶按不同方位、不同部位采集;树干和树皮按上、中、下3个部位、不同方向(对大树而言,分为阴面和阳面)采集,茎干使用生长锥取树芯;根按照不同的方位采集细根(直径<2 mm)、中根(直径为2~10 mm)以及粗根(直径在10 mm以上)。
灌木层、草本层和地被层:记录1 m×1 m样方各种类后,将样方内所有灌木、草本和苔藓全部收获,灌木、草本按种混合分别采集分地上部分和地下部分,地被物作为一个整体整株采集。
凋落物:在各样地随机放置10个1 m×1 m的收集框用来收集凋落物。分月进行收获。
粗木质物残体:粗木质物残体包括立枯、倒木和占桩。分别采集立枯、倒木和占桩的皮和木质部,组成混合样。
土壤样品的采集采用挖土壤剖面法,每个样地挖取3个土壤剖面,分层进行取样,并挑出土壤混合样中的根系和石砾,带回实验室风干处理,进行土壤有机质、pH值以及C、N的测定。
1.2.2 生物量测定方法
乔木:乔木层生物量的采用生物量生长方程进行计算。
M=a×(D2H)b(1)
式中,M为乔木层生物量,D表示胸径,H表示树高,参数 a,b根据样本采用最小二乘法加以确定。各优势种生物量的计算采用罗天祥等建立的生物量方程(朱兴武等,1988;鄢武先等,1991;宿以明等,2000;Luo et al.,2002;刘兴良等,2006;李登秋等,2013)。
灌木、草本、苔藓和凋落物:采用收获法获得。每个样地均取1个1 m×1 m的代表性小方框。将方框内所有灌木、草本、苔藓和凋落物按种类全部收获后,灌木、草本分地上部分和地下部分,苔藓为全株,凋落物分枯枝和叶,烘干后分别称量其干重。
立枯、倒木:目测估量样方内立枯、倒木的干重,取样地内或附近易获得和易携带的、具代表性的立枯、倒木作为标准参照物带回实验室称量其干重后较正目测值。
1.2.3 数据处理
采用单因素方差分析法(one-way ANOVA)分析不同演替阶段植被生物量的差异,并采用最小显著差法(LSD)进行多重比较。数据分析软件为SPSS 16.0,作图软件为0rigin 8.0。
2 结果与分析
2.1原生演替不同阶段植被与环境特征
冰川退缩后的原生裸地(S0)主要由一些冰碛物组成,没有土壤的积累,养分状况较差,地面温度变幅也比以后任何时期都大,一些矿质养分的不足往往会成为影响植物定居、生长的限制因子(Hobbie et al,1998),其中以氮素和光的影响最为突出。
底碛经过3年的裸露和地形变化,土壤N含量相比于裸地有了明显提高,pH值略有下降,地面温度变幅也稍有缩小(表2),环境条件得到一定的改善。在原生裸地形成后的第4年就形成了以马河山黄芪(Astragalus mahoshanicus)、直立黄芪(A.adsurgens)为优势种的草本群落,最初的草本植物群落呈斑块状地岛分布,一些木本植物如川滇柳(Salix rehderana)、冬瓜杨(Populus purdomii)和沙棘(Hippophae rhamnoides)等则生长在草本植物周围,先锋群落中植物种类和数量较少。早期具有固氮作用的豆科植物黄芪为土壤中氮含量的初步累积做出了重要的贡献,改善了冰川退缩区域缺氮的状况,土壤有机质含量也大大增加,pH值则有所降低,在距今 17年左右的冰川退缩区(1998年样地,S1),形成了冬瓜杨、柳树和沙棘幼树群落(表1)。

表2 不同演替阶段样地环境与土壤特征Table 2 Environment and soil characteristics of plots at different stand stages
最初形成的稀疏的植物群落经过约10~20年的演替后,形成了相对郁闭的冬瓜杨、柳树和沙棘小树群落(1980年样地,S2),地面蒸发减少,调节了温度、湿度变化,黄芪优势度显著降低。而同样具有固氮根瘤的柳树和沙棘生长加快,土壤中N含量迅速增大,有机质积累增加,生境条件得到了极大的改善。
由于冰川退缩迹地水热条件较好,植物生长迅速,冬瓜杨凭借其较快的高生长速度和较高的光合速率,占据了更多的空间,群落郁闭度大大增加,此时林内生境有利于阴性植物峨眉冷杉(Abies fabri)和麦吊云杉(Picea brachytyla)的种子发育,它们先后进入林地(1966年样地,S3)。柳树和沙棘幼苗数量减少,以老树和壮树为主。
随着冬瓜杨生态位的拓展、种群的自疏和他疏作用加强,柳树和沙棘因得不到足够的光照和养分,逐渐在种间竞争中败下阵来,生长速度逐步减慢,演变为衰退种群,冬瓜杨高和径生长则保持在较高水平,占据着林上大片空间(1958年样地,S4)。同时由于种内和种间斗争,群落内大量林窗出现,林下灌木、草本植物种类与数量有明显的增加。土壤中N含量略有降低,有机质含量则达到了整个演替序列的最高值,pH值仍保持下降的趋势(表2)。
接着又经过近 20年的演替,植物群落特征和生境条件变化逐步减缓,群落郁闭度、空气湿度与温度达到了整个演替序列上的最大值。土壤逐步分化出淀积层,土壤A层中N含量变化不大,pH值继续降低,群落进入到以冬瓜杨和峨眉冷杉为优势种的针阔混交林阶段(1930年样地,S5)。这一阶段林下灌木与草本植物种类与数量相比于前一阶段有明显减少,可能是由于林冠层郁闭度增大使得灌木层和草本层植物所能利用的光能越来越少,最终使喜阳的灌木与草本逐渐衰减。
经历了125 a的演替后,群落最终进入到以峨眉冷杉和麦吊云杉为优势种的针叶林顶级群落(1890年样地,S6),云、冷杉由于生长加快逐渐占据主林层,冬瓜杨则由于接收不到足够的光照,逐步退出群落,同时由于云冷杉的自疏作用出现大量林窗,为灌木和草本的生长创造了充足的光照条件,其种类和数量又有所回升。但是此时土壤有机质以及N含量相比于前一阶段却减少了,土壤pH值也进一步降低(表2),土壤结构还不尽完善,有待继续发育。
2.2原生演替不同阶段群落生物量变化及分配特征
由图 2a可以看出,植被演替不同阶段群落总生物量与乔木层生物量变化表现出相似的趋势,即随着演替时间的增长,其生物量均呈现显著的指数增长(r2=0.991,P<0.001;r2=0.990,P<0.001)。群落总生物量从演替初期的10.195 Mg·hm-2逐渐增至顶级群落的366.122 Mg·hm-2,增加了35倍。就生态系统各层次而言,乔木层对活植物体总生物量贡献最大(>89.871%,图2c),由此可见,乔木层是原生演替不同阶段生物量的主体部分,乔木层生物量的增加是导致群落总生物量随演替进行不断增加的主要原因。乔木层生物量占总生物量比例最高值出现在样地S2,为98.380%,但是在S2以后,这一比例则随着演替的进行不断降低。

图2 原生演替不同阶段群落生物量变化及其分配Fig. 2 Biomass changes and distribution of vegetation along the chronosequence
其他各层生物量对总生物量的贡献较低(灌木层<7%,草本层<5%,地被层<5%),随着演替的进行,灌木层、草本层与地被层生物量表现出显著的阶段性的变化规律。根据图2b,我们可以将林下各层生物量变化划分为 3个阶段:S1~S4,S4~S5,S5~S6。从S1到S4,为落叶阔叶林阶段,林下各层生物量均随演替时间的增加而不断增加,其中灌木层生物量增长最为迅速;从S4到S5,群落处在由落叶阔叶林到针叶林的过渡阶段,灌木层生物量增加变缓,而草本层和地被层生物量则表现出完全相反的变化趋势,前者生物量迅速降低,后者则快速提高,地被层生物量在样地 S5达到了整个演替序列的最高值10.422 Mg·hm-2;最后当群落进入针叶林阶段以后(S5~S6),灌木层、草本层生物量进一步增加,而地被层生物量则略有降低,比 S5减少了0.966 Mg·hm-2。
2.3原生演替不同阶段乔木层生物量的分配特征
不同演替阶段,不同树种在群落结构中的比重不同,进而造成了乔木层生物量在各个树种间分配特征的差异(王晓莉等,2014)。原生裸地经过17 a的发育之后(S1),主要生长着柳树、沙棘和冬瓜杨等先锋树种,三者生物量之比为12∶7∶4,以柳树生物量所占比重最大(图3a);接下来的20~70 a(S2~S4),冬瓜杨生物量迅速由38.523 Mg·hm-2增至整个演替序列上的最大值189.668 Mg·hm-2,其占乔木层总生物量的百分比也由17.149%(S1)增至87.305%(S6)。而柳树和沙棘生物量则在S2达到最大值后逐渐降低,其占乔木层总生物量的比重也分别由53.126%和29.724%降至8.996%和3.563%。原生裸地经过 80多年的演替后,柳树和沙棘已基本完全退出群落,喜阳的冬瓜杨的生态位也渐渐被耐荫的云冷杉所占据,随着冬瓜杨等阔叶树种的退出,群落开始进入到以云冷杉为建群种的常绿针叶林阶段(S5),直接表现为云冷杉生物量自S4开始出现后就加速增长,在样地S6达到最高值331.745 Mg·hm-2,是冬瓜杨最盛时期(S4)生物量的 1.75倍,同时在这一阶段乔木层总生物量的分配中也占据了绝对优势(>99%)。

图3 原生演替不同阶段乔木层生物量变化及其分配Fig. 3 Biomass changes and distribution of tree layer along the chronosequence
不同演替阶段乔木层生物量的变化是由各个树种生物量决定的,而不同树种生物量最终又取决于各个器官生物量的大小。由图 3d可见,在整个演替序列上,树干占乔木层总生物量的比例一直是最大,在56.388%~72.658%之间;其次是枝和根,叶对乔木层总生物量贡献最小,不到 8%。演替前期,柳树、沙棘和冬瓜杨等落叶阔叶树种均处于营养生长较为旺盛的时期,各器官生物量也随林龄的增加不断生长,其中干和根的生物量的增长最为迅速,表现为净积累的过程,枝和叶生物量的增加则随演替的进行增加到一定阶段后开始出现下降趋势(图 3b);演替后期干和根生物量继续积累,而枝和叶生物量则在云冷杉完全取代冬瓜杨以后有所下降。
2.4原生演替不同阶段粗木质物残体量以及年叶凋落物量的变化
由图4可以看出,粗木质物残体从冰川退缩以后第35年(S2)左右开始出现,其生物量为5.785 Mg·hm-2,随后不断增加到第85年(S4)达到最大值 81.102 Mg·hm-2,而后又有所下降。指数方程y=17.554×exp(x/45.608)-32.035(r2=0.999,P=0.001)可很好地拟合粗木质物残体量从S2到S4的变化曲线。年叶凋落物量的变化随着演替的进行也满足指数 增 长 方 程y=-4.860×exp(-x/20.571)+2.698(r2=0.9832,P=0.031),S4是其增长速率的拐点,在这之前,年叶凋落物量增长迅速,之后年叶凋落物量增长趋于平缓。
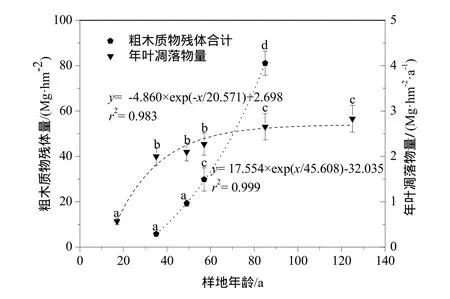
图4 原生演替不同阶段粗木质物残体和年叶凋落物量变化Fig. 4 Biomass changes of coarse woody debris and annual leaves litter amount alonng the chronosequence
3 结论与讨论
3.1讨论
有研究指出,森林生物量变化与许多生物学和非生物学因素密切相关,如区域的水热条件、土壤环境以及森林的类型、年龄、优势种的组成和郁闭度等(黄从德等,2008;杜虎等,2014),而生物量分配特征则主要受到其生活型和生态位的影响(王晓莉等,2014)。海螺沟冰川退缩区群落生物量随演替的进行不断增加,演替初期仅有 10.195 Mg·hm-2,随后不断增长,最终达到 366.121 Mg·hm-2,与地带性植被贡嘎山东坡峨眉冷杉成熟林相近(程根伟等,2003),但要高于世界温带针叶林生物量的平均值 307 Mg·hm-2(Cole et al.,1981)。说明经过了 120多年的演替,海螺沟冰川退缩区生态系统植被生物量已达到成熟林水平,高于欧洲以及北美冰川退缩区的(Borman et al.,1990;Mound et al.,2002)。这可能是因为海螺沟冰川属于季风海洋性冰川,与欧洲、北美等处于高纬度的冰川相比,海螺沟冰川退缩区具有气温高、降水量大的特点,水热组合条件较好,为早期先锋植物的定居提供了良好的环境基础。例如 Chapin et al.(1994)通过对阿拉斯加Glacier Bay冰川原生演替研究发现,先锋植物在冰川退缩第 10年后才开始侵入原生裸地,且最初形成的先锋群落没有乔木幼苗成分,裸地形成的第15至20年才有柳树和赤杨(Alnus rubra)进入先锋群落。而本研究中在冰川退缩后第4年就有种子植物黄芪、柳叶菜等生长发育,并有柳树、冬瓜杨和沙棘幼苗伴生在其周围。
本研究表明,演替不同阶段生物量的绝大部分来自乔木层,其他各层对总生物量贡献较低。这可能是由于不同演替阶段乔木层结构的变化决定了植被可利用的光、热、水和养分等资源,进而制约着林下植被的分布格局和生物量的分配(张昌顺等,2012;肖洒等,2014;史山丹等,2012)。根据演替不同阶段优势种的组成可将演替划分为3个阶段:S1~S4为落叶阔叶林阶段,S4~S5为阔叶到针叶林过渡阶段,S5~S6为常绿针叶林阶段。随着演替的进行,乔木层生物量不断增加,总体表现出云、冷杉成熟林(S6)>冬瓜杨、云冷杉混交林(S5)>冬瓜杨成熟林(S4)>柳树、沙棘成熟林(S3),而乔木层生物量所占比例最高值则出现在样地S2,主要是因为此时林分密度比较大,群落的郁闭度也在整个演替序列上最高,达0.947(表2),林下植物生长空间较小,因此生物量所占比例较低(肖洒等,2014)。从S2到S4,由于种内竞争与种间竞争的加剧,出现大量林窗,群落郁闭度开始下降,有利于灌木、草本以及地被植物的生长,林下各层生物量所占比例逐渐提高。到了 S5以后,群落郁闭度再一次上升到一个较高值,空气湿度也达到了整个演替序列最高水平,为苔藓、石松等地被植物的生长提供了有利的环境条件。最后随着群落演替到针叶林阶段以后,由于云冷杉的自疏作用,再次出现了大量林窗,群落郁闭度降低,促进了灌木层以及草本层生物量的进一步增长。
乔木层生物量分配特征的变化是不同演替阶段树种相互替代的结果。从生态位理论和竞争排斥原理相结合的角度来看,演替总是朝着适应性强以及生态幅大的物种的方向演进(常丽等,2013)。Whittaker也指出(演替过程中)“1个优势种对土壤和气候的改变为另外 1个种的侵入提供了可能性,新侵入的种变为优势种以后,继续对环境进行改变,使环境抑制第一个种而为第3个种的侵入提供可能,第 3个种的侵入又继续改变环境”(Whittaker,1993;郑晓翾等,2007)。本研究中虽然早期原生裸地的环境条件比较恶劣,土壤养分贫瘠,但是相对于物种数量来说,资源还算比较丰富,因此柳树、沙棘和冬瓜杨仍能够共同生长,物种之间的关系以种内竞争为主(S1~S2),产生的粗木质物残体量及年叶凋落物量也相对较少。相对来说,柳树和沙棘比冬瓜杨更能适应原生裸地缺氮的环境,生长较好,所以演替初期柳树、沙棘和冬瓜杨生物量之比为12∶7∶4,以柳树为主。随后一段时期,柳树和沙棘的固氮作用使得土壤N含量迅速增加,为冬瓜杨的生长提供了充足的养分,冬瓜杨凭借其快速的高生长速度,迅速成长,生态位得到进一步拓展,其生物量占乔木层的比例也由 S1的17.15%增至S4的87.31%(图3c),沙棘和柳树则在种间竞争中败下阵来,相继退出群落,导致粗木质物残体量增长迅速。到了S4,群落进入到冬瓜杨大树阶段,冬瓜杨由于冠幅较大,占据大部分上层空间,群落郁闭度明显增加,不利于冬瓜杨种子的萌发,却为喜荫的云冷杉幼苗的生长提供了绝佳的机会,冬瓜杨开始慢慢被云冷杉所取代,逐步退出群落。在针阔混交林阶段(S5),粗木质物残体量也达到了整个演替序列的最大值。另外,由于演替前期阔叶树种年叶凋落物量较高且分解较快,土壤中有机质含量达到整个演替序列的最高值,加速了云冷杉幼苗的生长发育,其生物量也由S4的0.294 Mg·hm-2一跃增至S5的268.001 Mg·hm-2,逐渐进入主林层。经过125 a的演替之后,云冷杉已完全取代冬瓜杨成为新的优势种,生态系统结构与功能相对稳定,植被演替发展到顶级群落(张继义等,2003)。
3.2结论
海螺沟冰川退缩区域植被演替是群落更替的过程,表现为群落结构和功能及其环境的变化是一个有序的、可以观测的连续过程。总的来说,海螺沟冰川退缩区活植物体生物量随演替的进行不断增加,从先锋群落的10.195 Mg·hm-2不断增长到顶级群落的366.121 Mg·hm-2,接近于地带性植被贡嘎山东坡峨眉冷杉成熟林(程根伟等,2003),但要高于世界温带针叶林生物量的平均值 307 Mg·hm-2(Cole et al.,1981)。粗木质物残体量和年叶凋落物量也随着演替的进行不断积累,其中粗木质物残体量在针阔混交林阶段(S5)达到最高,年叶凋落物量则随演替的进行呈指数增加的趋势。
不同演替阶段群落生物量的分配特征也不尽相同。就不同层次而言,不同演替阶段乔木层生物量在各个层次分配中占绝对优势(>89.871%),其他层次所占比例较小,总体表现为:灌木层>地被层>草本层,林下各层生物量分配受到群落环境影响较大。就各个树种而言,演替前期,群落生物量主要来自柳树、沙棘和冬瓜杨等阔叶树种,后期云冷杉等针叶树种对群落生物量贡献较大。就各个器官而言,乔木层生物量的器官分配以树干所占比例最高,为56.388%~72.658%,枝和根的比例次之,叶所占比例则最小。
BORMAN B T, AIDLE R C.1990. Changes in Productivity and Distribution of Nutrients in a Chronosequence at Glacier Bay National Park, Alaska [J]. Journal of Ecology, 78(3): 561-578.
CHAPIN F S, SHARMAN L C. 1994. Mechanisms of Primary Succession Following Deglaciation at Glacier Bay, Alaska [J]. Ecological Monographs, 64(64): 149-175.
COLE D W, RAPP M. 1981. Elemental cycling in forest ecosystems. In: Reichle DE (Ed) Dynamic Properties of Forest Ecosystems [M]. Cambridge University Press, New York.
COWLES H C. 1899. The ecological relations of the vegetation on the sand dunes of Lake Michigan [M]. University of Chicago Press.
CROCKER R L, MAJOR J. 1955. Soil Development in Relation to Vegetation and Surface Age at Glacier Bay, Alaska [J]. Journal of Ecology , 43(2): 427-448.
FASTIE C L. 1995. Causes and ecosystem consequences of multiple pathways of primary succession at Glacier Bay, Alaska [J]. Ecology, 76(6): 1899-1916.
HOBBIE E A, MACKO S A, SHUGART H H. 1998. Patterns in N dynamics and N isotopes during primary succession in Glacier Bay, Alaska [J]. Chemical Geology, 152(1): 3-11.
HODKINSON I D, COULSON S J. 2002. Primary community assembly on land-the missing stages: why are the heterotrophic organisms always there first? [J]. Journal of Ecology, 90(3): 569-577.
HOSHIZAKI K, NIIYAMA K, KIMURA K, et al. 2004. Temporal and spatial variation of forest biomass in relation to stand dynamics in a mature, lowland tropical rainforest, Malaysia [J]. Ecological Research, 19(3): 357-363.
HOUGHTON R A. 2005. Aboveground Forest Biomass and the Global Carbon Balance [J]. Global Change Biology, 11(6): 945-958.
LUO T X, SHI P L, LUO J, et al. 2002. DISTRIBUTION PATTERNS OF ABOVEGROUND BIOMASS IN TIBETAN ALPINE VEGETATION TRANSECTS [J]. Acta Phytoecologica Sinica, 26(6): 668-676.
JOHNSON E A, MIYANISHI K. 2008. Testing the assumptions of chronosequences in succession [J]. Ecology Letters, 11(5): 419-431.
JONES G A, HENRY G H R. 2003. Primary plant succession on recently deglaciated terrain in the C--anadian High Arctic [J]. Journal of Biogeography, 30(2): 277-296.
KIMMIS J P. 1987. Forest ecology [M]. New York: Macmilan: 407-409.
MAUN A M P M A. 1981. Dispersal and floating ability of dimorphic fruit segments of Cakile edentula var. lacustris [J]. Canadian Journal of Botany, 59(12): 2595-2602.
MOUND M, KUMMETZ E, HEIN M, et al. 2002. Growth and carbon stocks of a spruce forest chronosequence in central Europe [J]. Forest Ecology & Management, 171(3): 275-296.
OHTONEN R, FRITZE H, PENNANEN T, et al. 1999. Ecosystem properties and microbial community changes in primary succession on a glacier forefront [J]. Oecologia, 119(2): 239-246.
OLSON J S. 1958. Rates of Succession and Soil Changes on Southern Lake Michigan Sand Dunes [J]. Botanical Gazette, 119(3): 125-170.
PICKETT S A.1989. Space-for-time substitution as an alternative to long-term studies[M]. Likens G E, ed. New York: Berlin Heidelberg, 110-135.
COLLINS S L, ADAMS D E. 1983. Succession in grasslands: Thirty-two years of change in a central Oklahoma tallgrass prairie [J]. Vegetation, 51(3): 181-190.
OVERMAN J P M, WITTE H J L,SALDARRIAGA J G. 1994. Evaluation of regression models for above-ground biomass determination in Amazon rainforest [J]. Journa lof tropical Ecology, (10): 207-218.
SAATCHI S S, HOUGHTON R A, DOS S A, et al. 2007. Distribution ofaboveground live biomass in the Amazon basin [J]. Global Change Biology, 13(4): 816-837.
VETAAS O R. 1994. Primary Succession of Plant Assemblages on a Glacier Foreland-Bodalsbreen, Southern Norway [J]. Journal of Biogeography, 21(3): 297-308..
WHITTAKER R J. 1993. Plant population patterns in a glacier foreland succession: pioneer herbs and later-colonizing shrubs [J]. Ecography, 16(2): 117-136.
WIEGLEB G, FELINKS B. 2001. Primary succession in post-mining landscapes of Lower Lusatia - chance or necessity [J]. Ecological Engineering, 17(2-3): 199-217.
LI ZONG X, HE Y Q, PANG H X, et al. 2008. Source of major anions and cations of snowpacks in Hailuogou No.1 glacier, Mt. Gongga and Baishui No.1 glacier, Mt. Yulong [J]. Journal of Geographical Sciences, 18(1): 115-125.
常丽, 何元庆, 杨太保, 等. 2013. 玉龙雪山白水1号冰川退缩迹地的植被演替[J]. 生态学报, 33(8): 2463-2473.
陈富斌, 罗辑. 1998. 贡嘎山高山生态环境研究 [M]. 第2卷. 北京: 气象出版社.
程根伟, 余新晓, 赵玉涛, 等. 2003. 贡嘎山亚高山森林带蒸散特征模拟研究[J]. 北京林业大学学报, 25(1): 23-27.
杜虎, 曾馥平, 王克林, 等. 2014. 中国南方3种主要人工林生物量和生产力的动态变化[J]. 生态学报, 34(10): 2712-2724.
冯宗炜. 1999. 中国森林生态系统的生物量和生产力[M]. 科学出版社.
何磊, 唐亚. 2007. 海螺沟冰川退化迹地土壤序列的发育速率[J]. 西南大学学报: 自然科学版, 29(7): 139-145.
黄从德, 张健, 杨万勤, 等. 2008. 四川省及重庆地区森林植被碳储量动态[J]. 生态学报, 28(3): 966-975.
霍常富, 程根伟, 鲁旭阳, 等. 2010. 气候变化对贡嘎山森林原生演替影响的模拟研究[J]. 北京林业大学学报, 32(1): 1-6.
李登秋, 居为民, 郑光, 等. 2013. 基于生态过程模型和森林清查数据的森林生长量估算对比研究[J]. 生态环境学报, 22(10): 1647-1657.
李逊, 熊尚发. 1995. 贡嘎山海螺沟冰川退却迹地植被原生演替[J]. 山地学报, 13(2): 109-115.
李宗省, 何元庆, 贾文雄, 等. 2009. 全球变暖背景下海螺沟冰川近百年的变化[J]. 冰川冻土, 31(1): 75-81.
刘兴良, 马钦彦, 杨冬生, 等. 2006. 川西山地主要人工林种群根系生物量与生产力[J]. 生态学报, 26(2): 542-551.
罗辑. 1996. 贡嘎山东坡植被原生演替的种间协变[J]. 山地学报, 14(4): 235-238.
史山丹, 赵鹏武, 周梅, 等. 2012. 大兴安岭南部温带山杨天然次生林不同生长阶段生物量及碳储量[J]. 生态环境学报, 21(3): 428-433.
王晓莉, 常禹, 陈宏伟, 等. 2014. 黑龙江省大兴安岭主要森林生态系统生物量分配特征[J]. 生态学杂志, 33(6): 1437-1444.
宿以明, 刘兴良, 向成华. 2000. 峨眉冷杉人工林分生物量和生产力研究[J]. 四川林业科技, (2): 31-35.
肖洒, 吴福忠, 杨万勤, 等. 2014. 川西高山森林生态系统林下生物量及其随林窗的变化特征[J]. 生态环境学报, 23(9): 1515-1519.
许中旗, 李文华, 鲍维楷, 等.2005. 植被原生演替研究进展[J]. 生态学报, 25(12): 3383-3389.
鄢武先, 宿以明. 1991. 云杉人工林生物量和生产力的研究[J]. 四川林业科技, 12(4): 17-22.
张昌顺, 谢高地, 陈龙等. 2012. 地形和乔木结构对热带北缘与南亚热带过渡带林下植被分布的影响[J]. 资源科学, 34(7): 565-566.
张继义, 赵哈林. 2003. 植被(植物群落)稳定性研究评述[J]. 生态学杂志, 22(4): 42-48.
郑晓翾, 赵家明, 张玉刚, 等. 2007. 呼伦贝尔草原生物量变化及其与环境因子的关系[J]. 生态学杂志, 26(4): 533-538.
钟祥浩, 罗辑, 吴宁. 1997. 贡嘎山森林生态系统研究[M]. 成都: 成都科技大学出版社.
朱兴武, 肖瑜, 蔡文成. 1988. 山杨天然次生林生物量的初步研究[J].青海农林科技, (1): 30-34.
Dynamics of Vegetation Biomass Along the Chronosequence in Hailuogou Glacier Retreated Area, Mt. Gongga
YANG Dandan1,2, LUO Ji1*, SHE Jia3, TANG Ronggui1
1. Insitute of Mountain Hazards and Environment, Chinese Academy of Science, Chengdu 610041, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Sichuan Academy of Environmental Sciences, Chengdu 610031, China
In order to better understand the vegetation biomass changes and distribution characteristics of different components along time, seven sampling plots (S0~S7) representing different succession stages (bare land, 17 years, 35 years, 49 years,56 years,85 years and 125 years) were chosen through investigation of the 125-year-old chronosequence in Hailuogou glacier retreated area, Mt. Gongga. Results showed that stand age and plantation types were two main factors leading to biomass change. Total living biomass of the vegetation and biomass of the tree layer both presented an exponential growth pattern along time, separately ranged from 10.195 Mg·hm-2to 366.122 Mg·hm-2, from 9.162 Mg·hm-2to 332.461 Mg·hm-2. Tree layer contributed most to the total living biomass, which accounted for more than 89.871 percent during all stages; in contrast, biomass of the other layers only had little influence on the total biomass and showed a tendency of shrub layer > ground cover > the herb layer. What’s more, these underwood layers were more affected by the community environment. Biomass of the coarse woody debris and annual leaves litter also accumulated during the succession chronosequence, biomass of the coarse woody debris increase to the peak at S5, where the mixed broadleaf-conifer forest was growing while the biomass of the annual leaves litter fitted well with the exponential growth model during the whole succession chronosequence. In the former 60 years of succession, biomass of the tree layer mainly came from deciduous broad-leaf species such as Salix rehderana, Hippophae rhamnoides and Populus purdomii, in the last 60 years of succession, evergreen coniferous trees, for instance, Abies fabri and Picea brachytyla contributed most to the total layer biomass (>93.070%). As for biomass allocation related to various organs, the trunk occupied the greatest proportion of the tree layer, which made up 56.388%~72.658% of the total biomass,branch and roots came secondly, while leaves took up the least proportion among all the organs.
Hailuogou; glacier retreated area; primary succession; biomass
10.16258/j.cnki.1674-5906.2015.11.014
Q948;X171.1
A
1674-5906(2015)11-1843-08
国家自然科学基金项目(41471416;41272200;40871042)
杨丹丹(1992年生),女,硕士研究生,研究方向为植被生态与环境。E-mail: fydiane@163.com *通信作者:罗辑(1960年生),男,研究员。E-mail: luoji@imde.ac.cn
2015-09-08
引用格式:杨丹丹, 罗辑, 佘佳, 唐荣贵. 贡嘎山海螺沟冰川退缩区原生演替序列植被生物量动态[J]. 生态环境学报, 2015, 24(11): 1843-1850.
