油用芍药属牡丹组繁育系统研究
2015-12-02吴明海何亚平费世民陈秀明王乐辉蒋俊明
吴明海,何亚平,费世民,陈秀明,王乐辉,蒋俊明
(1.四川省长江造林局凉山分局,四川冕宁 615608;2.四川省林业科学研究院,四川成都 610081)
多功能是众多传统林木种质资源的林业现代化内在要求,木本粮油开发潜能成为我国芍药牡丹类林木的重要产业化培育方向。在中国,牡丹被称之为“百花之王”,在1000年前的瑞典,牡丹被称之为“药中皇后”(Gambrill,1988)。芍药属(Paeonia L.)由3个组组成,即牡丹组(sect.Moutan)、芍药组(sect.Paeonia)和北美芍药组(sect.Oneapia)(Sang et al.,1997;赵宣,2007)。牡丹组有8种两亚种,广泛分布于西北地区、中原地区、江南地区、西南地区(赵宣,2007),均为灌木,野生于灌木疏林的林下、林缘(苏建荣等,2010;马莘等,2011;唐红等,2012),牡丹富含亚麻酸,是健康的食用油(张振中等,2014),其粮油潜力近期受到关注。芍药属包含肉质花盘牡丹组、革质花盘牡丹组、芍药组3个部分,均可作为木本食用油的原料生产材料。潜力评价(capacity evaluation)是自然资源人工开发利用的中间环节。传统名花自身的粮油潜力开发,反映了人类智慧驾驭生殖规律、改造生殖规律,培育优质定向基因造福人类的能力。
木本粮油低产低效与繁育系统本身的生物生态学存在直接关系,如花综合特征、花各性器官的寿命、花开放式样、自交亲和程度和交配系统,它们结合传粉者和传粉行为是影响生殖后代遗传组成和适合度的主要因素,其中交配系统(育性)是核心(刘建全,2003)。结实量主要受到传粉结实等生殖物候过程影响,大多数珍稀濒危植物或多或少存在生殖障碍,诸如雌雄蕊发育不同步、花粉败育、花粉管不能正常到达胚囊及胚胎败育等(周丽华等,2007),表现为雌花子房比例、花粉活力、传粉者限制、自交亲和度、胚胎败育率、生殖配置等。被子植物自交后结籽率的降低通常由早期近交衰退(inbreeding depression)与晚期自交不亲和(self-incompatibility)导致(郝伟祺等,2011),均涉及到上述因素。异交和自交是植物繁育系统研究的中心问题(刘建全,2003),亲属选择(kin-selection)或配子体选择是居群遗传结构形成与繁殖体属性的机制。牡丹结籽率可能是合子前亲属选择、合子后胚胎选择综合作用的产物,胚珠成功受精率和受精卵成功发育率成为决定结籽率的重要指标。
结籽率形成的复杂性,也是生殖系统复杂性的表现,是植物内部的遗传机制和外部环境相互作用的产物,树木濒危过程往往与牡丹低产结实量具有相似的原因,芍药科牡丹组中也有部分种类本身为濒危物种,如矮牡丹、滇牡丹等,生殖系统的效能低下,如结实结籽率低,是树木种子产量低和珍稀濒危的因素之一(陈远征,马样庆,2007);植物濒危的内部因素包括遗传力、生殖力、生活力、适应力的衰竭等,外部因素是生境丧失、片断化、气候变迁等(周丽华等,2006)。在相同的外界生境条件下,普通植物尚能正常生长发育,而濒危植物则不能,关键是其内在属性所致(张文辉等,2002)。野生滇牡丹种群数量下降与其自身繁殖特性有关(李奎,2012);有性生殖潜能低下是矮牡丹濒危的重要因素(赵一鹏等,2009)。
和谐思想认为,事物再生形成经历多元环节,每一环节都可能成为事物形成数量和质量的制约因素,也存在源头决定性的制约因素与中途协同性的制约因素之别。从营养生长到生殖孕育,再到开花结实,果实成熟,脱落而花芽再生,经历了循环的生活史过程,也伴随着个体和群体构型的演变,其促成结实的生活史因素多,影响结实数量和质量的因素复杂。芍药牡丹植物繁育系统集成研究利于木本粮油的定向培育与高产稳产,运用和谐思想,寻找木本粮油繁育系统优化促成结实的适宜的度,及其生产实践定向培育技术的时间次序。定向培育的众多维度,均需要服从于种实生产,其种质、立地、结构、传粉管理等服从于坐果率和结籽率的提高和稳定,芍药牡丹繁育系统信息和特征重要性就显现出来了,能帮助揭示低产原因和高产促成技术途径,以及木本粮油潜力定性定量评价。
国外对芍药牡丹研究开始于20世纪50年代(Waiters,1952),我国开始于20世纪末期,是分类学家起的头(罗毅波等,1998)。牡丹芍药繁育系统研究资料相对丰富,积累结论较多,木本花卉的杂交育种相当丰富,但对木本粮油结实促成的研究成果应用综述还没有。基于潜力评价需要,本文为综述材料,文献从期刊网获得。数据资料从杨山牡丹新都栽培基地中观察测量获得,其中涉及的杨山牡丹育性测定、访花频率、接近频率观测方法见《Pollination ecology:a practical approach》(Amots Dafni,1992)。杨山牡丹枝条物候属性和生殖特征计量方法参照费世民等(2013)。
1 油用牡丹的花部综合征
牡丹芍药类落叶灌木,新梢停长时,花芽孕育随即启动,叶序基部芽蕴蓄枝芽叶芽花芽原基,铜陵牡丹将这一时间定为6月~9月,新梢木质化时期,增粗增壮时期(李兆玉,2013)。有花植物在花序的结构、类型、着生位置、花数以及花部的各种表型特征等方面所展示出的复杂的多样性,常常代表了在其漫长的进化历程中对多种多样的传粉模式的适应(钦俊德等,2001),也直接影响着结籽率和结实率,成为遗传表达和环境修饰后决定结籽率结实率的有机体结构性因素。牡丹为单花顶生,因花枝粗细不同而开花次序不同,呈现大花枝早花,小花枝晚花,过小枝不花。阳山牡丹测定表明,展枝抽梢期枝条生殖属性与大小呈现对数函数关系[Y长=23.412Ln(x)+6.5914(R2=0.9986);Y径=0.1611Ln(x)+0.3605(R2=0.9959)],暗示,枝条大小决定牡丹现蕾与否和开花物候早晚。但在盛花期时,人工浇水使得这种格局打破,而枝条由于异速生长而发生营养枝加粗蓄积、花蕾枝稳定、花枝加高、未受精枝枯萎而壮枝成功坐果的格局。微观尺度上开花坐果微观格局与枝条大小存在依赖性,但在坐果时期,则传粉受精成功与否则影响了生殖行为的持续性,杨山牡丹牡丹花开放式样受制于母枝大小,壮枝培育与合理生殖营养枝配置能优化个体和群体花综合征。母枝大小对个体花综合征呈现决定性作用(费世民,2013),包括同期枝条功能分化格局,对传粉模式和坐果产生结构限制,是牡丹坐果率建成的基础条件。开花期直接影响结实率,陆婷(2010)对粗柄独尾草的研究也证明了这一点,如初花期的结实率最高,其次是盛花期,末花期结实率最低(图1)。

图1 杨山牡丹盛花期枝条大小与功能格局
滇牡丹随着花瓣展开,花药由内而外逐步散粉,属雄蕊离心发育方式,持续时间可达5 d~6 d,有时直到花瓣干枯或凋落后花粉仍有残留或未散尽(李奎等,2013)。四川牡丹花药开裂的方式是纵裂(骆劲涛,2013)。矮牡丹雄蕊离心发育,但最先开裂的仅仅是内轮花药中向阳的几枚花药,然后由内轮向外轮依次开裂散粉;与此同时,先开裂花药两侧的花药也依次开裂;整体来看,同一轮中向阳的花药最先开裂,不同轮之间花药开裂则是遵循离心方式依次开裂;最内轮花药开裂时,一般柱头还不能接受花粉;次轮或更外轮花药开裂时,柱头才能接受花粉;次轮或外轮花药能伸至柱头位置,有时甚至花丝倾斜,花药倒在柱头之间,花药开裂面直接与柱头面接触。矮牡丹可能是雄蕊先熟(罗毅波等,1998)。Schlising(1976)认为 P.califomical雌蕊先熟,在花瓣和雄蕊仍未张开时,柱头已张开,表面湿润、具粘性;一天或更长时间后,柱头表面突起物变硬、干燥,此时最内轮雄蕊开始散粉。野生芍药及栽培单瓣花芍药雄蕊从第一枚花药开裂至全部花药开裂大约需4 h~6 h,到雄蕊全部干萎3d左右,栽培重瓣花芍药存在着个体差异(红雨等,2006)(表1)。

表1 不同栽培设施条件下杨山牡丹成都引种栽培花持续期
单花持续时期也是影响坐果率、结实率和结籽率的直接因素,在传粉者限制的环境风险发生时,则花综合征呈现持续期延长以提高柱头接收花粉的时间历期,也提高了花粉输出的时间历期,如高山低温地区(Binham et al.,1998)。在微观天内阴晴云雨异质性天气情形下,传粉者行为与之紧密相关(牡丹)。四川牡丹单花花期6 d~13 d,居群的花期4月9日~6月4日(骆劲涛,2013)。同属植物芍药,花的单花期在品种、居距群间有一定差异,野生芍药自花瓣张开到花瓣和雄蕊全部脱落约6 d~7 d,栽培单瓣花芍药需要7 d~8 d、重瓣花芍药需要10 d~13 d,不同居群、不同品种的芍药花期相同,均为23 d~27 d(红雨等,2006)。滇牡丹的单花花期一般为6 d~9 d,明显大于一般栽培品种,这与其温度、光照等自然物候密切相关。以采自云南狮子山牡丹园的成熟花粉为材料,研究了贮存温度和湿度对花粉活力的影响,发现低温有利于花粉活力的保持(王兵益等,2001)。牡丹野生生境为中山高山亚高山地带,为灌木乔木林下林缘生境,本身具有相对较低的传粉者稳定性,花持续期与其他木本粮油植物相比,相对较长,如麻疯树(2 d~4 d),但短于高原草地植物麻花艽(9 d)。杨山牡丹成都栽培时,海拔较低,为300 m左右,热量显著大于海拔在2 000 m~3 600 m的牡丹芍药野生栖息地的气候条件,散粉期为1 d左右,在隔离传粉媒介条件下,才能勉强维持11 d左右,单花整体开放历期也在9 d左右,隔离传粉媒介的温室中15 d~17 d左右(如表2所示)。故,花持续期显著低于野生种类和普遍的高山种类,呈现持续期短的现象;这点可能是人工栽培条件下,面临繁育系统变异的问题,其生殖行为和基因库可能会发生截然不同的结局。
杨山牡丹花部综合征中雄性健壮程度与花冠硕大是同步进化的(如图2所示)。伊犁郁金香花的雌雄蕊长度与花瓣长大小间存在正相关性,而雌雄蕊空间接近程度与花瓣大小间呈负相关性(艾沙江·阿不都沙拉木,2013)。滇牡丹同一植株上1个~两个花朵簇生或互生1年生枝叶腋,少数3个花朵互生,3 a生以上植株在合适环境条件下均可正常开花,萌孽植株次年便可开花结实(李奎等,2013)。四川牡丹花托为碗形,中部略突;苞片为绿色,披针形、叶状或者枝条状,数目3枚~5枚;萼片绿色,离生萼片,3枚~5枚;卵圆形或匙形,先端急尖;花冠辐射状,花被卷叠式为覆瓦状。花朵直径67 mm~188 mm,花瓣长41 mm~87 mm,宽34 mm~57 mm;花瓣数量8枚~12枚,颜色为白色、淡紫色或深红色;花丝长6.9 mm~9.8 mm,花药长4.4 mm~7.7 mm(骆劲涛,2013)。大花冠、大花瓣、多雄蕊是芍药属牡丹组革质花盘亚组植物的普遍特征(如表1所示),肉质花盘亚组的滇牡丹则呈现小花特点,但其花粉结块,低胚珠大种子,偏离了传统牡丹的演化方向。在自然授粉条件下,伊犁郁金香的结籽率与花大小间存在正相关性,在隔离昆虫而自主自花授粉时,则结籽率与花大小间存在负相关性(艾沙江·阿不都沙拉木,2013);独尾草繁殖对策研究也表明,结实率与花展示大小呈正相关(陆婷,2010),暗示花大小对虫媒传粉和适合度具有决定性促进作用。牡丹大花展示可能是牡丹进化稳定维持的关键因素。

图2 杨山牡丹成都栽培基地花冠大小与雌雄蕊群大小同步发展现象
四川牡丹雌蕊为离生雌蕊,心皮两枚~6枚(骆劲涛,2013)。阳山牡丹成都栽培时,心皮5.58个(浮动在4个~9个之间),雄蕊数232枚(浮动在86枚~382枚之间),平均一个心皮拥有39.6枚花药,胚珠数58.34枚,单子房胚珠数10.5(未发表数据)。高山植物伊犁郁金香,其花雌雄器官获得资源比例随着花的增大更偏向于雌性方向,存在着大小依赖的性分配特点(艾沙江·阿不都沙拉木,2013)。四川牡丹花朵没有蜜腺但具有强烈的气味,且气味随着花朵的开放会发生改变(骆劲涛,2013),传粉者报酬缺乏,只有花粉回报,并不具有晚期被子植物的花蜜结构,对传粉者吸引力实际上是相当不足的。根据取样滇牡丹花药,得到的P/O值为(6 124~9 713):1(李奎等,2013)。Schlising(1976)还对P.califomica的花粉和胚珠数目进行了统计,花粉对胚珠的比列为46 735∶1~76 133∶1,花粉数目高于胚珠数目5个数量级。四川牡丹的P/O指数的试验结果无法用于判定,原因是P/O值超越了Cruden总结的判定范围(骆劲涛,2013)。而超量的花粉可能发挥了诱物和陷阱的作用,对传粉者趋近吸引作用显著。传粉者不稳定环境中,可能大花和长寿命是同步演化的,如高山地区五花草甸现象(青海植物志),而野生牡丹高山林隙林缘,硕大显著的花部结构与花持续期可能发挥了进化增益的作用。这种大花花综合征,高吸引与高回报,也是牡丹属植物稳定进化的重要保证,是油用培育结实增产的基础条件。
杨山牡丹成都栽培时,在个体地径在0.67cm~1.04 cm之间,个体枝干长度在6 cm~19 cm之间,花枝枝径0.43 cm~0.8 cm之间,个体冠幅在26×17 cm~50×55 cm,个体高度在23 cm~43 cm之间,能达到生殖个体大小的阈值,其年龄在3 a生,即第3年就能发生身体构建营养生长向生殖生长的转变。但从比重看,杨山牡丹第3年时,绝大多数依然达不到这种阈值,可能第4年生是杨山牡丹普遍达到生殖阈值的生殖年龄。这种生殖大小和年龄是杨山牡丹种子幼苗建成节律相适应的,如第1年为生根期,地上部分1 cm~2 cm;第2年为枝干高生长期6 cm~16 cm以下长度的高生长;第3年为高径生长,这种高径同步加速生长则利于富余生殖器官的建成(费世民等,未发表资料),即步入开花结实的器官建成生活史。
2 油用牡丹的传粉过程
油用牡丹结实率结籽率问题涉及到传粉者活动效率,众多研究证实,自然授粉结实率低于人工补充授粉,表明滇牡丹自然状态下的昆虫传粉很可能不能完全满足柱头接收花粉的要求(李奎等,2013),传粉者限制在滇牡丹中是存在。由于受到诸多因素的影响,即使是稳定的传粉者集团,往往在这些因素作用下,可能也呈现不稳定传粉的情形。影响牡丹传粉的因素主要有生境内访花昆虫种类和数量、天气条件、同花期植物种类和数量等(李奎等,2013)。天气状况影响花展示大小和昆虫访问频率,从而也对独尾草结实率产生影响(陆婷,2010)。观测表明,矮牡丹在11点~14点这段时间内,蜂类拜访的次数最多,东坡或东北坡居群,14点以后基本没有阳光直射到矮牡丹花上,蜂类停止在花中活动;蜂类在矮牡丹花中活动频率受天气影响较大,阴天或雨天它们很少在花中活动(罗毅波等,1998)。阴雨天对滇牡丹传粉的影响是巨大的,气温下降,昆虫很少在花间活动,花粉随雨水散落,光照不足影响蜜液的分泌量,很多花朵在授粉最佳时期不能成功授粉,造成自然条件下结实率的下降(李奎等,2013)。芍药属牡丹组均无蜜腺,可以认为昆虫主要在矮牡丹花中采集或采食花粉,而不是蜜液(罗毅波等,1998)。同花期植物对其中某一种植物传粉昆虫传粉效率的影响有两个不同的方面:一是同花期植物种类越多、数量越大,吸引到群落来的传粉昆虫的种类就越多、数量也越大,每一种植物被采访的机会增加;二是同花期植物对某种植物上传粉昆虫的竞争,影响到传粉昆虫对这种植物的采访频率,进而影响到传粉效率。芍药伴生蜜源植物野婴粟(Papauer nudicaule)、山丹(Lilium purmilum)、小黄花菜(Hemerocallis minor)、山刺玫(Rusadavurica)等还接收芍药传粉者[大淡脉隧蜂(Lasioglossum sp.)、芫菁、金龟子和天牛类)]的访问,同花期植物的竞争也是导致芍药居群间结实率不同的原因之一(红雨等,2006)。与矮牡丹伴生的许多植物花期和矮牡丹相同,如红柄白鹃梅、搜疏、紫茧、石竹、山萝花等,花能分泌出各种蜜液,吸引一些具吸器的蜂类,如中国四条蜂Tetravalonia chinensis、金跄条蜂 Anthophora fulvitarsa、带黄胡蜂 Vespula shidai和红光熊蜂 Bombus ignitus。这些蜂类经常在矮牡丹花朵附近飞行,但极少在其中活动,红光熊蜂个体较大,进出矮牡丹花时速度快,对花的振动也较大,或许对矮牡丹的传粉有所帮助,但这种帮助只有利于自花授粉(罗毅波等,1998)。
四川牡丹的传粉昆虫的有小青花金龟、跳甲、长尾管蚜蝇、大灰食蚜蝇、阿坝蜜蜂、切叶蜂和隧蜂,其中数量最多的是鞘翅目的小青花金龟和跳甲(骆劲涛,2013)。Paeonia broteroi传粉者集团主要为Andrena spp.(地蜂)、Apis mellifera(意蜂)、Bomhus ter-restris(熊蜂)、Halictidae(遂蜂科)、Formicidae(蚁科)、Other Hymenoptera(膜翅目)、Coleoptera(鞘翅目甲虫类),以及其他类群(Alfonso,2002)。通过昆虫标本的收集和鉴定认为,滇牡丹花间活动最多的昆虫为蜂类,主要为中华蜜蜂(Apis(Sigmatapis)cerana cerana)和无斑宽痣蜂(Macropis(Sinomacropis)immaculata)(李奎等,2013)。矮牡丹传粉昆虫观察和鉴定结果为,天皇地蜂(Andrenam ikado Strandet yasumatsu)、星地蜂(A.stellaria Hrashima)、地蜂(Andrenam sp.)、上海淡隧蜂Lasioglossum simplicior Cockerell、淡脉隧蜂 Lasioglossum sp.,还有数量较多的斑驼弯腿金龟Hybovalgusbioculatus、长毛花金龟Cetoniam agnifica,4种甲虫,3种体型较大,长15 mm~20mm,1种体型较小,长3 mm~5 mm(罗毅波等,1998)。Schlising(1961)在P.califomica花中收集到17种蜂;Schlising(1976)也观察认为,蜂类是P.califomica的主要传粉媒介,仅观察到一种小甲虫,身体长3 mm~5 mm。Proctor&Yeo(1979)也提到Anthophora acerovum在芍药属Paeonia spp.的花中采集花粉,以及Paeonia delavay的花中可散发出一种似山楂香的气味吸引甲虫。Yakovlev&Yoffe(1957)认为 P.anomala,P.moutan,P.suffruticosa,P.wittm annipana等种类的花中有许多甲虫、蚂蚁和其它昆虫活动。
1916年,Diels提出原始被子植物是由甲虫类传粉的,这一学说得到Takhtajan,Gottsberger等许多人的支持,也受到Bemhard&Thien(1987)的异议。观察发现,甲虫、金龟子、天牛等将牡丹是花的啃食者、交配场所、休息居所,在矮牡丹的传粉媒介中,大体型甲虫应是一种不可靠的传粉者(罗毅波等,1998)。Semp1e在1974年也报道过Poper的花粉粒对甲虫附着能力很差;Bemhard&Thien(1987)认为甲虫不一定都是传粉者。芍药科牡丹组为基部原始被子植物,其传粉者集团具有混合性和过渡性,反映了低效传粉者集团(甲虫、金龟子、天牛)向高效传粉者集团(蜂类)转变的混合特征,反映了芍药牡丹组进化的基部性和过渡性,以及与传粉者的协同进化特性。牡丹传粉者集团复杂性也为油用牡丹高效授粉昆虫选择提供了丰富的资源。往往自然条件下,传粉者行为迟钝,兼性访问较多,而转化者少,如甲虫取食花瓣、熊蜂取食另种花蜜,牡丹缺乏花蜜,花粉作为访花报酬,具有原始性,与昆虫传粉者的协同适应性高效性花部综合征尚未得到稳定进化,可能整体上为低效结实者。牡丹组植物其野生生境本身为高山环境,且,为林缘、林下,蜂类稀少,相当不稳定,可能种类之外,更为重要的是传粉者的绝对数量,访问频次对传粉效率具有直接的评价作用。芍药野生居群主要的传粉昆虫为芫菁、金龟子和天牛,高传粉效率的传粉昆虫大淡脉隧蜂数量不到主要传粉昆虫的1/10,明显低于农校人工居群,而其主要为效率高的意大利蜜蜂(Apis meldiera Linnaeua),且,其数量占绝对优势(红雨等,2004)。
在人工栽培种群中,野生牡丹访花昆虫均能观察到,如甲虫、金龟子、熊蜂、中华蜜蜂、蚂蚁、蝴蝶等,但受到成都平原规模化油菜花蜜源伴生植物的影响,其大量的意大利蜜蜂也成为杨山牡丹的访问者,没有发生低频率的竞争作用,而成为高频率的促进作用。记录54.5 h,熊蜂和意蜂相对访花频率分别为0.0115次·min-1、1.8553次·min-1(访问次数/记录时间),后者为前者的160.89倍;其访花特点为盗粉行为,在花蕾刚刚露瓣而花瓣松弛,存在缝隙时,访问油菜的意蜂直接钻入花内,在花药尚未散粉,就将花粉取食一空,散粉期基本上为盗粉期,为1.02 d±0.58 d,而在隔离传粉者时,散粉期为11.13 d±5.27 d,这种盗粉行为一则没有为自花授粉服务,二则没有为异花授粉服务。意蜂访问存在几种类型,第1种为采集花粉,第2种为趋近行为,第3种为花瓣吸食,第四种为花瓣上清洁身体,其接触柱头的方式为下部采粉触碰或四周上下贴附,意蜂速度低,采粉专业,而熊蜂则飞行速度快,迅速触碰花粉而离开,并没有看到熊蜂专业采集牡丹花粉的现象,可能寻觅花蜜是主要访花目的。杨山牡丹这种因为伴生植物的传粉者而富集传粉者现象,不是传统的竞争行为,而在生活史中为促进行为;杨山牡丹因意蜂盗粉行为而产生过度花粉无效输出的现象,即过度采粉,没有在柱头可受性、花粉活力最高的时期,进行最为高效的采粉输出,而是在花药柱头均为张开时进行强度收集花粉,每分钟1.8533次,且,停留时间相对较长,尤其是在9点~12点左右,为高频访花时段,访花频率在2.19次·min-1~2.57次·min-1,其余观测时段则普遍低于0.895次·min-1(图3)。
过度访花,从检测看,自然传粉条件下结籽率为15%,在去除花药时结籽率依然为15%。这表明,野外环境下,基本上均为异花授粉(可能有单株同化受精的存在可能)。人工辅助杂交授粉与隔离传粉者的自然授粉条件下,结籽率分别为19%和19%(温室内无访花、无摇动)。过度访花和无访花相比,过度访花可能一定程度上降低了结籽率,而无访花一定程度上也未能获得很高的结籽率,二者都较低,但表明,适度访花对杨山牡丹结籽率提高是必要的。观察发现,意蜂密集访花,在花粉初级呈现期2 h内盗粉殆尽,而剩余则低频率回访搜索(包括熊蜂),故,不利于多频次接收花粉输入而提高授粉量。适度控制蜂群规模,而延迟访花历期,尽可能次级花粉呈现时访问,能增进花粉柱头活力最优的吻合性,提高传粉效率,必要时增进延迟自交,增强授粉保障性。

图3 杨山牡丹访花频率的时间格局
3 油用牡丹的花粉柱头活性特征
访花昆虫的采访高峰与花粉活力、柱头可授期三者完全重叠,异花受精的成功率就很高(钦俊德,1987)。芍药露色期较长,1周左右;松雷一大蕾一开放的过程需要1 d~2 d,个体之间差异明显,部分花朵从松蕾到开放只需12 h;盛花期4 d~6 d;盛开的花朵每天16:30左右开始出现闭合的趋势,花瓣合拢;花药在开放第1 d时陆续开裂散粉,但是也有少部分个体在花未完全开放即大蕾期就已经散粉(贺丹等,2014)。滇牡丹在花瓣打开后由内轮到外轮逐渐散粉,花瓣打开前1 d至打开后3 d授粉成果率均较高,直到柱头变暗硬化,散粉仍在持续,可认为具有雌蕊先熟的特性,结合套袋试验的结果,可认为滇牡丹也应属于异花授粉植物(李奎等,2013)。对芍药花粉活力和柱头最佳可授期的研究可知,同一朵花的柱头可授期和花粉活力较强期重叠较长(约为3 d),芍药的最佳授粉期在开花后4h左右(红雨等,2003),这与芍药的主要传粉种昆虫的活动节律同步(红雨等,2004),所以保证了其异花授粉的成功率(红雨等,2006)。
四川牡丹单花散粉的花粉活性呈现出先上升后下降的趋势,而摘下插在水中的试验花朵则散出花粉的活性则呈现出不规律性;单个雄蕊散出花粉的活性同样是先升后降,开花后1 d~2 d内雄蕊开始散粉,在第3 d~4 d达到花粉活性最高值(骆劲涛,2013)。滇牡丹花粉在4℃条件下贮藏1 a后仍具有活力,常温条件下存放10 d后仍有较高活力,利用贮藏了1 a的滇牡丹花粉对中原品种杂交时仍可结实(李奎等,2013)。
四川牡丹柱头可授性,柱头一般从开花第2 d~3d开始具有可授性,一直持续到开花第11 d;在6 d~8 d达到最大可授性,柱头粘液分泌一般从第3 d~4 d开始,一直持续到开花第9 d(骆劲涛,2013)。对于滇牡丹的柱头可授期,适时采集花粉直接对开花前2 d至开花后5 d的花朵进行授粉试验,时间分别记录为 -2 d,-1 d,0 d,1 d,2 d,3 d,4 d,5d,不同时期授粉花朵数为15~25,开花前1 d至开花后3 d结实率大于70%,开花后4d授粉结实率迅速下降,在柱头变硬变暗之后失去接受花粉的能力(李奎等,2013)。牡丹典型的柱头可授性如滇牡丹的研究结果,图3引自原文,可见一斑。杨山牡丹盗粉行为发生在0 d前,距离花瓣绽放至少1 d,在花瓣漏出缝隙时,蜜蜂就强行进入而盗粉殆尽,其访花行为与柱头优化的活性期明显不符合,如滇牡丹柱头可受性期为0 d~3 d(结实率为80%~90%),尽管0 d以下也有40%~70%和3 d以外有30%~70%的结实率(如图4所示)。

图4 滇牡丹柱头可授性曲线(引自李奎等,2013)
芍药牡丹花粉育性与花粉环境温度或花药暴露时间有关,高温干燥时花粉容易失活。新鲜花粉培养5 h~6 h萌发率趋于稳定,储藏花粉培养7 h~8 h萌发率趋于稳定;低温(-18℃)或超低温(-75℃)条件均适宜牡丹花粉的储藏,花粉的耐储藏性依品种而异,‘凤丹’花粉耐储藏性最好,储藏一年花粉萌发率为69.23%,‘岛锦’、‘墨池金辉’花粉萌发率为20%~30%(董兆磊,2010)。在对P.californica的研究中,发现5%~90%的花粉不育(Walters,1942,1952,1956);在矮牡丹中不育花粉的比例为15.82%~54.97%(潘开玉等,1999);大花黄牡丹花粉育性高达96.17%(王士泉等,2012),远缘杂交种‘和谐’的花粉活力为4.1%(荆丹丹等,2011),‘红盘托金,和‘凤丹’的活力花粉所占比例高达92.4%和91.2%,花粉平均萌发率最高时可达85.1%和88.0%,‘粉玉奴’活力花粉仅占40%~50%,‘粉玉奴’活力花粉仅占40%~50%,花粉萌发率仅有30%~50%,异常花粉数比例则高达50%~60%(宋春花,2011)。牡丹花粉胚珠比较高,尽管花粉育性很低,但从雌、雄配子数目及传粉的效率等方面来看,矮牡丹和P.califomica的结实情况应好于实际的结实情况。Schlising(1976)、Grant(1975)都认为P.califomica结实情况差的原因是由于其杂合基因型使得配子体生活力降低而造成的。
Schlising(1971)利用放射性碘标记表明甲虫可携带花粉在不同植株花朵之间传播。矮牡丹可能是雄蕊先熟的,但从蜂类的活动规律来看,特别是星地蜂在访问矮牡丹花之前就可能携带有该种的花粉,并有时直接落在雄蕊群内轮,再进入雌蕊群;认为异花授粉在自然状态下是完全可能的,并认为矮牡丹应属异花授粉植物(罗毅波等,1998)。红雨等(2004,2006)对内蒙古高格斯台罕乌拉自然保护区的芍药P.lactiflora传粉研究表明,认为P.lactiflora不存在无融合生殖,有微弱的自交性,无闭花授粉,是以昆虫为媒介、异交为主要传粉方式的物种。四川牡丹具有很强的异交能力(21.49%结籽率)、较弱的自交能力(7.13%结籽率)和极其微弱的无融合生殖能力(0.06%结籽率),属于混合交配系统,得到ISI=0.9和OCI指数结果为4的说明(骆劲涛,2013)。Grant(1975、1964)认为 P.califomica 是自花授粉为主的,并认为正是由于自花授粉才使得该种的杂合性不被自然选择所淘汰;但Schilising(1976)认为该种是异花授粉植物,并且该种雌蕊先熟的特性更加强了其异花授粉的可能性。自然保护区的芍药两个野生居群的传粉强度和传粉效率小于农校居群,两个野生居群结籽率低于农校居群(红雨等,2006),可能存在不同的传粉机制,即传粉强度高低而形成两类适应性交配系统,即低强度的自花维持和高强度的异花维持。杨山牡丹成都栽培时,温室中隔离传粉者与田间开放传粉时,其均结实,但其传粉机制截然不同,即温室为自花授粉,而田间为异花授粉(这得到田间去雄稳定结籽的证实)。‘凤丹’为异花授粉植物,天然杂交及人工异株异花授粉结实率达到50%以上;同株异花及自花授粉结实率显著下降,雌雄蕊异熟,显著降低自交性;无花粉处理有微弱的结实性(董兆磊,2010)。
牡丹花粉呈现区分为初级呈现和次级呈现(陆婷,2010),访花发生的时间对花粉输入输出活力具有决定性作用,总体看,初级呈现时采粉可能会增进异花传粉,次级花粉呈现时会增加自花授粉。芍药属牡丹组种类雌雄成熟时间存在些许左右,但雌性先熟暗示,初级呈现采粉促进了异花输入,而次级呈现促进了自花花粉的触碰。滇牡丹在开花前2d柱头就有可受性,结实率可达到35%(李奎等,2013)。杨山牡丹成都栽培时,属于初级呈现就采粉,而促进了异花传粉,但并不是同一朵花的初级呈现增进了异花传粉,而次级呈现则促进了自花传粉,而是一朵花在传粉媒介多时,则容易呈现盗粉行为,而初级呈现时传粉,而媒介少时则容易呈现次级呈现时传粉。以专业采集花粉为主的蜜蜂和以采集花蜜为主的熊蜂,构成了牡丹访花传粉模式的分异点,而人工养蜂则访花强度大而呈现初级呈现传粉模式,而熊蜂则因为牡丹缺乏花蜜而呈现快速触碰传粉而呈现次级呈现传粉模式,前者为异花传粉促进模式而后者为自花传粉促进模式。观测表明,杨山牡丹花从初开到花粉败落,意蜂访问呈现剧烈递减趋势,而熊蜂访花则呈现稳定状态,但熊蜂无法发生盗蜜行为,而意蜂能发生盗粉行为(如表2所示),表明,杨山牡丹的意蜂访问盗尽了花粉,而熊蜂访问则无法履行了自花促进传粉的作用,而在意蜂和熊蜂并存时,往往以意蜂传粉效果为主,而熊蜂为主体时,如人工养蜂未涉及的野生居群为主体时,则可能次级花粉输出传粉模式能够形成。杨山牡丹从初开花向衰落花,意蜂趋近频率增加相当显著,意味着采粉回报减少而花冠吸引力增加,熊蜂趋近频率也呈现增加状态,暗示汇报减少而趋而不访增多。
花粉活力和柱头可受性受到温度、水分因素的影响,低温时可能均呈现延长趋势,而干燥时呈现衰退趋势。此时,传粉发生与花粉活力、柱头可受性度吻合与否是传粉模式分化的关键。牡丹不同程度的雌性先熟,意蜂(栽培)和熊蜂等(野生)主导的初级花粉、次级呈现传粉特征,其柱头可受性在开花前1 d和开花后3 d具有较高活性,新鲜花粉5 h~6 h或储藏7 h~8 h活性较高,杨山牡丹实际观察表明,初级呈现盗粉呈现异花传粉模式,而次级呈现搜索访花呈现自花传粉模式,前者吻合性好,而后者吻合性差,则导致了自花传粉的补充性地位。牡丹兼性自交发生延迟,为延迟自交,具有生殖保障作用,而专业采粉者不足而花蜜为主熊蜂传粉系统主导的次级花粉呈现传粉是延迟自交发生的主导条件,这种在杨山牡丹栽培条件下并不明显,而温室中控制试验预计能达到次级呈现效果,仅仅停留在闭门闭户早期,而门户开放时,则意蜂涌入,则导致初级花粉呈现的传粉模式,而延迟自交只能发生在1号温室的隔绝传粉者的等待传粉者的群体,即发生了自动自交(也呈现延迟行为),传粉者不来而花药贴近柱头而发生了自花传粉行为,此时,柱头反卷曲可能是自交发生的根本保证和混合交配系统进化的保障性特征。扁蕾(段元文等,2007;杨霁琴,2013)、大翅霸王(李兵,2012)等延迟自交也是依靠柱头-花药的次级呈现趋近触碰性,牡丹革质花盘亚组反卷柱头可能对次级呈现传粉行为的自交性和异交性均具有巩固性作用。分子检测可能是牡丹交配系统定量化评价的必要条件,可能面临发芽率问题的困扰,即代表性的足量的种子发芽率难保。

表2 杨山牡丹访花和趋近频率的初次级呈现特征
4 油用牡丹的育性特征
油用牡丹结籽率结实率低可能与合子前花粉萌发到受精卵形成的配子体选择有关,往往从花粉阻塞到花粉管分枝、通道阻塞等,均形成了精子细胞释放失效的环节。被子植物双受精过程包括花粉在柱头上萌发、花粉管沿花柱生长、精细胞释放、精卵结合等一系列复杂的过程(徐恒平等,1995),花粉与柱头、花粉管与花柱及雌雄配子之间3个层次中任何一个层面的不亲和都会降低受精率(胡适宜,2005),进而影响结实结籽率。正常的牡丹、芍药双受精过程中,花粉管在授粉后2 h~3 h内便可进人胚珠(李嘉压,1999)。‘紫凤羽’、‘红盘托金,的杂交组合在杂交套袋状态下,授粉后72 h可观察到少数花粉管进入胚珠;开放状态下,进入胚珠花粉管数量增加(宋春花,2011)。
芍药属包括牡丹在内的交配系统研究,发现,自然结实率在58.5%~89%之间,结籽率在5.85%~28.6%之间,相对较低;交配系统研究也表明,芍药属包括牡丹均为自交授粉和杂交授粉均能结实,即自交亲和而异花传粉的繁育系统特征(如表3所示)。结籽率结实率低可能除了上述的传粉过程和花粉柱头活力因素外,可能还受到花粉管伸长、合子形成的过程,生殖过程也包含了不同亲缘关系配子体结合力或育性的问题(属于合子前因素),甚至包含了胚胎发育因素(属于合子后因素)。受精前障碍主要表现为:①花粉管不能在柱头上砧附萌发;②花粉管虽然能在柱头上萌发但是不能进入花柱;③花粉管虽然能进入花柱,但因为花粉管的生长异常而不能进入胚珠(李辛雷等,2008)。花粉管异常又包括:花粉管内部和尖端沉积胼胝质塞;花粉管尖端膨大成节状或球状;花粉管分枝等现象(宋春花,2011)。
人工授粉后‘凤丹白’花粉能够在芍药柱头上正常萌发,但是达到最大萌发量的时间比离体培养所用的时间长,部分花粉管能够伸入柱头,最远可伸人到花柱1/2处,未见花粉管穿过花柱进入子房,完成受精。花粉萌发和花粉管伸长受到了一些阻碍,具体表现为授粉8 h后观察到有花粉豁附到柱头,少数花粉萌发,花粉管伸长;授粉16 h后花粉萌发个数明显增加,花粉管继续伸长,但未伸人柱头,柱头乳突细胞产生胼胝质反应;授粉24h后花粉萌发量达到最高。但是花粉管存在不同形式的畸形,尖端异常膨大,有的花粉管在柱头表面出现缠绕、扭曲、肿胀、结合、分枝等现象,并有部分花粉管伸长迟缓;授粉48 h后,观察到有少量花粉进人柱头,花粉管成束状向花柱延伸,柱头上积累的胼胝质成散点状分布;授粉72 h~96 h,萌发的花粉数逐渐减少,胼胝质积累持续增加(贺丹等,2014)。
母本‘粉玉奴’花朵柱头在花开放第2 d开始分泌黏液,随着时间增加,黏液分泌量逐渐增多,但是第2 d~3 d个体差异较大,第5 d黏液分泌量达到最大,随后逐渐减少,第7 d柱头部分个体有黏液分泌,第8d的柱头没有观察到黏液(贺丹等,2014)。柱头湿润状态,野生芍药可从开花第1 d保持到第5天、栽培单瓣花芍药可从开花第1 d~2 d保持到第6 d,重瓣花芍药有个体差异(红雨等,2006)。花粉落到柱头上随即萌发,双受精过程发生在授粉后4 h~48 h,主要集中在授粉后10h前后。‘凤丹’的受精方式为珠孔受精。精卵融合与精细胞和极核的融合几乎同时完成,以雌、雄性核仁完全融合作为双受精结束的标志,属于有丝分裂前配子融合类型(董兆磊,2010)。对组间杂种(以伊藤杂种为材料)的育性进行观察研究发现,其花期柱头分泌粘液速度快,集中在开花后2 h~3 h,胚珠在开花后第4 h左右开始萎缩,10 h左右完全败育;珠被退化早,胚囊发育迟缓,开花后10 h左右才能到达珠孔端。珠心和珠被发育的高度不协调,可能是导致胚珠败育、杂交不结实的重要原因(王越岚,2009)。
远缘杂交的成功依赖于植物传粉受精过程中每一个环节,从花粉粒的附着、萌发,花粉管的生长,双受精的完成,到合子、胚乳的发育等,其中任何一个环节发生障碍,都会直接导致杂交的失败。因此,授粉前亲本花粉育性、雌蕊发育状况,授粉后花粉在柱头萌发及花粉管生长情况,对杂交结实率有很大的影响(贺丹等,2014)。牡丹杂交父母本遗传距离越大,其结实率影响的主导性不同,种间结种率高低受母本育性的影响较大,亲和性的影响较小;组内(亚组间)远缘杂交的平均结种率受母本育性和亲本亲和性共同影响其结种率;组间远缘杂交的结种率高低的主要受亲本亲和性大小的影响(王越岚,2009)。不同杂交类型间,杂交亲和性各不相同,亚组内近缘种间杂交亲和性高,平均结较高为18.70粒·朵-1,并在母本个体差异较小时,父本花型与杂交结种率有较大的相关性;亚组间远缘杂交的平均结种率明显比组内近缘种间杂交的低,为1.83粒·朵-2,说明该类型杂交在一定程度上不亲和,选择亲和性较高的亲本杂交组合是提高杂交结实率的有效途径;组间远缘杂交的亲和性极低,平均结种率极仅为0.52粒·朵-2,寻找亲和性较高的组间远缘杂交组合以及育性较强亲本资源是突破这类型杂交的重点(肖佳佳,2010)。‘凤丹’与日本牡丹杂交结实率较高,江南品种次之,与美国黄牡丹杂交种杂交结实率极低,败育现象严重(董兆磊,2010)。
牡丹与芍药远缘杂交表现出严重的不亲和性,授粉579朵,结果数30获得种子49粒,结果率5.2%。在开花期标记300个芍药花朵,果熟期统计标记花朵的结果数为228,芍药的自然结实率为76%(贺丹等,2014)。综合几种杂交细胞学研究结论,牡丹杂交不亲和的细胞学机理主要有:父本的败育;柱头对花粉的主动识别作用导致受精过程障碍;胚乳发育异常导致了胚胎的败育(宋春花,2011),如表3所示。

表3 几种牡丹杂交亲和性的细胞学解释(宋春花,2011)
国外已培育出了不少牡丹与芍药组间杂种,但均高度不育,雄蕊常呈花丝状,花药无花粉或少量花粉。远缘杂交种‘和谐’的花粉活力为4.1%,说明‘和谐’雄性有一定的育性(荆丹丹等,2011),但‘和谐’作母本与牡丹、芍药杂交不亲和。牡丹芍药没有无融合生殖现象,居群内交配成油用牡丹基地的基本传粉行为。李守丽等(2006)对大百合与百合属间杂交授粉后花粉管的行为进行了观察,百合的花粉萌发与花粉管的伸长速度相对于自交滞后,个别花粉管出现末端分叉、膨大或变细或中间部分折叠,花粉管内脐眠质不规则的大量沉积,这些识别反应导致结实率低。因此,受精前障碍是远缘杂交结实率低的重要原因,授粉后花粉管正常生长并穿过柱头是牡丹芍药远缘杂交较高结实率的重要保障。种类近缘杂交利于提高结实率结籽率,而最为亲缘的自花传粉可能也具有较高的结实潜能,杨山牡丹栽培观察表明,在存在花粉收集传粉者情形下,花粉很难正常呈现,盗粉行为显著而虽然牡丹P/O相当大,花粉量相当大,但依然呈现花粉限制或传粉者限制,尽管访花频率相当高,花粉输出相当显著。在野生和人工栽培居群中,芍药牡丹群落存在两大系统,一个是捕食者集团,昆虫同资源种团(guide);一个是受精者集团,牡丹芍药及其同花期伴生的植物蜜源粉源植物,二者是相互独立而相互作用的和谐系统,构成了芍药牡丹的传粉系统(包括花和传粉者)。牡丹与伴生植物传粉者具有竞争作用(罗毅波等,1998;红雨等,2006),也有促进作用,如油菜采粉者意蜂与杨山牡丹异花传粉保障。合理控制人工养蜂的规模,改过度采粉为轻度采粉,则可能芍药牡丹结实结籽会发挥近缘交配的作用,提高最可靠育性的量和质,进而提高牡丹芍药的结实结籽率(表4)。
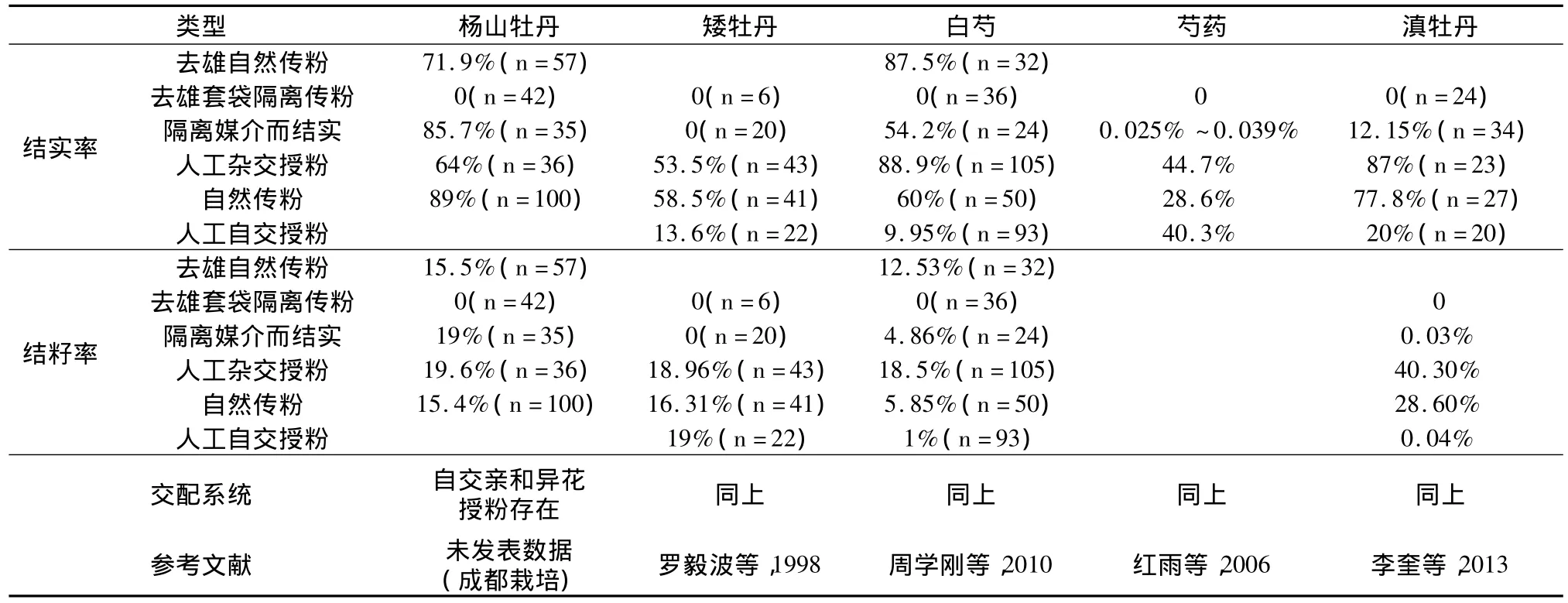
表4 芍药属(包含牡丹组)植物的结实率和结籽率育性检测控制试验结果
5 油用牡丹受精卵的发育命运
受精卵不总是全部发育为完整的我胚胎,解剖芍药牡丹果实,总是存在不同大小的败育胚胎,这些胚胎是在受精卵发育为成熟种子的历程中逐渐败育的,构成了牡丹芍药结籽率的合子后选择机制。杨山牡丹果荚解剖时发现,在受精卵发育为种子途中,存在败育遗迹,也存在成熟种子;还存在一种情况,即无完好种子,而仅有败育胚胎,但果荚也能正常发育,暗示,受精卵发育是启动果实发育的关键,在没有受精时,果荚能假膨大,但无法持续发育。不同类型处理的杨山牡丹败育率在5.04%~8.32%之间(如表5所示),也存在诸多因素促使胚胎不能正常发育。
胚胎发育的启动标志如何从花部综合征上进行度量?一些研究发现,在双受精成功后则胚胎发育启动,花部营养向子房集中,花冠缺乏维持有机物质和能量供应,则花瓣逐渐脱落,或者全然闭合,不再重复性或适应性的开闭运动。杨山牡丹花果发育观察中,发现在传粉发生后即发生花瓣脱落,而温室中传粉发生较慢,则花粉散落较晚,二者从露蕊开始,散粉至开始落瓣,温室经历5.326 d,半开放温室落瓣经历6.036 d,田间经历4.579 d,表明进入受精后合子发育为胚胎,进而建成种子的过程。故,种子发育启动则从芍药牡丹落瓣为标志较为适宜。经过计算,杨山牡丹种子发育完整时间为120 d~130 d,在成都新都基地,基本从3月中旬、下旬开始到当年7月中旬、下旬结束。
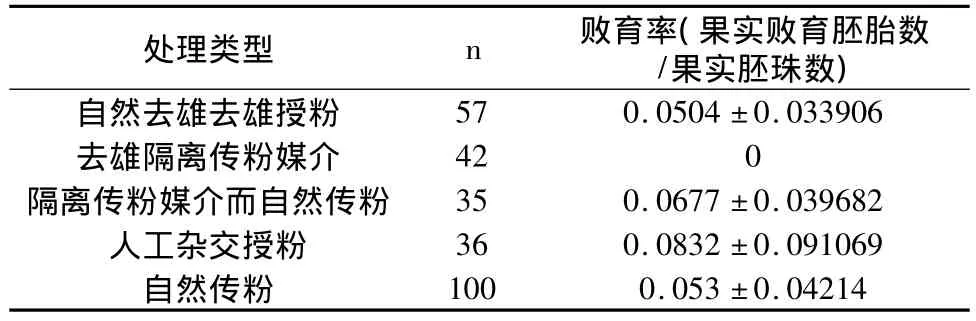
表5 不同试验处理杨山牡丹的败育率
关于 P.californica,Walter(1962)认为,每个胚珠中有30个~40个大孢子母细胞可进行减数分裂,受精时,每个胚珠中有1个~4个大配子体;Cane et al(1961)以及Walter(1962)还发现每个胚珠中可具几个前胚或胚,因而每个发育的胚珠中就具补偿合子或胚生活力较低的能力;Walter(1962)甚至还讨论了这些配子体或幼胚之间发生竞争的可能性。从 Caneetal(1961)及Walter(1962)等人的工作看,在P.californica中每个胚珠都应该是可育的,尽管他们并没有进行这方面的统计。
在牡丹芍药受精的卵均未发育为成熟胚胎时,‘紫凤羽’自然状态和‘紫凤羽’ב红盘托金’杂交状态下,其胚胎均发育正常,但因实验材料限制未能找到胚胎发育从棒形胚到子叶型胚的全过程,胚乳发育形式为核型胚乳,经历游离核阶段和细胞化阶段,并在合点端发现胚乳吸器(宋春花,2011)。对组间杂种(以伊藤杂种为材料)的育性进行观察研究发现,其花期柱头分泌粘液速度快,集中在开花后2 d~3 d,胚珠在开花后第4 d左右开始萎缩,10 d左右完全败育;珠被退化早,胚囊发育迟缓,开花后10 d左右才能到达珠孔端。珠心和珠被发育的高度不协调,可能是导致胚珠败育、杂交不结实的重要原因(王越岚,2009)。‘凤丹’为核型胚乳,初生胚乳核不经过休眠直接进入分裂阶段,形成游离核胚乳,持续时间约为23 d,授粉后45 d胚乳完成细胞化,进入生长成熟阶段,授粉后85 d胚乳发育成熟。合子休眠期为8 h~12 h,胚的发育略晚于胚乳的发育,但其细胞化时期与胚乳基本同步。胚的发育经历原胚阶段、器官分化阶段和生长成熟阶段3个连续的发育时期。其中原胚阶段的变化过程最为复杂,持续时间约为55 d,合子的分裂并不伴随着细胞壁的产生,形成游离核原胚,游离核增值到一定数量即开始细胞化,细胞化原胚发育到一定阶段即沿着球形胚-心形胚-鱼雷胚-子叶胚的发育过程完成胚发育,授粉后125 d胚发育成熟(董兆磊,2010)。‘紫凤羽’、‘凤丹’、‘粉玉奴,自然状态下、以‘粉玉奴,为母本的两个杂交组合杂交后,其胚胎均存在胚胎异解现象,败育胚胎中,胚乳絮状异解导致了胚败育(宋春花,2011)。
杨山牡丹结籽率和败育率呈现线性关系,在样本数为137时,R2为0.094时能呈现显著性(如图5所示);杨山牡丹结籽数和败育数也呈现正相关,相关系数相对很高,R2为0.4603。受精卵形成幼胚能否发育为成熟种子,可能与有机资源供应有关,在有机资源充足时,发育概率较大,而在有机资源不足时,败育概率较大。麻疯树木本粮油研究时,将母枝大小与无机立地两者综合起来,发现败育率与结构、资源制约有关,即结构限制和立地资源限制时,胚胎败育概率增加(费世民等,2011)。有机和无机营养合成运输功能低下时,幼胚发育时种子大小和数量上同步调节,一则数量上减小,二则质量降低,可能胚珠本身的异质性对胚胎发育也产生分化,同时,优先占位可能对败育也能起到解释作用。结籽率与败育率同步增长,暗示,胞间呈现竞争作用,完好种子越多则败育胚胎增加,这种胞间竞争也是胚胎发育与结构资源因素相平衡的重要机制。从结籽率提高看,在存在胚胎败育时,就不要考虑传粉不足的问题,这是木本粮油高产稳产技术模式关注焦点的关键指标。油用牡丹考虑传粉,是因为坐果率的问题,牡丹芍药的坐果率徘徊在58.5%~89%之间,存在很大的提高余地,在坐果率达到95%以上时,则油用牡丹的传粉制约问题就无须考虑了。上述数据是在栽培早期条件下获得的,光温水肥容和谐时可能呈现截然不同的坐果率和结籽率,能好于当前所有的试验数据,也许结籽率和败育率的同步增长关系会发生改变,则油用牡丹定向培育关注焦点问题也会改变。从杨山牡丹胚胎败育看,传粉问题和立地、结构问题同样重要,这种传粉问题可能属于类似于病虫害管理一样的,是定向培育的重要维度。具体措施,适度而禁止大规模人工养蜂,可能是一种提高授粉效率的方法,延长访问期,花粉全悉高效利用但能保持3 d~54 d的访问时期就能提高授粉。传粉必须在结构、资源保障基础上考虑,否则多维制约犹如木桶原理,任何一个环节短板都会降低油用牡丹总体的培育实效。
“花多果少”、“胚珠多种子少”,是植物的普遍特征,也是生殖生态学上的重要假说,结实率和结籽率是度量二者的关键指标。牡丹种子一般呈椭圆形或卵状球形,成熟时一黑色或黑褐色,一些结实率高的品种由于蓇葖果内种子多互相挤压而呈多面形(刘心民等,2005),千粒重随着品种而各异,一般观赏牡丹千粒重为195 g~311g(杨辉等,2006)。研究发现,芍药属很多种类结籽结实率与产量普遍较低(罗毅波等,1998;;李奎等,2013;杨辉等,2006;周学刚等,2010;Schlising,1976;红雨等,2006)。安徽原产的杨山牡丹引种到成都栽培时,当年结实率89%,结籽率15.39%,单果2.16粒,单荚9.45粒(n=100)。油用牡丹芍药的高产促成结实和优质稳产,成为定向培育的基本要求。这里建立一种分析评价方法,以利于生产方法措施的制定,如依照等级配置要求来合理配置个体资源,而营养优先配置,其次才是生殖配置,故,首先保证群体营养条件,如立地、光照、结构、遗传等,提高群体生产力,其次,考虑传粉过程对坐果率的保证,尽可能将坐果率提高到95%以上。再次,考虑败育率,在败育率存在时,则表明可能存在结构资源性制约因素,此时,将二者适当调节,提高有机资源对胚胎发育的保障,进而达到提高整体产量并保持连年优质稳定的目的(费世民等,2013),从花—果—种子转化率上建立牡丹芍药的高产稳产监测属性的长尺度多年分多立地的构型和产量数据计量方法,这是木本粮油研究的重要思路。只有这样才能运用本综述集成的开花结实的花部综合征、传粉过程、花粉柱头活性特征、育性特征,以及受精卵的发育命运等,来结实定向培育方法技术体系自身对牡丹高产稳产的内在影响因素。
6 油用牡丹繁育系统综述运用
牡丹传统以来,作为花卉和药用,具有广泛的栽培基础,长期以来形成了5大牡丹群。油用牡丹是21世纪以来的新事物,但牡丹天生低产,单产相当低,150 kg·hm-2~300 kg·hm-2是上限,其油品质相当高,营养保健价值好,成为我国粮油安全新兴产业的重要方向。芍药属植物均具有较高的花粉胚珠比,花硕大,高雄性投入可能与林缘早春开花吸引传粉者而实现生殖保障有关。杨山牡丹引种到四川成都,测量观测发现,其花雌雄蕊大小(雄蕊直径、雌蕊高、花丝长、花药长)与花冠大小呈现正增长关系,传份昆虫吸引力与雌雄适合度潜能具有正相关性。花枝大小对花大小、开花早晚具有作用。隔离传粉者能延长开花历期,增进雌雄适合度实现的时间期限,如油菜花盛开传粉者数量频度很大,则仅仅有1 d左右的开放时间,而花瓣脱落,花的雄性功能终止,雌性功能增加停止,而花粉萌发和受精则启动了。壮枝利于早花早实,从观察看,壮枝—大花也具有较高的雌雄适合度和结实潜在条件。
杨山牡丹具有显著的初级花粉呈现,即在花瓣尚未展开时,花药尚未开放时,为花药呈现,但此时蜜蜂盗粉行为使得花粉散失殆尽,而在花瓣开放时,基本为无粉花药;花粉自然散粉仅仅能发生在温室条件下,温室条件下隔离了传份昆虫,使得花粉能自然呈现,即通过花药爆裂而花粉溢出,为次级花粉呈现。杨山牡丹独特的盗粉行为,使得传粉过程呈现畸形状态,即牡丹的雄性投入,可能生态系统服务作用明显,而适合度作用次之,如保障蜂群食物供应。这种传粉效率的低下,使得芍药牡丹可能存在一种悖论,即花粉异常丰富,而传粉效率相当低下,呈现花多果少种子更少的特点。牡丹独特的散粉行为,可能对生活史具有深远的影响,即降低了结实量而可能存在种子品质或成苗能力相对较强的权衡现象,即时间权衡。这点在牡丹自身的种子繁殖特性得到验证,即牡丹种子具有独特的生活史二分现象,早期萌发成苗种子,即第一个春化作用发生后立即萌动,而存在土内发育现象,即春化作用发生后,不立即出土萌动而是在土内埋藏至秋冬,萌动生根,而来年出土。这种种子萌发的时间分工,可能表明其具有独特的繁殖策略,即低产而萌发率相对较高。

图5 杨山牡丹结籽数(率)和败育数(率)的线性回归分析
杨山牡丹,乃至芍药属的结实结籽的传粉限制,不是花粉相对多少来定位的,而是结实率和结籽率,人工试验表明,自然授粉和人工辅助授粉,在结实率和结籽率上存在些许差异,不构成传粉限制,但人工辅助授粉似乎提高了一些结实率和结籽率。而观察资料,而不是芍药属多种试验数据,更能证明传粉限制,即杨山牡丹雌蕊多5枚,而结实率自身浮动在60%~90%,结籽率浮动在5.85%~28.60%,前者说明存在未授粉的雌蕊,存在传粉限制,后者说明,存在大量未授粉的胚珠,存在传粉限制。人工辅助授粉并未得到满意的结实率和结籽率,这表明,充分授粉提高结实率和结籽率的潜力是有限的,暗示,试验时可能存在其它的原因,如结构制约、资源制约,以及遗传制约,但传粉限制是确定的,从未授粉的雌蕊和很低的结籽率得到直接的证明。初级花粉呈现时访花频度相当高,趋近频度相对较低;次级花粉呈现时,花瓣脱落,则访花频率很低,趋近频率相对很高。和传统的花粉限制调节方法不同,杨山牡丹促成结实,调节花粉限制的方法则为,适度授粉,遏制初级花粉呈现时期的传粉行为,而促进花粉次级呈现时期的传粉行为,即芍药属牡丹组必须适度控制同花期伴生植物种类的访花蜂群规模而不能短时超强度过度采粉。人工辅助授粉能帮助提高授粉效率,但从实践中是无法完成的,需要在控制伴生油菜蜂群数量规模,才能保障牡丹花粉次级呈现,才能得到人工辅助授粉的花粉源,而控制伴生植物的蜂群而遏制盗粉行为,培育次级呈现花粉,为人工辅助授粉提供条件。自然条件下,很难得到花粉,都是盗粉行为惹的祸。
在育性上,依照隔离媒介而结实来判断,则芍药属种类自交亲和性存在,如杨山牡丹、白芍、滇牡丹等;也存在自交不亲和,如矮牡丹、芍药等。在育性上,依照人工自交辅助授粉来看,则矮牡丹、白芍、芍药、滇牡丹等,均是自交亲和的,而成都引种时杨山牡丹这组试验缺乏,而温室隔离媒介自然传粉则清晰证明自交亲和的。这种试验测定数据可能缺乏多年多居群重复性,不能得到可靠的结论,综述中刻意回避这个问题较为合适。从杨山牡丹实际来看,自交亲和性是良好的,异花传粉也是良好的,这组试验为亲自观察结果,结论是可靠的,即杨山牡丹呈现自交亲和而异花授粉存在混合交配现象,其他牡丹也试验测定表明为这种交配的特性,如矮牡丹、白芍、芍药、滇牡丹等(红雨等,2006;李奎等,2013;罗毅波等,1998;周学刚等,2010)。北美植物Collinsia verna(春锦龙花)具有较低的访花频率,发现,该植物呈现自交亲和与异花授粉的混合交配系统特性,在传粉者媒介稀少时则为自动自花授粉(autonomous self-pollination),自交率估算也表明,在野生环境下,传粉者稀少而自交率增加(Susan Kalisz et al.,2004)。芍药野生居群主要的传粉昆虫为芫菁、金龟子和天牛,高传粉效率的传粉昆虫大淡脉隧蜂数量稀少,农校人工居群的高效率的意大利蜜蜂数量占绝对优势(红雨等,2004)。牡丹混合交配系统,在不稳定传粉环境中,可能存在自交和异交授粉的均衡问题;杨山牡丹温室隔离昆虫试验也表明,露天栽培去雄自然授粉,结实率和结籽率基本上与露天自然授粉的结实率和结籽率相当(如结实率71.9%vs 89%、结籽率15.5%vs 15.4%),暗示,蜂群密度大时,访花频率高时,异花授粉相对较为显著,而温室内仅仅为自动自花授粉,呈现交配育性与传粉环境高度依赖性,即杨山牡丹也呈现Collinsia verna的类似自动自交的生殖保障机制;从生物学看,杨山牡丹的反卷柱头是实现自动自交的重要性状。此时,专门探讨育性或交配系统特点,从运用看,油用牡丹高产稳产,则绝对控制蜂群而自动自花授粉,可能是合适的,然后人工辅助授粉,或者借助轻度蜂群授粉而保障长时间授粉效率的提高。
花多果少的生殖生态学现象,表明,不但传粉自身存在结实率的选择败育,结籽率也存在选择败育,使得受精卵形成后其命运也存在多向性,如结籽和败育。杨山牡丹不同结籽处理的结籽败育率浮动在5%~8%之间,自然结籽率15.4%,且,败育率和结籽率呈现正相关,暗示,受精卵越多,结籽率越多,败育率也越多,亲属同胞为竞争关系。受精前障碍,表现为萌发障碍、花粉管伸长障碍、精卵结合障碍,如存在花粉管内部和尖端沉积胼胝质塞;花粉管尖端膨大成节状或球状;花粉管分枝等现象(宋春花,2011)。受精后,受精卵发育受到胚珠自身的结构特点(如母体营养供应疏导能力)影响,还受到种子数量的权衡作用,即同胞竞争,种子数量大时这种同胞竞争作用较为明显,表现为种子数量和败育种子数量、结籽率和败育率之间的正相关关系。简单归因于花粉因素是不合适的,大量研究,尤其是探究芍药牡丹杂交育种、远缘杂交育种时,结实率结籽率低的问题,做了大量的解剖学工作,为分析这一问题提供了素材(董兆磊,2010;宋春花,2011;李辛雷等,2008;贺丹等,2014)。受精后雌性适合度或命运则受到种子数量的显著调节。
依照花粉柱头最优活性期,适当调整伴生植物访花昆虫的规模,将访问强度控制在轻度上,而不过度在处级呈现花粉时期盗粉而短时间集中移走花粉,而使得花粉输入的频次和概率严重不足,进而制约受精卵的比重和胚胎形成的初期数量,即传粉行为发生与最优花粉柱头活性相吻合。花粉活力和柱头活力可能对受精卵数量质量存在影响。芍药牡丹花粉活力变异很大,从40%到90%均有,花粉育性相对较低,受环境因素影响较为明显,如温度。花粉活力问题可能对结实率和结籽率产生解释力。
开花传粉受精以及受精卵发育等生活史过程,均与芍药牡丹营养结构和无机资源环境紧密相关。无机资源立地调节要适宜于芍药牡丹定向培育的需要,利于发挥该种质最大的遗传潜能。结构控制要保障当年壮枝高产和可持续营养的适度均衡,这是坐果率提高到95%以上的关键。资源制约和结构制约的优化控制,能为败育率的降低或结籽率败育率正增长关系的平衡化或负化提供基础条件。油用芍药牡丹高产稳产需要运用和谐思想,使得定向培育各个维度均能得到生活史多环节的促成结实正贡献,能服务于高油高产多抗,使得传粉媒介行为与生物学最优区间吻合,以充分发挥遗传学因素,而达到高效优质可持续生态培育的目标。找寻生产环节,并认识早期环节的决定性与晚期环节的包容性,只有不遗余力的巩固好早期环节,才能为晚期环节或目标的实现创造条件,种子与来年种子产量是关键,而不仅仅涉及到当年高产,还有来年高产,存在显著的时间权衡现象。这些需要在林油一体化基地建设中统筹兼顾。
油用牡丹是木本粮油功能的开发利用,是林木多功能的体现,是现代林业理论指导下的产物。油用牡丹主要是凤丹,或杨山牡丹,即从传统的药用牡丹发展而来。牡丹生殖过程自身具有天生的低产性,本综述仅仅是定性概括,有限的试验数据中得到的结论,需要量化测定,来精确评估芍药牡丹花卉药用和食用多功能利用潜能。牡丹为花王,芍药为花相,是我国传统花卉中杰出者,在我国园林观赏文化中具有突出地位,成为我国审美文化的重要构成。牡丹多元食用功能的研究也为粮油产业功能建设提供了条件。
牡丹组为属下单位,是芍药属的,芍药科目前为单属科,约30多种,分布于北温带,大部产亚洲,部分种类分布在欧洲南部和北美洲西部,其自身在繁育系统上存在地理、遗传和生态联系,人为干扰也存在重要的进化影响,其繁育系统量化测定需要在芍药属广泛的背景下进行,才有价值,找寻这类基部被子植物的生殖特征,探究从裸子植物向被子植物过渡时的生殖生物学和生态学特性,这种整体规律探究利于评价基于整体演化植物界的木本粮油产业化的潜力。芍药属生活型分化明显,有大灌木、小灌木与草本,种类不多,在栽培利用和理论探索上均具有良好的材料特征。芍药牡丹自古为名花,种质材料收集工作完成很早,自然野生和人工栽培条件下繁育系统的变异及其理论解释可能对综合利用有利,纯野生、栽培和集约化培育条件可探究人工培育的进化潜力。
芍药牡丹理想生殖潜力的发掘,这种生殖潜力是结合野生、粗放栽培和长期驯化梯度条件下的繁育系统变异而探讨的人工定向培育条件下生殖产量的阈限问题。这是该类花卉药用植物作为粮油种质资源开发利用中伴随的一个重要问题,即在定向培育科技研究中自然潜力和人工潜力的问题,人工改良能力量化评估问题。自然开花结实的芍药牡丹与人工培育的芍药牡丹,其繁育系统特性和生殖产量比对,就能显示人工改良能力的潜能,这是木本粮油潜力评估的基础数据。
芍药牡丹广泛谱系上的繁育系统量化研究,能从系统学、分类学、分子生物学、传粉生物学、生殖生物学、生殖生态学、森林培育学、经济林、生态学等方面广泛建立多学科的交叉上形成认识有机联系的渠道和平台,促进众多问题的系统认识,增进问题解决的深度和广度。不单单从一个片段研究,还从开花、传粉、花粉生长和受精过程,以及受精卵命运上系统研究评价,在地理、生态、遗传、人为干扰等因素作用下的生活史研究,培养多元系统综合科学思维方法和更有潜力性的挑战性的实用性的研究任务。
牡丹芍药生产群体的生殖潜能评价研究不能脱离群体结构,如纯林、混交林,二者存在不同的制约因素,其繁育系统自身的变异和结实潜力可能存在差异,也存在遗传适应的潜在条件。地理气候、密度与群落结构、生殖年龄、水肥、树体构型等对生殖潜能的表达存在综合作用,也伴随着问题的多根本性、连锁效应等现象,还要从有机与无机生态平衡上探究光温水土容多元立地因素对整体生殖潜力的塑造作用。在生态学整体上,无机和有机,无机综合系统和有机综合系统上探究高产稳产的培育方法体系,其中蕴含着生物生态学量化的人工运用潜力。遗传对生殖潜力贡献到底有多大,乔灌草生殖配置特征决定了草本生殖潜力明显大于乔木灌木,这种生活型尺度上遗传,即种属遗传差异对生殖塑造能轻而易举的证明这个问题,即贡献很大,而在种下、种上遗传贡献的量化评价则反映了基础研究和应用研究,以及示范研究的深度和功底。
芍药牡丹自古为花卉,多元人工驯化品种繁育系统特征和结实潜力评价问题,如花部综合证、传粉过程、花粉柱头活力、花粉管生长和受精卵命运等,选择可度量的性状进行量化评估,以选择优良品种。这种生殖潜力量化评估方法,也能促进粮油潜力变异体和优株的发现,这种量化评估是基于对繁育系统规律的本质研究运用的结果,没有本质规律的研究,就没有量化评价运用。精细过程的认识是潜力评价必不可少的信息。
芍药牡丹野生栖息地的保护修复,与历史以来人工驯化种质资源的集中收集、定向培育改良和量化生产示范研究,以及人工驯化繁育系统再度逸生,丰富自然野生基因库构成。从野生到人工,从人工到野生,也涉及到繁育系统的变异适应进化的问题,而人工改良在这一过程中发挥了撬动作用。珍稀濒危和经济价值种质的迁地保护和科普价值,在带动芍药牡丹繁育系统研究中将发挥重要作用,国家尺度和省区尺度的动植物园公园花园牡丹芍药专类园,以小规模试验示范基地,可能为其种质的安保、产业、环保、防护和景观多功能的有序发挥创造了条件。芍药牡丹繁育系统单一理想环境评价和面上多元地理环境下的协作研究,需要并举。
[1]Alfonso M Sanchez-lafuente.Floral variation in the generalist perennial herb Paeonia broteroi(Paeoniaceae):differences between regions with different pollinators and herbivores[J].American Journal of Botany,2002,89(8):1260~1269.
[2]Gambrill K W.King of flowers,queen of herbs:the peony[J].Pacific Horticuture,1988,49:26~32.
[3]Grant V.The Architecture of the Germplasm[M].New York:John Wiley and Sons,1964.
[4]Grant V.Genetics of Flowering Plants[M].New York:Columbia Univ Press,1975.
[5]Hong Y(红雨),Liu Q(刘强).Foraging and pollination insects of Paeonia lactiflora Pall.(芍药的访花昆虫和传粉昆虫)[J].Entomological Knowledge昆虫知识),2004,41(5):417~423.
[6]Robin A.Bingham,Andrea R.Orthner.Efficient pollination of alpine plants[J].Nature,1998,391:38~239.
[7]Sang T.Daniel J Crawford,Tod F Stuessy.Chloroplast dna phylogeny reticulate evolution,and biogeography of Paeonia(Paeoniaceae)[J].American Journal of Botany,1997,84(9):1120~1136.
[8]Schlising R A.Reproductive proficiency in Paeonia californica(Paeoniaceae)[J].Amer J Bot,1976,63(8):1095~1103.
[9]Susan Kalisz,Donna W.Vogler & Kristen M.Hanley.Context-dependent autonomous self-fertilization yields reproductive assurance and mixed mating[J].Nature,2004,430:884~887..
[10]Waiters J L.Heteromorphic chromosome pairs in Paeonia californica[J].Am J Bot,1952,39:145~151.
[11]Waiters J L.Spontaneous meiotic chromosome breakage in natural populations of Paeonia californica[J].Am J Bot,1956,43:342~354.
[12]Walters J L.Distribution of structural hybrids in Paeonia californica[J].Am J Bot,1942,29:270~275.
[13]艾沙江·阿不都沙拉木.伊犁郁金香的繁殖生物学特性及其生态适应对策[M].乌鲁木齐:新疆农业大学(博士学位论文),2013.
[14]陈远征,马样庆.濒危植物生殖生态学研究进展[J].中国生态农业学报,2007,15(1):186~189.
[15]董兆磊.‘凤丹’(Paeonia ostii‘Feng Dan’)生殖生物学的初步研究[M].北京:北京林业大学(硕士学位论文),2010.
[16]段元文,何亚平,张挺锋,等.高山植物扁蕾的延迟自交机制[J].植物生态学报,2007,31(1):110~117.
[17]费世民,何亚平.麻疯树生殖生态学[M].北京:中国林业出版社,2013.
[18]费世民,何亚平,王乐辉,等.金沙江干热河谷麻疯树结籽率及其影响因素研究[J].四川林业科技,2011,32(1):1~12.
[19]郝伟祺,赵鑫峰.被子植物早期近交衰退与晚期自交不亲和[J].生物多样性,2011,19(1):.
[20]贺丹,王雪玲,高晓峰,等.牡丹芍药远缘杂交亲和性[J].东北林业大学学报,2014,42(7):65~68.
[21]红雨,刘强.芍药的传粉生物学研究[J].广西植物,2006,26(2):120~124.
[22]荆丹丹,刘政安,李新旭,等.牡丹远缘杂种‘和谐’育性的初步研究[J].林业科学,2011,47(10):59~62.
[23]李兵.荒漠植物大翅霸王雌性先熟和延迟自交的功能特性[M].新疆农业大学(硕士学位论文),2012.
[24]李奎.滇牡丹保护生物学与遗传多样性研究[M].北京:中国林业科学研究院(博士学位论文),2013.
[25]李奎,郑宝强,王雁,等.滇牡丹的开花特征及繁育系统[J].东北林业大学学报,2013,41(1):63~67.
[26]李守丽,石雷,张金政.大百合与百合属间授粉后花粉管生长发育的观察[J].园艺学报,2006,33(6);1259~1262.
[27]李兆玉.铜陵牡丹生产与加工[M].合肥:安徽科学技术出版社,2013.
[28]刘心民,程逸远,张雾,等.牡丹种子萌发特性与播种繁殖技术研究进展[J].河南林业科技,2005,25(4):38~40.
[29]陆毛珍.芍药科Paeonsaceae系统位置的研究[M].兰州:西北师范大学,2006.
[30]陆婷.独尾草属两个同域种的繁殖对策[M].乌鲁木齐:新疆农业大学(博士学位论文),2010.
[31]罗毅波,裴颜龙,潘开玉,等.矮牡丹传粉生物学的初步研究[J].植物分类学报,1998,36(2):.
[32]骆劲涛,2013.四川牡丹的传粉生物学研究[M].成都:四川农业大学(硕士学位论文).
[33]马莘,夏颖,马庆庆,等.四川牡丹群落物种生态位及空间关联度分析[J].重庆师范大学学报(自然科学版),2011,28(4):57~61.
[34]孟丽,郑国生.部分野生与栽培牡丹种质资源亲缘关系[J].林业科学,2004,40(5):110~115.
[35]潘开玉,温洁,罗毅波,等.矮牡丹小孢子发生和雄配子体发育及其与该种濒危的关系[J].植物分类学报,1999,37(3):244~252.
[36]宋春花,2011.芍药杂交的细胞学研究[M].济南:山东农业大学(硕士学位论文).
[37]苏建荣,刘万德,郎学东,等.濒危植物大花黄牡丹与生境地群落特征的关系[J].林业科学研究,2010,23(4):487~492.
[38]覃曼,冯铁辛,杜亚然.不同历史时期牡丹种植与地域社会发展——以四川盆地区域为中心的考察[J].西南农业大学学报(社会科学版),2010,8(6):88~91.
[39]唐红,张亮,刘文兰,等.甘肃子午岭野生紫斑牡丹种群及生境[J].东北林业大学学报,2012,40(5):51~58.
[40]王兵益,王伟,丁开宇.滇牡丹花粉贮存方法的探索[J].云南大学学报(自然科学版),2001,(S1):.
[41]王士泉,李慧敏,王腊,等.大花黄牡丹花粉育性研究[J].广东农业科学,2012,23:30~31.
[42]王越岚.牡丹的杂交育种及组间杂种育性的研究[M].北京:北京林业大学(硕士学位论文),2009.
[43]肖佳佳.芍药属杂交亲和性及杂种败育研究[M].北京:北京林业大学(硕士学位论文),2010.
[44]杨辉,戴林森,史国安.观赏及药用牡丹的结实力、种子特性及生化成分的观测分析[J]. 河南科技大学学报(自然科学版),2006,27(5):76~78.
[45]杨霁琴.滇西北4种扁蕾属植物主动自交机制及繁殖分配策略研究[M].中国科学院大学 (硕士学位论文),2013.
[46]张振中,邵律,高毅.中国林业经济学会李育才理事长:油用牡丹增收送健康[J].上海经济,2014,(4):26~28.
[47]赵宣.芍药属牡丹组(Paeonia sect.Moutan)种间关系及栽培牡丹的起源[M].重庆:西南大学(博士学位论文),2007.
[48]赵一鹏,蔡祖国,李本勇.珍稀濒危植物矮牡丹研究进展[J].河南农业科学,2009,(7):14~17.
[49]周丽华,蔡秀珍,张宏亮.珍稀濒危植物的濒危机制与保护对策[J].湖南人文科技学院学报,2006,(6):43~46.
[50]周学刚,张丽萍,王艳芳,等.白芍原植物的传粉特性研究[J].中国农学通报,2010,26(14):177~181.
