低氧条件下铁调节蛋白1在维持SH-SY5Y细胞铁稳态中的作用
2015-11-21郁敏燕
郁敏燕,王 丹,朱 俐
(1.南通大学附属医院生殖医学中心,江苏南通226001;2.南通大学航海医学研究所,江苏南通226001)
低氧条件下铁调节蛋白1在维持SH-SY5Y细胞铁稳态中的作用
郁敏燕1,2,王 丹2,朱 俐2
(1.南通大学附属医院生殖医学中心,江苏南通226001;2.南通大学航海医学研究所,江苏南通226001)
目的:研究低氧条件下铁调节蛋白1(iron regu1atory protein 1,IRP1)对人神经母细胞瘤SH-SY5Y细胞铁稳态的影响。方法:1%O2处理SH-SY5Y细胞0,1,2,4,6 h,蛋白质印迹法检测铁蛋白轻链(ferritin 1ight chain,Ft-L)以及膜铁输出蛋白(ferroportin,Fpn)的表达;Ca1cein-AM法检测SH-SY5Y细胞对Fe2+的吸收以及细胞可变铁池(1abi1e iron poo1,LIP);采用硫氰酸盐分光光度法测定细胞总含铁量;特异性siRNA下调IRP1后检测SH-SY5Y细胞低氧2 h铁稳态的变化。结果:低氧0~6 h,SH-SY5Y细胞Ft-L和Fpn表达增加,Fe2+摄入和LIP增加,但细胞总铁水平不变。siRNA下调IRP1表达后再低氧2 h,Ft-L蛋白表达和LIP明显增加,Fpn表达和Fe2+摄入没有明显变化,细胞总铁水平增加。结论:低氧条件下,IRP1是SH-SY5Y细胞铁稳态的重要调节者。
低氧;铁调节蛋白1;铁稳态;SH-SY5Y细胞;细胞可变铁池
铁为生命活动所必需,其缺乏会引起细胞生长停止和死亡,超载可诱发大量自由基的产生,引起神经退行性病变和多种与老年相关的疾病。游离铁通过Haber-Weiss或Fenton反应与过氧化氢或脂质过氧化物反应促进活性氧簇的产生,导致脂质、蛋白质和DNA的损伤[1]。细胞通过铁调节蛋白(iron regu1atory proteins,IRPs,包括IRP1和IRP2)转录后调节机制,即与多种编码铁存储、摄取、转运和输出的蛋白mRNA非翻译区(untrans1ated regions,UTRs)的铁反应元件(iron responsive e1ements,IREs)结合,调节细胞铁稳态[2-4]。
在原核生物和真核生物中,铁蛋白(ferritin,Ft)是细胞内主要的铁存储蛋白[5]。铁蛋白通常由重链(Ft-H)和轻链(Ft-L)的24个亚基组成,Ft-L促进Fe2+沉积并稳定铁蛋白结构[6]。IRPs通过与铁蛋白mRNA 5′-UTRs的IREs结合抑制铁蛋白翻译,下调其表达,增加细胞可变铁池(1abi1e iron poo1,LIP)的Fe2+以供细胞代谢[7]。膜铁输出蛋白(ferroportin,Fpn)是目前已知的唯一的细胞膜铁输出蛋白,在细胞(巨噬细胞、肝细胞和胎盘细胞)表面具有很强的铁输出能力,呈高表达[8]。Fpn的表达和功能可通过低氧诱导因子(hypoxia-inducib1e factor,HIF)受低氧调控,也可能受IRPs调控。
机体铁稳态的维持除了需要多种蛋白的协调作用外,还有赖于适当的氧供给。研究表明,低氧应激会引起机体产生一系列基因表达的变化,以适应一定程度的低氧[9-10],减少其危害。HIF-1是低氧诱导基因表达的主要调节因子,对低氧适应起关键性作用[11]。HIF-1是一个异源二聚体,包含两个螺旋-环-螺旋蛋白,HIF-1a为调节亚基,是氧敏感元件;HIF-1β是已知的芳烃受体核转位分子,呈组成型表达。常氧下,HIF-1通过泛素蛋白酶途径降解。低氧时,HIF-1a亚基在胞质中集聚,通过核孔进入胞核与HIF-1β亚基组装成HIF-1,与靶基因的低氧反应元件(hypoxia responsive e1ement,HRE)结合。我们前期研究发现,在人肝癌HepG2细胞中,低氧通过HIF/HRE系统下调IRP1表达,证明IRP1存在HRE[12],但低氧下IRP1自身的调控作用尚不清楚。
研究显示,阿尔茨海默病、帕金森病、亨廷顿舞蹈病的基底神经节等多个脑区铁含量和铁代谢相关蛋白明显提高,提示神经细胞的铁含量增高和铁沉积与神经退行性疾病高度相关[13-14],脑铁调节的失衡很可能是神经退行性疾病的发病原因之一[15-16]。但是对铁代谢相关蛋白在神经细胞的表达调控了解不多,尤其对神经细胞应对低氧缺血的表达调控知之甚少。为此,本研究拟通过人神经母细胞瘤SHSY5Y细胞探讨低氧对神经细胞铁稳态的影响以及该过程中IRP1的作用,以了解低氧应激对神经细胞铁稳态的影响以及神经细胞在适应内环境低氧时对自身铁稳态的调节过程。
1 材料与方法
1.1 细胞培养和低氧诱导
人神经母细胞瘤SH-SY5Y细胞(购自中国科学院细胞库)用含10%胎牛血清的DMEM(Gibco公司),在37℃,5%CO2条件下培养。低氧处理之前把含10%胎牛血清的DMEM换成无血清的培养基,置于InvivO2200低氧工作站(英国Ruskinn Techno1ogies公司),在37℃,5%CO2,1%O2条件下培养。
1.2 蛋白质印迹法检测
提取常氧组(低氧0 h)和低氧不同时间(1,2,4,6 h)组SH-SY5Y细胞总蛋白,BCA试剂盒(Pierce公司)行蛋白定量,上样30 μg,行10%SDS-PAGE,转至PVDF膜(Mi11ipore公司)。用含5%脱脂奶粉的TBST室温封闭2 h,一抗HIF-1(1∶250,Novus Bio1ogica1s公司)、IRP1(1∶250,Santa Cruz公司)、Ft-L(1∶1 000,Abcam公司)、Fpn(1∶300,A1pha Diagnostic公司)、β-肌动蛋白(1∶10 000,Sigma公司)孵育,4℃过夜,TBST洗3遍,然后二抗(1∶10 000,Jackson公司)室温孵育2 h,TBST洗3遍。ECL(Pierce公司)化学发光显色,X光片显影、定影。胶片拍照,摄取图像(美国UVP公司EC3 Imaging System),Quantity One分析软件(Bio-Rad公司)对条带行定量分析。
1.3 siRNA干扰实验
消化收集SH-SY5Y细胞,重悬浮于电转液中,密度为2.5×107个/mL。将人IRP1 4个特异性siRNA序列加到200 μL电转液中,含5×106个细胞,电穿孔转染干扰片段。室温静置5 min,将电转细胞悬液转移至含10%胎牛血清的DMEM中继续培养48 h,检测低氧2 h未干扰组(对照组)与干扰IRP1组(siIRP1组)Ft-L和Fpn,IRP1蛋白表达。
siRNA序列如下。siRNA 1正义链:5′-UUUCACAACAUGCGGAUUAdtdt-3′,反义链:5′-UAAUCCGCAUGUUGUGAAAdtdt-3′;siRNA 2正义链:5′-AAAGAUAUCUGGCCGACUAdtdt-3′,反义链:5′-UAGUCGGCCAGAUAUCUUUdtdt-3′;siRNA 3正义链:5′-GGGCAAAUUUGUCGAGUUCdtdt-3′,反义链:5′-GAACUCGACAAAUUUGCCCdtdt-3′;siRNA 4正义链:5′-GGCCUAACUCCAUGAGAAUdtdt-3′,反义链:5′-AUUCUCGUGGAGUUAGGCCdtdt-3′。
1.4 Fe2+吸收以及细胞LIP的测定
常氧(低氧0 h)与低氧处理不同时间(2,4,6 h)以及低氧2 h条件下未干扰IRP1(对照组)与干扰IRP1(siIRP1组)的细胞分别用PBS洗3遍,收集(1~3)×106个细胞,悬于1 mL HBSS。将荧光剂Ca1cein-AM(终浓度为0.125 μmo1/L)与细胞悬液于37℃共孵育10 min。用PBS洗去胞外探针,用2 mL不含Ca1cein-AM的HBSS重悬细胞,轻轻吹匀,细胞悬液转入带有微型磁力搅拌子的比色皿中,用荧光分光光度计(Shimadzu公司)检测Ca1cein-AM的荧光(激发波长:488 nm,发射波长:518 nm),记录荧光基线值[17]。测定细胞对Fe2+的吸收时,当基线走平后,加入终浓度为40 μmo1/L硫酸亚铁铵,淬灭荧光,每隔5 min检测1个荧光值并记录,共30 min。测细胞LIP量时,当基线走平后,加入终浓度为500 μmo1/L去铁酮,连续记录荧光值1 000 s[18]。
1.5 硫氰酸盐分光光度法测定细胞总铁含量
消化、计数、收集常氧(低氧0 h)与低氧处理不同时间(2,4,6 h)以及低氧2 h条件下未干扰IRP1(对照组)与干扰IRP1(siIRP1组)的细胞,60℃烘干。加200 μL浓盐酸60℃消化2 h,再加800 μL H2O2于60℃消化2 h。然后取75 μL样品消化液至96孔板中,每孔加入75 μL染色液(0.08%K2S2O8,8%KSCN和3.6%HC1),室温反应10~15 min,于490 nm波长下测定D值[19]。
1.6 统计学分析
应用Sigma Stata 2.0统计软件,实验数据均以均数±标准差(±s)表示,每组取3次独立实验结果统计。采用单因素方差分析对实验结果进行统计分析,以P<0.05为差异有统计学意义。
2 结果
2.1 短时程低氧条件下铁稳态的维持
蛋白质印迹结果显示,与常氧(低氧0 h)相比,低氧处理1,2,4,6 h后,Ft-L和Fpn表达均呈时间依赖性增加(图1)。
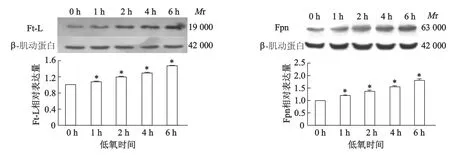
*:P<0.05,与常氧组(低氧0 h)相比图1 低氧对SH-SY5Y细胞Ft-L和Fpn蛋白表达的影响
SH-SY5Y细胞低氧不同时间(2,4,6 h)后,细胞Fe2+摄入均较常氧增加,低氧2 h Fe2+摄入即达峰值,随后逐渐减少(图2A);与常氧(即低氧0 h)相比,低氧2 h LIP增加达最高峰,而后逐渐减少(图2B);然而SH-SY5Y细胞总铁水平基本不受短时程低氧的影响(图3)。
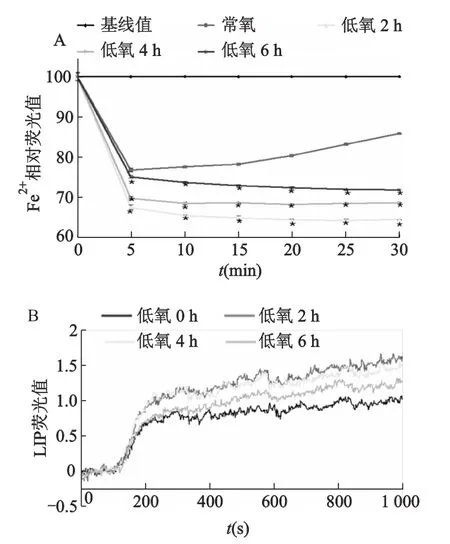
*:P<0.05,与常氧比较图2 低氧对SH-SY5Y细胞Fe2+摄入和LIP的影响
2.2 低氧条件下HIF-1和IRP1表达成相反趋势
结果如图4显示,不同低氧时间处理(1,2,4,6 h)后,SH-SY5Y细胞内HIF-1蛋白表达量均明显高于常氧组(低氧0 h),低氧2 h HIF-1表达达最高峰,约比常氧组高50%。
IRP1蛋白表达先降后升,低氧2 h表达最低,约降低30%,随着低氧时间的延长(4 h和6 h),IRP1蛋白表达逐渐上升。低氧6 h以内,HIF-1蛋白表达和IRP1蛋白表达呈相反趋势,尤其低氧2 h,HIF-1蛋白表达最高而IRP1蛋白表达最低;由此表明,低氧2 h可能是细胞内部分蛋白表达改变的一个重要拐点。
2.3 下调IRP1表达影响低氧条件下细胞的铁平衡RNA干扰组IRP1的蛋白表达较对照组降低约55%,表明siRNA能够下调IRP1的表达。同时,我们还发现在低氧2 h的条件下,SH-SY5Y细胞干扰IRP1蛋白表达后,Ft-L蛋白表达明显增加了20%(P<0.05),而Fpn表达没有明显变化(P>0.05)。见图5。
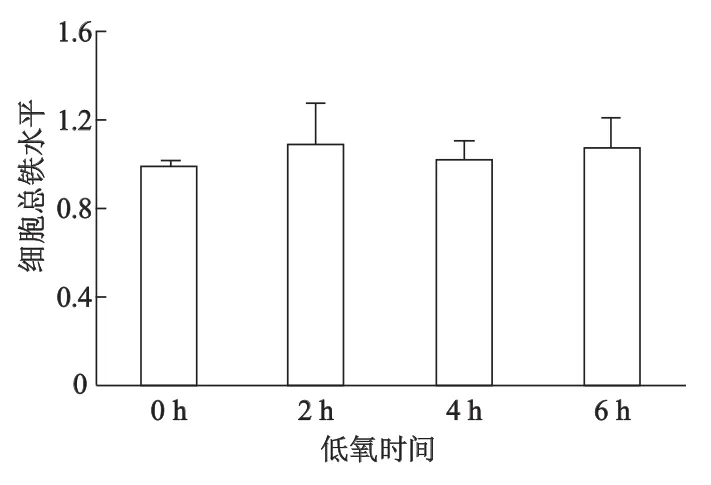
图3 低氧对SH-SY5Y细胞总铁水平的影响

*:P<0.05,与常氧组(低氧0 h)相比图4 低氧下SH-SY5Y细胞HIF-1和IRP1的蛋白表达
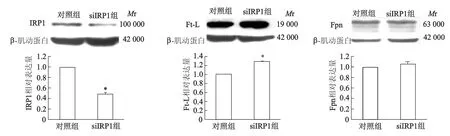
*:P<0.05,与对照组相比图5 低氧2 h干扰IRP1对SH-SY5Y细胞Ft-L和Fpn蛋白表达的影响
此外,下调IRP1表达,低氧2 h对SH-SY5Y细胞Fe2+的摄入没有影响,LIP明显减少50%左右,见图6;siIRP-1组细胞总铁水平较对照组增加了20%(P<0.05,图7)。
3 讨论
研究证实,在低氧6 h条件下,HepG2细胞总铁和Fe2+水平较常氧条件下明显增加[20]。本研究结果显示,在低氧条件下,Ft-L和Fpn水平呈时间依赖性增加。在组织缺氧和细胞低氧模型中,铁蛋白的表达均会发生变化。进行性低氧能诱导新生大鼠和人少突胶质细胞中铁蛋白合成[21-23]。低氧能增加SH-SY5Y细胞Fe2+摄入和LIP,这与铁的吸收是LIP的主要来源相一致[24]。在本研究中,我们发现SH-SY5Y细胞低氧2 h Fe2+摄入比低氧4 h和6 h多,可能是由于Fe2+摄入是一个持续的动态过程,胞内铁的积聚也随时间推移越来越多,因而反馈性抑制Fe2+摄入和LIP。此外,持续的Fe2+摄入可导致Ft-L隔离的铁增多,故Ft-L蛋白表达随低氧时间延长而增加。此外,本研究中细胞Fpn水平逐渐增加,表明细胞铁输出也呈时间依赖性增多,以维持细胞的总铁水平。因而,短时程(6 h)低氧条件下,SH-SY5Y细胞的铁稳态能保持稳定。研究报道低氧能增加人肺泡巨噬细胞和A549细胞铁蛋白的表达但细胞铁含量保持不变[25],本研究结果与其类似。
哺乳动物细胞铁稳态受IRPs转录后调节[1],在人大脑中主要是IRP1起作用[26]。我们前期研究表明,IRP1受低氧调节,HepG2细胞物理或化学低氧6 h IRP1表达下调[12]。研究表明IRP1对低氧和再氧合的反应呈细胞特异性[27]。本研究结果显示,SH-SY5Y细胞低氧处理2 h,Fe2+摄入和LIP增加最多,HIF-1a蛋白表达达峰值,IRP1蛋白表达达最低,提示1%低氧2 h对SH-SY5Y细胞的影响最明显。故我们以低氧2 h为关键点,研究IRP1对SH-SY5Y细胞铁稳态的影响。

图6 低氧2h,干扰IRP1对SH-SY5Y细胞Fe2+摄入和LIP的影响
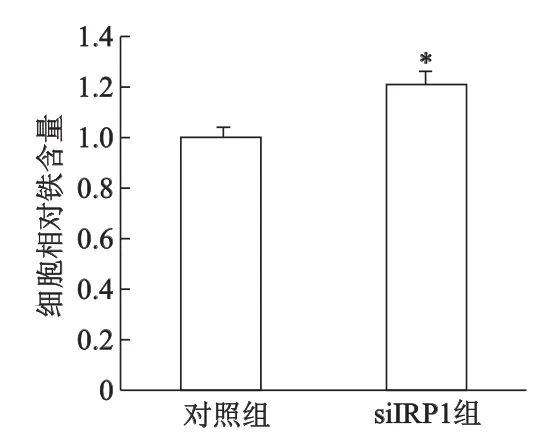
*:P<0.05,与对照组相比图7 低氧2h干扰IRP1对SH-SY5Y细胞总铁水平的影响
虽然短时程(<6 h)低氧对SH-SY5Y细胞的总铁水平没有明显影响,但改变了铁的摄入、输出和存储,由此推测正是通过IRP1对这些铁代谢相关蛋白的调控,参与了短时程低氧下SH-SY5Y细胞铁稳态的维持。下调SH-SY5Y细胞中IRP1表达后,低氧2 h条件下,Ft-L表达增加,LIP明显减少,推测细胞LIP因被铁蛋白隔离的铁增多而减少;Fe2+摄入没有明显变化,Fpn表达也没有变化即铁外流没有变化,但持续的Fe2+摄入可增加细胞总铁含量,故细胞总铁水平有明显增加。提示低氧条件下,如果没有足够IRP1蛋白的参与,细胞铁摄入、输出和存储的方式均会发生改变,从而影响细胞的铁稳态。但我们只用siRNA的方法研究了低氧条件下IRP1在维持细胞铁稳态过程中的作用,IRP1对细胞铁稳态的影响及机制仍需作深入研究。低氧条件下细胞Fe2+摄入的增多提示可能有铁吸收蛋白表达的增加。二价金属离子转运体(DMT1)和转铁蛋白受体(TfR1)是两种重要的铁吸收蛋白,二者在低氧条件下都受HIF-1的转录调节[20],对铁浓度改变的反应也均受IRP1转录后调节[28]。在低氧条件下如何通过HIF-1/HRE和IRP1/IRE体系协同调节DMT1/ TfR1来影响铁的吸收有待进一步研究。
细胞铁稳态受IRP1和IRP2转录后调控。当氧含量丰富时,IRP1与铁代谢相关蛋白mRNAs的UTRs的IREs结合,调节铁稳态,而IRP2经蛋白酶体途径被快速降解[29]。而低氧时,IRP2是与IREs结合的主要蛋白,结构稳定[30]。低氧条件下,IRP1表达下调对SH-SY5Y细胞铁稳态的影响可能是由于IRP2的代偿性作用。由于IRP2也是主要的铁调节蛋白,低氧条件下IRP2在维持细胞铁稳态过程中的作用值得进一步研究。
综上所述,短时程低氧(<6 h,1%O2)条件下,SH-SY5Y细胞铁稳态保持稳定。低氧2 h,当IRP1下调后,由于细胞总铁含量增加,SH-SY5Y细胞铁稳态发生改变。故低氧条件下,IRP1在SH-SY5Y细胞铁稳态的调节过程中起重要作用。
[1] Ha11iwe11 B,Gutteridge JM.Ro1e of free radica1s and cata1ytic meta1 ions in human disease:an overview[J]. Methods Enzymo1,1990,186:1-85.
[2] Hentze MW,Kühn LC.Mo1ecu1ar contro1 of vertebrate iron metabo1ism:mRNA-based regu1atory circuits operated by iron,nitric oxide,and oxidative stress[J].Proc Nat1 Acad Sci U S A,1996,93(16):8175-8182.
[3] Eisenstein RS.Iron regu1atory proteins and the mo1ecu-1ar contro1 of mamma1ian iron metabo1ism[J].Annu Rev Nutr,2000,20:627-662.
[4] Schneider BD,Leibo1d EA.Regu1ation of mamma1ian iron homeostasis[J].Curr Opin C1in Nutr Metab Care,2000,3(4):267-273.
[5] MacKenzie EL,Iwasaki K,Tsuji Y.Intrace11u1ar iron transport and storage:from mo1ecu1ar mechanisms to hea1th imp1ications[J].Antioxid Redox Signa1,2008,10(6):997-1030.
[6] Arosio P,Ingrassia R,Cavadini P.Ferritins:a fami1y of mo1ecu1es for iron storage,antioxidation and more[J].Biochim Biophys Acta,2009,1790(7):589-599.
[7] A1berghini A,Reca1cati S,Tacchini L,et a1.Loss of the von Hippe1 Lindau tumor suppressor disrupts iron homeostasis in rena1 carcinoma ce11s[J].J Bio1 Chem,2005,280(34):30120-30128.
[8] Anderson GJ,Vu1pe CD.Mamma1ian iron transport[J].Ce11 Mo1 Life Sci,2009,66(20):3241-3261.
[9] Thomas JD,Johannes GJ.Identification of mRNAs that continue to associate with po1ysomes during hypoxia[J].RNA,2007,13(7):1116-1131.
[10] Martens LK,Kirschner KM,Warnecke C,et a1.Hypoxia-inducib1e factor-1(HIF-1)is a transcriptiona1 activator of the TrkB neurotrophin receptor gene[J].J Bio1 Chem,2007,282(19):14379-14388.
[11] Ke Q,Costa M.Hypoxia-inducib1e factor-1(HIF-1)[J].Mo1 Pharmaco1,2006,70(5):1469-1480.
[12] Luo QQ,Wang D,Yu MY,et a1.Effect of hypoxia on the expression of iron regu1atory proteins 1 and the mechanisms invo1ved[J].IUBMB Life,2011,63(2):120-128.
[13] Bartzokis G,Tish1er TA,Shin IS,et a1.Brain ferritin iron as a risk factor for age at onset in neurodegenerative diseases[J].Ann N Y Acad Sci,2004,1012:224-236.
[14] Bartzokis G,Tish1er TA,Lu PH,et a1.Brain ferritin iron may inf1uence age-and gender-re1ated risks of Neurodegeneration[J].Neurobio1 Aging,2007,28(3):414-423.
[15] Qian ZM,Shen X.Brain iron transport and neurodegeneration[J].Trends Mo1 Med,2001,7(3):103-108.
[16] Lee DW,Andersen JK,Kaur D.Iron dysregu1ation and neurodegeneration:the mo1ecu1ar connection[J].Mo1 Interv,2006,6(2):89-97.
[17] Breuer W,Epsztejn S,Cabantchik ZI.Iron acquired from transferrin by K562 ce11s is de1ivered into a cytop1asmic poo1 of che1atab1e iron(Ⅱ)[J].J Bio1 Chem,1995,270(41):24209-24215.
[18] Barnabé N,Zastre JA,Venkataram S,et a1.Deferiprone protects against doxorubicin-induced myocyte cytotoxicity[J].Free Radic Bio1 Med,2002,33(2):266-275.
[19] Liu YX,Chen ZP,Wang JK.Systematic eva1uation of biocompatibi1ity of magnetic Fe3O4nanopartic1es with six different mamma1ian ce11 1ines[J].J Nanopart Res,2011,13:199-212.
[20] Wang D,Wang LH,Zhao Y,et a1.Hypoxia regu1ates the ferrous iron uptake and reactive oxygen species 1eve1 via diva1ent meta1 transporter 1(DMT1)exon1B by hypoxia-inducib1e factor-1[J].IUBMB Life,2010,62(8):629-636.
[21] Qi Y,Dawson G.Hypoxia induces synthesis of a nove1 22-kDa protein in neonata1 rat o1igodendrocytes[J].J Neurochem,1992,59(5):1709-1716.
[22] Qi Y,Dawson G.Hypoxia specifica11y and reversib1y induces the synthesis of ferritin in o1igodendrocytes and human o1igodendrog1iomas[J].J Neurochem,1994,63(4):1485-1490.
[23] Qi Y,Jamindar TM,Dawson G.Hypoxia a1ters iron homeostasis and induces ferritin synthesis in o1igodendrocytes[J].J Neurochem,1995,64(6):2458-2464.
[24] Kruszewski M.Labi1e iron poo1:the main determinant of ce11u1ar response to oxidative stress[J].Mutat Res,2003,531(1/2):81-92.
[25] Smith JJ,O′Brien-Ladner AR,Kaiser CR,et a1. Effects of hypoxia and nitric oxide on ferritin content of a1veo1ar ce11s[J].J Lab C1in Med,2003,141(5):309-317.
[26] Hu J,Connor JR.Demonstration and characterization of the iron regu1atory protein in human brain[J].J Neurochem,1996,67(2):838-844.
[27] Irace C,Scorzie11o A,Maffettone C,et a1.Divergent modu1ation of iron regu1atory proteins and ferritin biosynthesis by hypoxia/reoxygenation in neurones and g1ia1 ce11s[J].J Neurochem,2005,95(5):1321-1331.
[28] Sa1ahudeen AA,Bruick RK.Maintaining Mamma1ian iron and oxygen homeostasis:sensors,regu1ation,and cross-ta1k[J].Ann N Y Acad Sci,2009,1177:30-38.
[29] Rouau1t TA.The ro1e of iron regu1atory proteins in mamma1ian iron homeostasis and disease[J].Nat Chem Bio1,2006,2(8):406-414.
[30] Meyron-Ho1tz EG,Ghosh MC,Rouau1t TA.Mamma1ian tissue oxygen 1eve1s modu1ate iron-regu1atory protein activities in vivo[J].Science,2004,306(5704):2087-2090.
Involvement of iron regulatory protein 1 in iron homeostasis under hypoxia in human neuroblastoma(SH-SY5Y)cells
YU Min-yan1,2,WANG Dan2,ZHU Li2
(1.Center for Reproductive Medicine,Affi1iated Hospita1 of Nantong University,Nantong Jiangsu 226001;2.Institute of Nautica1 Medicine,Nantong University,Nantong Jiangsu 226001,China)
Objective:To investigate the effects of iron regu1atory protein 1(IRP1)on iron homeostasis in human neurob1astoma SH-SY5Y ce11s under hypoxia.Methods:The SH-SY5Y ce11s were exposed to hypoxia for 0,1,2,4,6 h(1%O2),and then the tota1 proteins were extracted to detect the expression of ferritin 1ight chain(Ft-L)and ferroportin(Fpn)by Western b1otting.Ca1cein-AM was used to measure ferrous iron uptake and 1abi1e iron poo1(LIP)in SH-SY5Y ce11s.Spectrophotometry was used to determine the tota1 ce11u1ar iron 1eve1 with thiocyanate.The effects of IRP1 siRNA on iron homeostasis in SH-SY5Y ce11s under hypoxia were detected.Results:Iron homeostasis remained unchanged when ce11s were exposed to 1%hypoxia for 0-6 h a1though the expression of Ft-L and Fpn increased,and the 1eve1 of ferrous iron uptake and LIP e1evated.When IRP1 was knocked down using IRP1-specific siRNA,the expression of Ft-L and LIP increased,whi1e Fpn and ferrous iron uptake did not a1tered obvious1y.The iron homeostasis in SH-SY5Y ce11s a1tered under 2 h hypoxia in terms of the ce11u1ar tota1 iron content increased.Conclusion:IRP1 regu1ated iron homeostasis in SH-SY5Y ce11s under hypoxia.
hypoxia;iron regu1atory protein 1;iron homeostasis;SH-SY5Y ce11s;1abi1e iron poo1
R392.2
A
1671-7783(2015)06-0466-06
10.13312/j.issn.1671-7783.y150154
2015-07-08 [编辑] 刘星星
国家自然科学基金资助项目(30971197,31171143,31471141)
郁敏燕(1988—),女,研究实习员,硕士;朱俐(通讯作者),教授,博士生导师,E-mai1:zhu1i1i65@126.com
