基于植被群落水分自营作用的干热河谷植被恢复技术初探
2015-12-02张春梅王东睿黄雪菊
田 雨,张春梅,谢 强,王东睿,黄雪菊
(1.四川省环境保护科学研究院,四川成都 610041;2.阿坝藏族羌族自治州环境监测中心站,四川马尔康 624000)
1 概述
我国是世界上干旱河谷分布最集中的国家之一,主要分布于金沙江、怒江、澜沧江和雅砻江的中、下游,大渡河和元江的中游,岷江上游和嘉陵江上游的白水河等河谷两岸海拔1 600 m以下的区域。其范围主要包含四川和贵州的西北部、西藏西南部和云南部分地区。干旱河谷通常分为干热河谷、干暖河谷和干温河谷3种类型(张荣祖,1992),干热河谷是其中生态环境最恶劣、植被最难以恢复的区域。
长期以来,人们试图在干热河谷按照一般林区造林,通过人工栽植,短期内使稀树灌草丛变成森林,虽然不惜一切代价人工补充水分和养分,在居民点附近可以种植一些树木,甚至在一些试验区、样板田,在人工浇灌情况下,也可营造数公顷的小片林地,但并没有达到在广大山丘坡地成林的目的。近年来,科研工作者加大了干热河谷区植被恢复技术的研究力度,试图找到一种新的技术来解决干热河谷区的植被恢复的难题。目前的研究工作主要集中于抗旱机制和抗旱物种挑选、保水保墒技术研究、造林模式研究、植被恢复新技术研究等方面。
近些年来,虽然我国投入了大量资金用于干热河谷区的植被恢复的研究和实践,但是效果一直不好,形成了“年年种树不见树,年年造林不成林”的现状。究其原因,主要是因为以前的方法大多是通过人工促进作用,增加植被的水分供应,以满足植被生长的水分需求,一旦人工作用消失,而植被还未能适应干旱环境,植被便会死亡。举例来说,金沙江干热河谷种植的多种乔木,在幼苗和幼龄期一般可存活,到成林时期则因土壤水分和养分不足而相继死亡。据20世纪70年代在云南省元谋县直播的云南松(Pinus yunnanensis)、思茅松(Pinus langbianensis)观测,干热河谷中种植的松树,第6年自然稀疏到1 050株·hm-2~6 750株·hm-2,第8年锐减到450株·hm-2~1 050株·hm-2,直到第18年后还在继续减少(费世民等,2003)。由此可见,在干热河谷区,通过人为方式外在补充水分来促进植被生长的造林模式是不可取的。
植被群落具有适应环境和改造环境的功能,要想在干热河谷区取得植被恢复的成功,就应该充分发挥植被改造环境的主观能动性。本文就是基于这一基本原理提出了干热河谷区植被恢复的新思路,并设计试验对该思路进行了验证。本研究工作可以为干旱区植被恢复和重建提供理论基础,对改善区域生态环境质量、促进社会经济持续发展具有重要意义。
2 理论基础
2.1 植被群落的水分自营作用
植被群落在适应区域地理环境和气候条件的同时,还具备通过生命过程对所在地的自然环境进行改造的功能。水分作为植被生长不可或缺的一个因素,植被群落必然会通过自身来改造水分环境(水分条件)以满足自身生长的需求,我们把植被群落的这种功能叫做植被群落的水分自营作用。
2.1.1 冠层对水分的调节作用
(1)冠层反射
植物在漫长的进化史中形成了一种有利于光合作用的光谱响应特性,植物叶面产生出对有效光合波段(可见光)高吸收(低反射率)和对无效光波段(热红外)高反射的特征。植物在可见光谱区(300 nm~600 nm波段)的反射系数较小(约0.1,与土壤的反射率相当),而在近红外光波段却存在反射率跃增,即从700 nm的波长开始,植物叶面的反射系数陡然增大到0.4~0.6,而且无论植物的种类和枝叶结构如何,都存在这种反射率的跃升,而水体、土壤或岩石则不具有这个特性。植物光合作用只能利用太阳光谱中的高能(短波)部分,低能(红外)光子达不到光合作用的量子阈值。而且红外光还有强烈的热效应,可引起植物体温升高,呼吸和异化代谢加强,不利于植物生长和营养物的积累。植物增大红外光反射率可有效地避免这种不利作用,获得最佳光能利用效益。
(2)林冠截留
林冠的截留作用对森林生态系统水分收入和对土壤水分的补充有着显著的影响。通常,截留的水分将用于森林生态系统的蒸散发,减少林地土壤水分损失,提高土壤水分含量。林冠层对雨水的截留和缓冲作用,会显著减少进入林地的水量,导致地表径流减少。
(3)林冠阻抗
林冠的阻抗作用主要体现在气孔阻抗和边界层阻滞作用方面。植物根系从土壤吸收水分,经导管向上移动,在根压和蒸腾拉力作用下,水分移动可达到树梢的叶子。叶子表皮有很多气孔,这些气孔在两个保卫细胞之间,水分进入保卫细胞时,细胞膨胀,气孔张开,散发开始进行。如果水分减少至一定程度,保卫细胞松弛,气孔关闭。这会减少水分的支出。影响气孔开闭的主要因素是光线强弱、叶的水分补给、空气的温度、湿度和生物化学变化。通常,气孔白天开着,夜晚关闭。温度小于4.5°C时,散发很少。很多的研究结果表明,在受到外界干旱胁迫时,植物会关闭气孔,增大水分输出阻抗,直接减少水分的输出。
此外,由于林冠边界粗糙度大,组织层次丰富,林冠还能降低风速,阻碍水汽交换,抑制对流扩散,降低林地的水分蒸散发。
2.1.2 枯落物对水分的调节作用
(1)截留和滞留作用
森林枯枝落叶层具有较大的水分截留能力。Miller(1977)研究表明森林植被和枯落物层都可以贮存1 kg·m-2~3 kg·m-2的降水。各种森林枯落物的最大持水率平均为自身干重的309.54%,林地枯落物层的最大持水量平均为4.18 mm(刘世荣等,1996)。枯落物层对土壤蓄水量有显著的影响。枯落物层本身具有良好的吸持水分和涵养水分的性能,在降雨季节能吸持大量的降水,同时能将多余的水分缓慢的渗透到地下,增加土壤蓄水能力;在干旱季节中,可将自身涵养的水分缓慢下渗,增加土壤水分,保持土壤湿度。枯落物吸收的水分主要用于林地的蒸发,如此一来,林地土壤的蒸发量会相对小于裸地,有利于林地保持土壤湿度。
(2)增加土壤入渗量
当降水量超过枯枝落叶层饱和持水量或降水强度超过其临界持水能力又不及入渗土壤时,林地坡面便产生径流。径流流经枯枝落叶层时,受到阻力,流速减小,水分下渗的时间延长,土壤入渗量增加,增大了森林生态系统的可利用水分,这对改善森林地水分收支平衡十分有利。
(3)抑制土壤蒸发
覆盖是一种古老的蓄水保墒措施,在农业和林业上得到了广泛应用。枯枝落叶层作为一种自然的覆被,不但能吸收太阳辐射,减少林地土壤所受的直接辐射,降低土壤温度,还能阻滞土壤和大气间的水汽交换,从而抑制土壤的蒸发。土壤蒸发是地气能量交换中的主要过程之一,它既是地表能量平衡的组成部分,又是水分平衡的组成部分(孟春雷,2007)。蒸发作用造成土壤含水量减少,影响植物正常生长。枯落物的存在有效抑制了林内土壤蒸发,保持了林地土壤水分,改善了土壤小环境条件,提高了水分的利用率。
(4)改善土壤性质
枯落物层对林地土壤的水分物理性质也有影响。它能使土壤变疏松,改善土壤温度,防止土壤严重冻结。在降雨的过程中还能起到保护、过滤作用,有效地使土壤免受雨水的直接冲击,同时还能过滤出致密的土壤颗粒,防止土壤因为大孔隙的堵塞,而减弱水分渗透的性能。
2.1.3 植物根系对水分的影响
植被根系对水分的调节作用主要是通过改善土壤性能来间接实现的。这种调节作用是通过根系对土壤的固持以及根系对土壤物理性状和结构的改善来实现的。
在浅层土壤中,林木的侧根组成了连续的和具有斜向牵引力作用的根网,对根际土层的强度有重要意义,小直径的侧根组成的密集根网如同具有斜向抗张强度的张力膜,既加固根际土层,又把下次土壤固持在原有位置(周跃,1999)。林木根系对土体的固持力主要决定于土壤中根表面积和根的抗拉力的大小。在黄土坡地上,林木根系与土壤间的静摩擦阻力是决定根系提高土壤抗剪强度增量的主要因素,长而粗的少量根系与土体间的摩擦阻力远大于短而细的多量根系。许多木本植物的主根直径粗大,根系深入土层可达3 m~5 m,对于稳固土体环境具有很大的积极作用。有的学者则认为土壤抗冲性强度值与小于1 mm径级的须根密度关系最密切,并将土壤剖面中100 cm2截面上小于1 mm须根数作为有效根系密度的指标(余新晓等,2004)。
2.2 基于植被群落水分自营作用的植被恢复技术
上述分析表明,植被群落的水分自营作用是通过植被自身、下垫面(主要是枯落物)、土壤(物化性质、微生物、土壤动物)等综合实现的(程根伟&田雨,2013)。要充分发挥植被的水分自营作用,就必须从植被、土壤、下垫面这几个方面进行改造:
(1)物种选择:选择耐旱物种,优先选择本地优势物种;
(2)土壤改良:一方面需要确定土壤厚度能够满足成熟林必需的土壤厚度需求,如土壤厚度不够,需要采取措施增加土壤厚度;另一方面,需要改善土壤物化性质,采用施肥等方式,提高土壤的保水能力;
(3)下垫面改造:植被栽植后,要在土壤表层覆盖一层枯落物或枯草,起到降低水分蒸发、增强水分蓄积和改良土壤的作用。
3 实践探索
3.1 研究区域
试验在四川省环境保护科学研究院西蒙得木基地内进行。基地位于位于四川省凉山彝族自治州南端会东县境内,总面积66.7 hm2,中心坐标为东经102°34'39.18″,北纬 26°22'34.25″。基地位于金沙江干热河谷内,干热少雨,雨季(5月~10月)降水量占年降水总量的90%以上。
3.2 试验方法
3.2.1 目标物种的选择
会东基地周围的优势植被为西蒙得木、合欢、车桑子和黄茅。其中,西蒙得木(Simmondsia chinensis L.)是一种多年生常绿灌木,原产美国西南部的加利福尼亚州、新墨西哥州、亚利桑那州和墨西哥北部热带沙漠。西蒙得木具有的极强抗旱特性,能在年降雨量仅125 mm的地区生长。西蒙得木的根系极其发达,主根可深入地下十几米,是理想的固土固沙的耐旱植物,且其树龄可长达200年左右,故有“沙漠克星”的誉称(徐亮,2006)。在会东基地,西蒙得木的生长状况要好于其物种。因此,本研究选取西蒙得木作为探索试验的目标物种。
3.2.2 验证试验
为了探明本文提出的植被恢复技术的可行性,设计了1 a生西蒙得木幼苗栽植和5 a生西蒙得木苗木移植两个试验。
幼苗栽植:栽植试验在会东西蒙得木基地的一处坡地上进行。该坡地坡度约为25°,土壤厚度约10 cm~25 cm。前些年曾进行过西蒙得木的栽种,但最终以失败告终。原来的坡地土层较薄,不适合植被的生长,为了满足西蒙得木幼苗的生长需求,需要进行坡改梯,坡改梯后土层厚约40 cm。幼苗栽植在7月雨季进行。栽种结束后在土壤表层再覆盖一层5 cm厚的枯落物层。栽种最初1个月内,补充一定的水分。在后期,不再补充水分。
成林苗移植:移栽试验在会东西蒙得木基地的一处缓坡地上进行。该坡地坡度约为5°~8°,土壤厚度大于80 cm。栽植方法为:在样地内采用块状清林后,挖穴60 cm×60 cm×60 cm,将基地内10年龄的西蒙得木移栽入穴,施基肥3 kg·穴-1。栽植结束后在表层覆盖10 cm的枯落物。
对照:为了对比试验效果。在基地内选取了一块坡度约为25°,土壤厚度约20 cm的裸地进行了幼苗栽植。栽植前不进行土壤改良,栽植后覆盖3 cm厚的枯落物层。除了栽种最初1个月内补充一定的水分外,不再补充水分。
3.3 试验观测
观测指标主要包括:样地土壤水分含量和植被生长状况。土壤水分用便携式土壤水分仪测定;植被的生长状况用植被的成活率来表征。
4 结果
4.1 土壤水分
观测结果表明,土壤水分含量呈现出移栽西蒙得木林地>西蒙得木幼苗林地>裸地的规律(图1)。
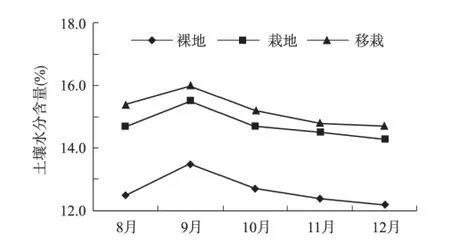
图1 裸地、栽植林地和移栽林地的土壤水分含量Fig.1 SWC of bare land,planting woodland and transplanting woodland
4.2 植被成活率
不同处理方式下西蒙得木的成活率有明显差距。对照组的西蒙得木成活率逐渐减低,栽植的西蒙得木幼苗在栽植后3个月内成活率稍有下降,然后趋于稳定;移栽的西蒙得木则完全成活(图2)。
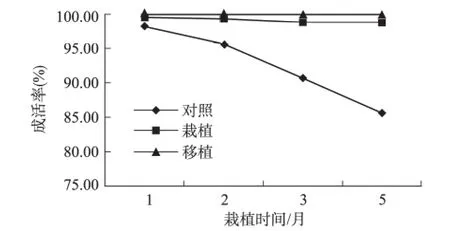
图2 植被的成活率随栽植时间的变化Fig.2 The survival rate of vegetation in bare land,planting woodland and transplanting woodland
5 讨论
在干热河谷区,水分是制约植被生长的关键因素。要解决水分亏损的难题,一方面要增加水分供给,另一方面则需要减少水分的无效消耗。人工浇灌是增加水分供给的最直接的方式,然而,一旦停止供给,植被则会因为无法克服干旱环境而死亡。因此,人工灌溉的方式只能短期的解决水分亏损问题,不能从根本上解决问题从而达到植被恢复的效果。
植被具有改造和适应环境的本能,能发挥自身的能动性调节水分,达到增加水分供给、减少无效水分消耗、高效利用水分以满足自身生长的目的。植物调节水分的功能除了自身以外,还受外界环境的影响。土壤和下垫面(主要是枯落物层)是影响这种调节作用发挥最关键的因素。
本文从土壤改良和表层枯落物覆盖两个方面着手,为西蒙得木的生存营造了良好的生存条件,使其能充分发挥“自营”水分的功能,从而克服干旱环境而得以成活。研究结果表明,经改造后,无论是西蒙得木幼苗栽植还是5 a龄成熟苗移栽都能取得良好的效果。
6 结论
本文基于植被群落的水分自营作用原理提出了干热河谷植被恢复的技术方法:充分利用植被改造环境和适应环境的本能,在物种遴选的基础上,侧重从土壤改良和下垫面营造两个方面着手,为植被的生存营造适当的生存条件,使其能充分发挥“自营”水分的功能,从而克服干旱环境而得以成活。结果表明,本文提出的干热河谷植被恢复新技术可行。
[1]费世民,王鹏,陈秀明,等.论干热河谷植被恢复过程中的适度造林技术[J].四川林业科技,2003,24(3):10~16.
[2]刘世荣,温远光,王兵,等.中国森林生态系统水文生态功能规律[M].北京:中国林业出版社,1996.
[3]孟春雷.土壤蒸发及水热传输研究综述[J].土壤通报,2007,38(2):374~378.
[4]徐亮.西蒙得木对金沙江干热河谷生态保护的作用[J].资源开发与市场,2006,22(5):452~453,463
[5]余新晓,张志强,陈丽华,等.森林生态水文[M].北京:中国林业出版社,2004.
[6]张荣祖.横断山区干旱河谷[M].北京:科学出版社,1992.
[7]钟祥浩.干热干旱河谷区生态系统退化及恢复与重建途径[J].长江流域资源与环境,2000,9(3):376~383.
[8]周跃.乔木侧根对土体的斜向牵引效应:I原理和数学模型[J].山地学报,1999,17(1):4~9.
