桑园地和玉米轮作地土壤pH变化的比较研究
2015-12-07凌宏文樊宇红朴河春
凌宏文,樊宇红,朴河春
中国科学院地球化学研究所环境地球化学国家重点实验室,贵州 贵阳 550002
桑园地和玉米轮作地土壤pH变化的比较研究
凌宏文,樊宇红,朴河春*
中国科学院地球化学研究所环境地球化学国家重点实验室,贵州 贵阳 550002
随着桑园植桑年龄的增加,桑地土壤质量降低,桑树生物量减少,严重制约着养蚕业的可持续发展。这可能与土壤pH降低、营养元素的流失和有害元素的积累有关。选取广西宜州和贵州荔波的桑树(Morus alba)种植园和临近种玉米(Zea mays)轮作地的土壤,分析土壤可交换性盐基阳离子(Ca,Mg,K和Na),可交换性铝(Al)、土壤pH等土壤参数,研究土壤pH在桑树地和玉米轮作地之间的差异,讨论影响土壤pH变化的因素。实验结果表明,相对于玉米轮作地,桑园地土壤pH 5年下降了0.38个pH单位。相应地,桑园地土壤可交换性盐基阳离子(Ca,Mg和K)的含量略低于玉米轮作地,桑地土壤交换性Al的含量(2.35±2.68) mmol·kg-1却显著高于玉米轮作地(1.44±2.13) mmol·kg-1(P=0.002)。因此,桑园地和玉米轮作地土壤可交换性Ca和Al之间呈现出显著的负相关关系。桑园地土壤可交换性NH4+-N含量略高于玉米轮作地。得出结论:随着桑园经营年代的增加,土壤酸化的同时,营养元素(Ca,Mg和K)的流失和有害元素(Al)的积累,土壤质量下降。玉米轮作缓解土壤酸化。显然,不同的经营模式要影响土壤pH变化。导致土壤pH变化的主导因素是碳和氮的循环。在农业生产过程中,我们应该提倡不同作物轮作,以适当的方式将秸秆还田,以保持良好的土地质量。
土壤pH;交换性盐基阳离子;交换性Al离子;桑园;玉米;轮作
广西河池市是我国第一大蚕桑生产基地,2012年桑园面积4.84万hm2(张建华等,2013),该地区种植桑树的产区相对集中,已形成传统的优势养蚕基地,常常多年连片、大规模种植。由于每年剪伐和一年中的多次用叶,桑树需要从土壤中吸收大量的营养物质。随着种植年限的增加,桑园土地生产潜力降低,桑树生产性能退化,严重制约养蚕业的可持续发展。为提高桑园生产力,提高化肥和农药的用量,不仅引起环境问题,还要缩短桑园的寿命。
土壤酸化是单一物种连续种植所带来的共同结果。比如,蔬菜(黄瓜和西红柿)连作后土壤pH呈现降低趋势(张作新和张树明,2015);草莓连作引起土壤酸化(赵海涛等,2014);随着参园年限的增加,参地土壤pH不同程度下降(徐厚来,2014);大豆连作导致土壤速效养分和pH下降(周丽萍等,2010)。然而,有研究证明,轮作换茬或套作可能是克服桑园土地退化的经济有效的措施之一。比如,利用不同作物轮作和套作,以及增施生物有机肥可改善土壤酸化状况(肖新等,2015);水旱轮作(丝瓜和豆瓣菜)明显缓解连作所带来的土壤酸化(袁建玉等,2014);黄瓜-玉米轮作也明显缓解设施连作土壤的酸化(唐艳领等,2014)。
植物通过一系列的机制,来改变土壤的物理、化学和生物的性质,如植物根系释放质子可酸化土壤(Richardson等,2009;樊宇红等,2014)。土壤pH是高度敏感性因子,它决定植物的生存和分布。土壤pH的改变将直接影响营养物的可利用性,或通过与土壤微生物的相互作用,来间接地影响营养物的可利用性(Richardson等,2009;Muthukumar等,2014)。土壤酸化是世界决大多数地区所面临的严重的农业问题,它影响世界大约40%的可耕地(Pinheiro等,2004;Rukshana等,2012)。我国土壤 pH变化整体表现为酸性化趋势(王志刚等,2008;郭治兴等 2011)。我国主要农田土壤 pH在20年的时间里平均下降约0.5个单位。其中,经济作物体系土壤酸化比粮食作物体系更为严重(杨世琪等,2010;Guo等,2010)。茶树种植引起的土壤酸化可看作自然和人为过程的综合作用(Wang等,2010)。
影响土壤pH变化的因素较多,其中,(1)碳(C)的循环主导着土壤pH变化的主要过程(Wang等,2010)。C转换所引起的pH值增加是通过H+离子与有机组分的结合,以及在植物残留物中的有机酸的脱羧基作用所实现的。H+与有机化合物的结合或分离的反应在植物残留物表面和土壤基质上进行,并贡献于土壤pH的变化(Wang等,2014)。因此,在植物残留物中的有机阴离子的含量,是影响土壤pH变化的主要因素之一(Tang和Yu,1999)。(2)通过N循环影响土壤pH变化。比如,有机N矿化成氨的作用能增加土壤pH值,而硝化作用降低土壤pH值(Limousin和Tessier,2007;Butterly等,2013)。因此,在植物残留物中的有机和无机N含量,是影响土壤pH变化的主要因素之一(Tang和Yu,1999)。(3)植物生长本身能够酸化或碱化土壤环境(Moody和 Aitken,1997)。比如,植物对铵的吸收能降低土壤pH(Limousin和Tessier,2007;Butterly等,2013);植物对NO3-的吸收能增加土壤pH值(Limousin和Tessier,2007;Butterly等,2013);豆科类植物的固氮作用降低土壤 pH(Tang等,1999)。植物根系所释放的分泌物(Limousin和Tessier,2007),植物对阳离子的吸收都要影响土壤pH(Muthukumar等,2014)。
土壤溶液中H+离子含量的增加引起土壤pH的降低,pH<5.5或更低的土壤归纳为酸性土壤(Muthukumar等,2014)。土壤酸化使土壤可交换性酸增加,而土壤中可交换性盐基阳离子含量减少,阳离子交换能力减弱。土壤pH的变化对重金属元素在土壤中的分配模式产生重要影响(凌宏文等,2007)。种植茶树所引起的土壤酸化也可导致土壤中可交换性Al和溶解性Al的增加(Wang等,2010)。土壤溶液pH的变化是影响微生物生理过程的直接原因,如土壤pH下降引起微生物较低的分解代谢速度(Clarholm和Skyllberg,2013)。然而,植物残留物对土壤pH变化的影响在文献中出现不同的结果,这是因为残留物组成和所使用的土壤的性质差异所致。特别是土壤的初始 pH对土壤 pH变化有重要影响(Butterly等,2013)。因此,有机物加入土壤可导致土壤pH的增加、降低、或保持不变(Rukshana等,2011)。茶树种植引起的土壤酸化(Wang等,2010),杉木的连载也引起生物量的降低(Piao和Liu,2011;Piao等,2012)。随着桑园植桑年龄的增加,与其他作物连作一样,是否有土壤酸化的倾向?本研究比较桑园地和玉米轮作地土壤pH的差异,讨论影响土壤pH变化的因素,研究结果对合理经营桑树种植园有指导意义。
1 研究地概况与研究方法
1.1 研究区概况
本研究所使用的土壤采自广西河池市宜州和贵州荔波。宜州位于广西中部偏北的位置。宜州气候属亚热带季风气候区。采样地位于东经108°07′11″~108°40′62″、北纬24°17′74″~24°40′95″之间,海拔高度在 110~285 m,年平均气温 19.6~20.2 ℃,年平均降雨量1300~1500 mm。土壤类型为红壤,其成土母质主要有残积物、堆积物、冲积-洪积物、沉积物等(杨艳芳等,2009)。荔波位于贵州南部,珠江流域上游,处于贵州高原向广西丘陵过度地带。采样地位于东经 108°00′50″~108°10′37″、北纬25°17′26″~25°29′51″之间,海拔高度500~720 m,年平均气温18.3 ℃,年平均降雨量1320.5 mm。荔波采样地多为碳酸盐盐类,但与砂岩交叉分布,所采集的土壤类型均为黄壤(樊宇红等,2014)。
荔波的桑树是从广西引进的。平均株行距大约0.5 m×1.2 m,5年生桑树的平均根径为5 cm左右。桑树<1 mm细根在0~20 cm土层内分布密度最大,表现出水平状的分布,深部的根系表现出垂直状的分布(Liu和Willison,2013;石娟华等,2008),说明桑树细根不但能利用表层土壤空间,还更能吸收利用深层土壤水分和养分(Liu和Willison,2013;石娟华等,2008)。桑树地下部分的生物量明显高于地上部分,地下部分的生物量达总生物量的53.1%(张光灿等,1997)。以养蚕为栽培目的的桑树,从5月上旬开始摘叶,5月底伐去全部枝叶,继续萌发枝叶后,从7月初至10月底一直处于摘叶养蚕中。考虑到土壤性质的不均匀性,以及不同母岩对土壤性质的影响,玉米轮作地选取是紧靠近桑地。桑地的施肥采用春、夏、冬3次施肥的方法,其使用量和次数并不一致。至于玉米轮作地,近一两年种植过玉米,之前的情况较为复杂,有的种植甘蔗、花生等其他作物,每年有3~6个月的休闲。施肥情况也不一致,有的采用常规分次施复合肥和尿素,有的采用一次性施肥,种植甘蔗有时不施肥。每个样点待测土壤由同一地块 8个随机点位 0~10 cm的表层土样混合构成。采样是在2014年5月末进行的。
1.2 研究方法
1.2.1 样品采集和分析
采集到的土壤样品用2 mm的筛子过筛、风干,并剔除植物残留物。然后取部分均匀的土壤样品研磨至粉状,过0.149 mm筛子,用于C和N含量的测定,所使用的仪器为元素分析仪(Vario MACRO,德国)。土壤可交换性阳离子用1 mol·L-1醋酸铵溶液提取,并用等离子发射光谱测定(ICP-OES-美国安捷伦公司)(Lindsay和Norvell,1978);可交换性Al和NH4+-N用1 mol·L-1的KCl溶液提取,可交换Al含量用等离子发射光谱测定,NH4-N含量用纳氏试剂比色法测定。Olsen P(速效P)用0.5 mol·L-1NaHCO3(pH=8.5,用NaOH溶液来调)溶液提取,用钼兰比色法测定。在样品分析中平行样的测定达20%,所得分析数据均符合要求。所有的实验在中国科学院地球化学研究所环境地球化学国家重点实验室里完成。
1.2.2 统计方法
采用SPSS 17.0对数据进行处理。在文中所表示的数据为平均值±标准偏差。在一般情况下,所测量的数据在5%的概率下进行线行回归和t检验,来确定其显著性差异。在此,我们所采取的是所测量的数据在 10%的概率下确定其显著性差异(Anderson等,1995)。
2 结果
2.1 桑地和玉米轮作地土壤pH对比
在桑地里,桑树的种植年龄约5年时间。与临近的玉米轮作地土壤pH相比,桑地土壤pH明显下降(表1),其下降幅度为0.38 pH单位。
2.2 桑地和玉米轮作地土壤中C、N和P含量对比
表1列举了桑地和玉米轮作地土壤中有机C、全N和全P的含量。虽然在统计学意义上有机C、全N和全P在桑地和玉米轮作地之间没有显著性差异,在桑地土壤中它们的含量略高于玉米轮作地(表 1)。桑地土壤可交换性 NH4+-N 的含量(4.23±0.97) mmol·kg-1也略高于玉米轮作地(4.02±0.65) mmol·kg-1(P=0.176)。土壤可交换性NH4+-N 的含量随土壤 pH 的增加而在桑地(r2=0.193)和玉米轮作地(r2=0.223)均在减少。桑地土壤速效 P(Olsen P)的含量为(1.88±1.28) mmol·kg-1,而玉米轮作地为(1.68±1.01) mmol·kg-1,但没有统计学意义上的差异(P=0.410)。显然,土壤中NH4+-N可在酸性条件下积累。
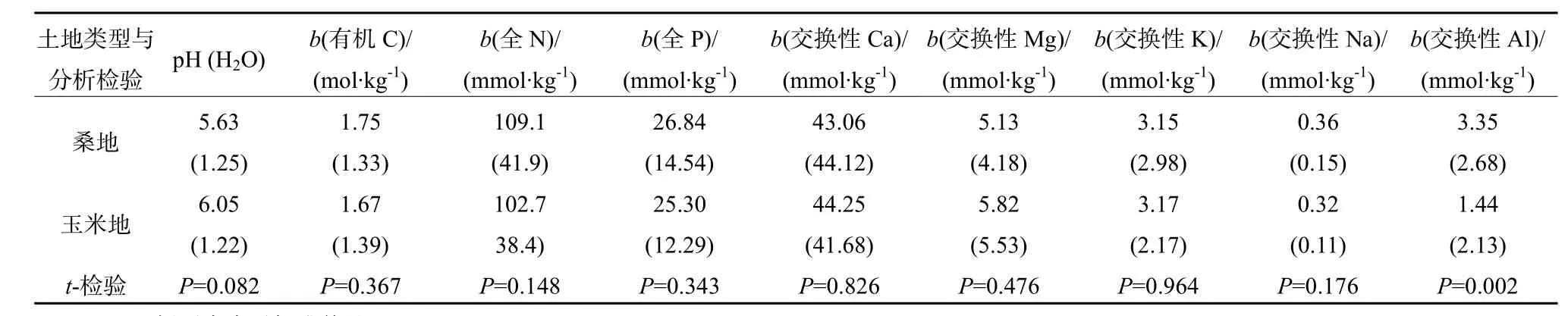
表1 桑地和玉米轮作地土壤pH与有机C、全N、全P、可交换性盐基阳离子的平均含量Table 1 Soil pH, and the contents of organic C, total N, total p, soil exchangeable base cation and Al
2.3 桑地和玉米轮作地土壤可交换性盐基阳离子(Ca,Mg,K和Na)含量的变化
虽然土壤可交换性Ca,Mg和K的含量在桑地和玉米轮作地之间没有显著性差异,在桑地中的含量略低于玉米轮作地。但桑地土壤可交换性Na的含量略高于玉米轮作地(表1)。桑地土壤可交换性Ca含量与土壤pH之间有显著的正相关关系(图1,r2=0.833)。同样,玉米轮作地土壤可交换性 Ca与土壤 pH之间也有显著的正相关关系(图 1,r2=0.761)。然而,其它盐基阳离子和土壤pH之间的相关性没有那么显著。比如,桑地土壤可交换性Mg与土壤pH(r2=0.404),K与pH(r2=0.074),Na与pH(r2=0.082)之间的相关性都要低于Ca与pH之间的相关性;同样,玉米轮作地土壤可交换性Mg与土壤pH(r2=0.329),K与pH(r2=0.146),Na与pH(r2=0.148)之间的相关性也低于Ca与pH之间的相关性。

图1 桑地和玉米轮作地土壤可交换性Ca含量与土壤pH的变化关系Fig. 1 Relationship between soil exchangeable Ca concentrations and soil pH
2.4 桑地和玉米轮作地土壤可交换性Al含量变化
桑地土壤可交换性 Al的平均含量显著高于玉米轮作地(表1)。它们的含量随土壤pH的增加而以指数形式降低(图 2):桑地为 y=1175.3e-1.2866x(r2=0.840); 玉米 轮 作地 为 y=722.6e-1.2221x(r2=0.730)。桑地和玉米轮作地土壤可交换性Ca和Al之间呈现出负的相关关系(图 3)。桑地土壤可交换性Al含量随土壤可交换性Ca含量的增加而以指数形式降低(图3,y=3.3484e-0.0322x,r2=0.657),玉米轮作地土壤可交换性 Al的含量也随土壤可交换性 Ca含量的增加而以指数形式降低(图 3,y=1.4519e-0.0266x,r2=0.405)。

图2 土壤可交换性Al含量与土壤pH的变化关系Fig. 2 Relationship between soil exchangeable Al and soil pH

图3 桑地和玉米轮作地土壤可交换性Ca含量和土壤可交换性Al含量之间的相关关系Fig. 3 The relationship between soil exchangeable Ca concentrations and soil exchangeable Al concentrations in mulberry and maize rotation soils
2.5 Ca/Al比值指示土壤酸化
在本研究中,土壤可交换性Ca/Al的mol比值在桑地(420±896)和玉米轮作地(564±1091)的之间没有统计学意义上的差异(P=0.527)。本研究中只出现一处Ca/Al比值小于1的情形。Ca/Al的mol比值小于10的,在桑地出现12个地点,在玉米轮作地出现9个地点。Ca/Al的mol比值在碱性条件下大于酸性条件(图4),是因为碱性条件下,交换性Ca含量大,而Al含量小的缘故。

图4 土壤交换性Ca/Al比值(Log值)与土壤pH变化的相关关系Fig. 4 The relationship of change of soil exchangeable Ca/Al ratios (Log values) with soil pH
3 讨论
实验结果表明,桑园的持续经营导致土壤 pH显著下降,必然导致营养元素的流失和有害元素的积累。比其玉米轮作地,桑地土壤pH的下降是由多种因素造成,其中C和N循环是导致土壤pH变化的主导因素。
3.1 植物残留物对土壤pH的影响
目前大多数地区,经济作物和农作物连带它们的秸秆被移走,带不走的调落物就地燃烧处理。相比之下,桑地里留下的残留物数量要少,因为绝大部分桑叶为养蚕被摘走的缘故。作物残留物的加入所诱导的土壤pH的变化,主要是通过H+离子的结合/分离的反应,以及 C和 N循环过程来实现(Rukshana等,2012)。农作物残留物中含有大量的盐基阳离子(Ca,Mg,K和Na),与之相平衡的主要是有机阴离子。植物残留物在微生物的分解作用下给土壤补充大量的盐基阳离子(Rukshana等,2012;Xiao等,2014),其结果是增加土壤pH。比如,巢菜(vetch)残留物的加入可引起较大的土壤pH的增加(Rukshana等,2012),但土壤初始pH较大时,它没有恒定的增加,是因为硝化作用的缘故(Xiao等,2014)。在低pH条件下矿物质N是以 NH4+-N的形式存在于土壤中,而在较高的 pH条件下,它是以NO3--N的形式存在于土壤中(Xiao等,2014)。在茶树种植园土壤中大量的盐基阳离子以茶叶的形式带走。残留物分解期间所释放的碱度不足于抵消在这些土壤中的较高的交换性酸度,因此,茶园土壤酸化较为严重(Wang等,2014),与本研究结果相一致。因此,作物残留物被返还给土壤是保持土壤肥力的一种可行的策略,可增加土壤有机质的积累;作物残留物的保留对土壤碱度的重新分布具有重要意义(Butterly等,2013)。
3.2 免耕对土壤pH变化的影响
桑园是基本不犁地,而玉米轮作地是犁地。未犁地的土壤显示强烈的pH剃度变化,其表层5 cm的土壤pH最低。Limousin和Tessier(2007)把未犁地土壤pH下降的原因归结为:(1)有机质的矿化作用;(2)所施入的铵态氮肥的硝化作用在土壤表层引起酸性效应;(3)根系的分泌作用,因为植物根系主要分布在土壤表层的缘故。在耕作系统中,可交换性Al的含量显著地与pH相关(Limousin和Tessier,2007)。
3.3 施肥对土壤pH的影响
桑园地和玉米轮作地均施用大量的复合肥和尿素来增加粮食和经济作物产量。在肥料中的化学物质与土壤矿物营养物相互作用改变了土壤 pH,这将间接地影响植物的生长和健康(Muthukumar等,2014)。佟德利等人(2012)的实验表明,尿素的使用显著增加土壤硝化速率,从而降低土壤pH。在农业生态系统中,铵态氮肥的施入、随后的农作物的吸收、农作物产物移走、淋溶所引起的N的损失,和硝酸盐的反硝化都能引起土壤的酸化。而硝酸盐肥料的施入引起土壤pH的增加(Rukshana等,2012)。赵晶等人(2010)的实验表明所有氮肥的处理均降低了土壤pH值,其中尿素和氯化铵处理使土壤pH降低最多,磷肥处理均引起土壤pH的小幅降低。虽然统计学上没有差异,桑地土壤NH4+-N含量高于玉米轮作地,显然是土壤酸化积累土壤 NH4+-N,是由于在酸性条件下抑制硝化作用的缘故(Xiao等,2014)。
3.4 植物的种植对土壤pH的影响
对照裸露地,松树种植园土壤的pH明显低0.3个单位(Liao等,2012)。豆科植物的种植就能酸化土壤,主要是因为豆科植物比非豆科吸收更多的阳离子的缘故(Muthukumar等,2014)。马尾松的种植导致土壤pH下降,其下降幅度与土壤初始pH相关,初始pH相对高的土壤,其下降幅度更大(杨平平等,2012)。在盐基阳离子-营养物缺乏的条件下,生长良好的维管植物就能增加阳离子-营养物的可利用性,是通过释放碳酸和低分子量的有机酸来促进矿物风化(Shi等,2014)。植物所诱导的矿物风化作用依赖于地质背景和生态系统状态(Shi等,2014)。植物释放各种低分子量羧酸,包括苹果酸,柠檬酸,草酸,乙酸等到根际(Rukshana等,2012),这些有机酸的释放能促进盐基阳离子的释放,并引起激发效应。这种激发效应可能是由于促进微生物的NO3-的吸收,氨化作用,以及土壤有机物分解的缘故,但低pH就抑制这种激发效应(Rukshana等,2012)。
3.5 Ca/Al比值指示土壤pH的变化
在森林生态系统中维持一定的可利用性的 Ca含量对森林健康和保持生产力很重要(Lawrence等,1997)。Limousin和Tessier(2007)的研究结果表明,交换性Ca的含量与pH高度相关(r2=0.95),与我们的结果高度一致(r2=0.78)。考虑到Ca和pH的高度相关性,未犁地土壤的酸化作用将伴随着0~5 cm 土壤层中的约 50%交换性 Ca的流失(Limousin和Tessier,2007)。Ca和Mg是性质相近的盐基阳离子。但可交换性Mg的行为(与pH的相关性 r2=0.36)与交换性 Ca的行为不同,因为Mg的分布随土壤深度增加而增加的缘故(Limousin和 Tessier,2007)。这种不平衡的行为应归结于镁是包含在黏土矿物的结构中,而Ca不存在类质同象现象(Limousin和Tessier,2007)。土壤的Ca的消耗是一种自然的成土过程(Gruba等,2013)。盐基阳离子的淋溶,最终导致土壤pH的降低,并改变了对Al的缓冲作用(Kleijn等,2008)。这将引起Al和NH4+-N在土壤溶液中的积累(Kleijn等,2008)。各种形态的Al在土壤中的结合和移动要受土壤 pH和溶解性有机碳的控制(Tejnecký等,2010)。
土壤中的 Al主要存在于结晶的铝硅酸盐矿物中,相对地耐分解。然而,Al的部分仍然是较为活性,随着土壤 pH的下降它的可移动性在加强(Tejnecký等,2010),它对有生命有机物造成威胁(Yvanes-Giuliani等,2014)。Al和pH之间有强烈的相关性,土壤 Al含量与土壤 pH有负相关性(Pinheiro de Carvalho等,2004),这与我们的结果一致。土壤交换性 Al与有机质有正相关性,是由于有机物络合 Al的缘故(Limousin和 Tessier,2007)。土壤以可交换的形式储存Ca,但在交换位置中 Al的饱和度的增加而 Ca的含量在减少(Lawrence等,1997)。
通常Ca/Al比值,在土壤水溶液中Ca和Al活性的摩尔比值(Ca/Al比值)是一种指示森林健康条件的重要指示剂,该值小于1表明对森林生长条件来说是一种负面条件(Gruba等,2013)。交换性Ca含量小于4%时,就可出现mol Ca/Al比值小于1(Gruba等,2013)。在本研究中只出现一处小于 1的地点。酸化将引起Ca/Al比值的减少(Roem等,2008;Kwak等,2011)。Lawrence等(1997)指出酸化作用所诱导的 Al的增加,加剧了森林土壤中的 Ca的减少,因此限制了残留物的分解和矿物风化来的Ca的保持(Gruba等,2013)。随着土壤酸化的进程,H+和Al离子(特别是土壤pH在5.5和4.0时)能够在阳离子交换部位上取代 Ca2+离子(Kwak等,2011),引起Ca2+的淋溶,最终减少土壤的Ca2+的可利用性(Kwak等,2011),是土壤质量下降的重要标志之一。
4 结论
随着桑园植桑年龄的增加,与作物的连作一样,土壤有酸化的趋势。相应地,土壤可利用性Ca、Mg和K营养物有流失,而有害元素Al有积累。这些因素都能用来解释桑园土地生产潜力降低,以及桑树生产性能退化的原因。为了从本质上揭示其桑园退化的原因,需要从化感作用、病原菌的增多,以及丛枝菌根真菌在内的土壤微生物区系失衡,以及它们之间的相互关系入手,进行深入研究。
ANDERSON D L, TUOVINEN O H, FABER A, et al. 1995. Use of soil amendments to reduce soluble phosphorus in dairy soils [J]. Ecological Engineering, 5(2-3): 229-246.
BUTTERLY C R, BALDOCK J A, TANG C. 2013. The contribution of crop residues to changes in soil pH field conditions [J]. Plant and Soil, 366(1-2): 185-198.
CLARHOLM M, SKYLLBERG U. 2013. Translocation of metals by trees and fungi regulates pH, soil organic matter turnover and nitrogen availability in acidic forest soils [J]. Soil Biology and Biochemistry, 63: 142-153.
GRUBA P, MULDER J, BROZEK S. 2013. Modelling the pH dependency of dissolved calcium and aluminium in O, A and B horizons of acid forest soils [J]. Geoderma, 206: 85-91.
GUO J H, LIU X J, ZHANG Y, et al. 2010. Significant acidification in major Chinese croplands [J]. Science, 327(5968): 1008-1010.
KLEIJN D, BEKKER R M, BOBBINK R, et al. 2008. In search for key biogeochemical factors affecting plant species persistence in heathland and acidic grasslands: a comparison of common and rare species [J]. Journal of Applied Ecology, 45(2): 680-687.
KWAK J H, LIM S S, CHANG S X, et al. 2011. Potential use of δ13C, δ15N, N concentration, and Ca/Al of Pinus densiflora tree rings in estimating historical precipitation pH [J]. Journal of Soils and Sediments, 11(5): 709-721.
LAWRENCE G B, DAVID M B, BAILEY S W, et al. 1997. Assessment of soil calcium status in red spruce forests in the northeastern United States [J]. Biogeochemistry, 38(1): 19-39.
LIAO C Z, LUO Y Q, FANG C M, et al. 2012. The effects of plantation practice on soil properties based on the comparison between natural and planted forests: a meta-analysis [J]. Global Ecology and Biogeography, 21(3): 318-327.
LIMOUSIN G, TESSIER D. 2007. Effects of no-tillage on chemical gradients and topsoil acidification [J]. Soil and Tillage Research, 92(1-2): 167-174.
LINDSAY W L, NORVELL W A. 1978. Development of a DTPA test for zinc, iron, manganese, and copper [J]. Soil Science Society of America Journal, 42(3): 421-428.
LIU Y, WILLISON J H. 2013. Prospects for cultivating white mulberry (Morus alba) in the drawdown zone of the Three Gorges Reservoir, China [J]. Environmental Science and Pollution Research, 20(10): 7142-7151.
MOODY P W, AITKEN R L. 1997. Soil acidification under some tropical agricultural systems. 1: Rates of acidification and contributing factors [J]. Australian Journal of Soil Research, 35(1):163-174.
MUTHUKUMAR T, PRIYADHARSINI P, UMA E, et al. 2014. Role of arbuscular mycorrhizal fungi in alleviation of acidity stress on plant growth [M]. Miransari (ed.), Use of Microbes for the Alleviation of Soil Stresses, Volume 1, doi:10.1007/978-1-4614-9466-9_3, Springer Science_Business Media New York, pp43-71.
PIAO H C, LIU C Q, WANG S J. 2012. Isotopic evaluation of the role of arbuscular mycorrhizae in the nitrogen preference in Chinese fir seedlings [J]. Pedobiologia, 55(3): 167-174.
PIAO H C, LIU C Q. 2011. Variations in nitrogen, zinc, and sugar concentrations in Chinese fir seedlings grown on shrubland and plowed soils in response to arbuscular mycorrhizae-mediated process [J]. Biology and Fertility of Soils, 47(6): 721-727.
PINHEIRO DE CARVALHO MÅA, SLASKI J, ABREU I, et al. 2004. Factors contributing to the development of aluminum resistance in the Madeiran maize germplasm [J]. Journal of Plant Nutrition and Soil Science, 167(1): 93-98.
RICHARDSON A E, BAREA J M, MCNEILL A M, et al. 2009. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms [J]. Plant and Soil, 321: 305-339.
RUKSHANA F, BUTTERLY C R, BALDOCK J A, et al. 2011. Model organic compounds differ in their effects oh pH changes of two soils differing in initial pH [J]. Biology and Fertility of Soils, 47(1): 51-62.
RUKSHANA F, BUTTERLY C R, BALDOCK J A, et al. 2012. Model organic compounds differ in priming effects on alkalinity release in soils through carbon and nitrogen mineralization [J]. Soil Biology and Biochemistry, 51: 35-43.
SHI Z Q, BALOGH-BRUNSTAD Z, GRANT M, et al. 2014. Thomashow L, Dohnalkova A, Stacks D, Letourneau M, Keller K. Cation uptake and allocation by red pin seedlings under catio-nutrient stress in a column growth experiment [J]. Plant and Soil, 378(1-2): 83-98.
TANG C, UNKOVICH M J, BOWDEN J. 1999. Factors affecting soil acidification under legumes. III. Acid production by N2-fixing legumes as influenced by nitrate supply [J]. New Phytologist, 143(3): 513-521.
TANG C, YU Q. 1999. Impact of chemical composition of legume residues and initial soil pH on pH change of a soil after residue incorporation [J]. Plant and Soil, 215(1): 29-38.
TEJNECKÝ V, DRÁBEK O, BORŮVKA L, et al. 2010. Seasonal variation of water extractable aluminum forms in acidified forest organic soils under different vegetation cover [J]. Biogeochemistry, 101(1-3): 151-163.
WANG H, XU R K, WANG N, et al. 2010. Soil acidification of alfisols as influenced by tea cultivation in eastern China [J]. Pedosphere, 20(6): 799-806.
WANG L, BUTTERLY C R, WANG Y, et al. 2014. Effect of crop residue biochar on soil acidity amelioration in strongly acidic tea garden soils [J]. Soil Use and Management, 30(1): 119-128.
XIAO K C, YU L, XU J M, et al. 2014. pH, nitrogen mineralization, and KCl-extractable aluminum as affected by initial soil pH and rate of vetch residue application: results from a laboratory study [J]. Journal of Soils and Sediments, 14 (9): 1513-1525.
YVANES GIULIANI Y A M, WAITE T D, COLLINS R N. 2014. Exchangeable and secondary mineral reactive pools of aluminium in coastal lowland acid sulfate soils [J]. Science of the Total Environment, 485-486: 232-240.
樊宇红, 凌宏文, 朴河春. 2014. 桑树(Morus alba)与丛枝菌根的共生对重金属元素吸收的影响[J]. 生态环境学报, 23(3): 477-484.
郭治兴, 王静, 柴敏, 等. 2011. 近30年来广东省土壤pH值的时空变化[J]. 应用生态学报, 22(2): 425-430.
凌宏文, 朱书法, 朴河春. 2007. 石灰岩和砂岩地区豆科类植物紫花光叶苕(Vicia villosa)中铁和锰的不同分配模式[J]. 地球与环境, 35(2): 97-102.
石娟华, 袁玉欣, 李玉灵, 等. 2008. 铁尾矿坝沙棘、桑树人工林生物量分配及根系分布研究[J]. 河北农业大学学报, 31 (4): 30-35.
唐艳领, 于迪, 胡凤霞, 等. 2014. 黄瓜-玉米轮作对设施连作土壤性状的影响[J]. 北方园艺, (11): 161-164.
佟德利, 徐仁扣, 顾天夏. 2012. 使用尿素和硫酸铵对红壤硝化和酸化作用的影响[J]. 生态与农村环境学报, 28(4): 404-409.
王志刚, 赵永存, 廖启林, 等. 2008. 近20年来江苏省土壤pH值时空变化及其驱动力[J]. 生态学报, 28(2): 720-727.
肖新, 朱伟, 杜超, 等. 2015. 轮作与施肥对滁菊连作土壤微生物特性的影响[J]. 应用生态学报, 26(6): 1-7.
徐厚来. 2014. 参地土壤pH值与连作障碍问题初谈[J]. 中国中医药现代远程教育, 12(3): 113.
杨平平, 徐仁扣, 黎星辉. 2012. 淋溶条件下马尾松针对土壤的酸化作用[J]. 生态环境学报, 21(11): 1817-1821.
杨世琪, 张爱平, 杨正礼, 等. 2010. 黄土高原果园土壤pH变化分析[J].中国生态农业学报, 18(6): 1385-1387.
杨艳芳, 李德成, 赵玉国, 等. 2009. 广西宜州市甘蔗低产原因的调查[J].土壤, 41(2): 315-319.
袁建玉, 周增辉, 张娜, 等. 2014. 设施蔬菜水旱轮作前后耕作层土壤盐分和pH值的变化[J]. 江苏农业科学, 42(3): 301-303.
张光灿, 杨吉华, 赵新明, 等. 1997. 桑树根系分布及水土保持特性的研究[J]. 蚕业科学, 23(1): 59-60.
张建华, 朱波, 廖先谟, 等. 2013. 广西河池桑树黄化的矫正试验[J]. 中国蚕业, 34(2): 12-15.
张作新, 张树明. 2015. 连作对蔬菜保护地土壤理化性状的影响[J]. 农业开发与装备, (3): 70-106.
赵海涛, 李良俊, 段朝珍, 等. 2014. 水生蔬菜轮作对大棚草莓连作土壤性质的影响[J]. 江苏农业学报, 30(2): 289-295.
赵晶, 冯文强, 秦鱼生, 等. 2010. 不同氮磷钾肥对土壤pH和镉有效性的影响[J]. 土壤学报, 47(5): 954-961.
周丽萍, 刘增文, 郭冠春, 等. 2010. 黄土残塬沟壑区林粮间作树种枯落叶对连作麦田土壤极化的防治效应[J]. 西北农业学报, 19(2): 111-117.
The Changes of Soil pH in Mulberry Plantation and Maize Rotation
LING Hongwen, FAN Yuhong, PIAO Hechun
The State Key Laboratory of Environmental Geochemistry, Institute of Geochemistry, Chinese Academy of Sciences, Guiyang 550002, China
With increasing age of mulberry plantation, soil degradation process enhanced, and leaf biomass decreased, which should be associated with decrease of soil pH, loss of nutrient elements and accumulation of harmful element. The soil samples were collected from Yizhou in Guangxi Province and Libo in Guizhou Province, China. The concentrations of exchangeable base cations (Ca, Mg, K and Na), and exchangeable Al, and soil pH and other soil parameters were analyzed in order to study the differences of soil pH between mulberry (Morus alba) and maize (Zea mays) soils, and to discuss the factors influencing the changes of soil pH. Results show that soil pH in mulberry plantations has decreased 0.38 unit relative to maize soils during 5 years. Consequently, the concentrations of base cations (Ca, Mg and K) in mulberry soils were lower than those in maize soils, though not statistically significant. But the concentrations of exchangeable Al in mulberry soils (2.35±2.68) mmol·kg-1were significantly higher than that in maize soils (1.44±2.13) mmol·kg-1(P=0.042). Therefore, there was a significant relationship between soil exchangeable Ca and Al concentrations. The concentrations of exchangeable NH4+-N in mulberry soils were higher than those in maize soils, though not statistically significant. Here, we concluded that soil pH decreased with continuous cropping in mulberry plantation. Correspondingly, soil available Ca, Mg and K concentrations decreased, and soil available Al concentrations increased. Factors causing decrease of soil pH in mulberry plantations relative to maize rotation soils are complex. Among them the cycling of carbon and nitrogen in plant-soil system is the main factor causing the changes of soil pH. Various managements for cropping system also determine the direction and magnitude of pH change. It is suggested that the rotation of cropping plants and return of plant residue into soils should be promoted for increasing soil quality.
soil pH; exchangeable base cation; exchangeable Al; mulberry plantation; maize; rotation
10.16258/j.cnki.1674-5906.2015.05.009
S153.4
A
1674-5906(2015)05-0778-07
凌宏文,樊宇红,朴河春. 桑园地和玉米轮作地土壤pH变化的比较研究[J]. 生态环境学报, 2015, 24(5): 778-784. LING Hongwen, FAN Yuhong, PIAO Hechun. The Changes of Soil pH in Mulberry Plantation and Maize Rotation [J]. Ecology and Environmental Sciences, 2015, 24(5): 778-784.
国家自然科学基金项目(4121004)
凌宏文(1968年),男,高级工程师,从事环境地球化学分析工作。E-mail:linghongwen@vip.skleg.cn *通信作者:朴河春,男,研究员,从事环境地质地球化学工作。E-mail:piaohechun@vip.skleg.cn
2014-12-25
