杉木全双列子代遗传变异分析
2015-11-17徐清乾吴成田许忠坤
徐清乾, 吴成田, 许忠坤*
(1.湖南省林业科学院, 湖南 长沙 410004; 2.靖州县排牙山国有林场, 湖南 靖州 418400)
杉木全双列子代遗传变异分析
徐清乾1, 吴成田2, 许忠坤1*
(1.湖南省林业科学院, 湖南 长沙 410004; 2.靖州县排牙山国有林场, 湖南 靖州 418400)
对杉木8个无性系全双列杂交组合的32年生成林的树高、胸径和材积作了杂交优势和自交衰退效应分析,并对配合力方差和配合力效应值进行了计算。得出如下结论: 3个性状都存在真实的遗传变异,一般配合力、特殊配合力、正反交效应方差都达到显著水平,且材积以特殊配合力的遗传方差比(GVC)最大。评选出适于杂交的优良无性系3个,适于建立双系种子园的优良无性系2组,适于选作二代优树的优良杂交组合19个。
杉木; 无性系; 全双列杂交; 配合力; 遗传方差
自1974年陈岳武等对杉木配合力育种的研究以来,南方各杉木产区(省)相继开展了双列设计和测交设计育种试验[1-3]。湖南省靖州县大堡子乡因其杉木速生、优质而闻名全国,1972—1974年,在该乡选择杉木一代优树250株,并在省内多处建立初级种子园。作者研究了大堡子乡8个杉木无性系的一般配合力(GCA)、特殊配合力(SCA)和正反交效应(REC),估算了杉木性状受遗传控制的强弱以及3种配合力的相对重要性[4-6],筛选了优良亲本和杂交组合,为省内杉木种子园的去劣和建立高世代种子园提供依据和繁殖材料。
1 试验区概况
试验地位于靖州县新厂乡哨团村,其地理位置109°20′33″E、26°21′28″N。该区地处亚热带季风湿润气候区,其气候温和,雨量充沛,四季分明,夏无酷热,冬少严寒;年平均气温17.0℃,1月平均气温4.9℃,极端低温为-8.4℃;7月平均气温为27.3℃,极端高温为38.4℃;年均降雨量为1300mm,相对湿度82%,≥10℃的年活动积温为4800~4900℃,日照时数1336h,全年无霜期长达290天。造林地为板页岩发育成的红黄壤,土层厚80~100cm,pH值为5.6,海拔高400 m;前茬为白栗、黄竹、马尾松等组成的松杂残林,气候和土壤条件均适宜杉木生长。
试验林面积为2.0hm2,初植造林密度2.2m×2.2m,间伐后保留密度为70株/667m2。
2 材料和方法
2.1 试验材料
1982年3月,在靖州县排牙山杉木初级种子园选择靖2、靖6、靖7、靖13、靖16、靖23、靖31、靖35等8个无性系,采用人工控制授粉的方法,进行64个组合的全双列杂交组配[7]。采摘杂交组合种子64份、参试无性系的半同胞家系种子8份和当地种源种子1份,于1983年2月育苗。
2.2 试验方法
1984年3月造林。采用随机区组设计,参试材料69个,其中全同胞家系60个(4个自交组合种子不发芽)、半同胞家系8个、当地种源(对照)1个,5次重复,8株单行小区。2014年12月,每木调查31年生砍倒木树高和胸径,并按下列公式计算单积。
V=0.000 058 777 042D1.973 699 831H0.896 461 57
2.3 优良家系入选标准
优良杂交组合和其它材料入选标准主要以材积生长量为主[11]。本研究优良杂交组合入选标准平均树高、胸径、材积分别大于当地种源(对照)的12%、20%、40%以上,且与亲本相比有杂种优势。用于杂交的优良无性系,其入选标准要求该无性系的半同胞家系材积大于当地种源(对照)20%以上。作为建立双系种子园的亲本,要求其正、反交材积都大于当地种源(对照)30%以上,且自交种子不发芽或不能有衰退现象。
2.4 数据分析
按Griffing提出的双列杂交试验方法3的分析程序[8],先对小区平均数进行方差分析、显著性检验,然后对组合平均数进行配合力方差分析,按随机模型估算出各遗传方差的分量;再按固定模型,对亲本的配合力效应值作出估算[9]。
杂交组合的杂种优势计算公式[10]:
H=(F1-OP)/OP×100%
式中:H为杂种优势程度;
F1为任何1个组合的平均值;
OP为该组合中1个优势亲本自由授粉子代平均值。
自交衰退计算公式:
D=(FS1-OP)/OP×100%
式中:D为自交衰退程度;
FSl为任何1个亲本自交Fl代家系均值;
OP为相应亲本的自由授粉子代平均值。遗传方差分量GVC用该项方差分量占该性状遗传总方差的百分比来衡量。
3 结果与分析
3.1 生长量及差异性
60个杉木全同胞家系31年生时平均树高21.4m,平均胸径27.1cm,平均单株材积0.6114m3,折合产木材642m3/hm2。靖州县当地种源(对照)平均树高19.3m,平均胸径24.4 cm,平均单株材积0.4570m3,折合产木材480m3/hm2。60个全同胞家系平均树高、胸径、单株材积分别比靖州县当地种源(对照)高10.9%、11.1%、33.8%。
生长量最大的组合与最小的组合单株材积分别为0.7513m3和0.3345m3,前者是后者的2.25倍。生长量最大的亲本半同胞和最小的亲本半同胞单株材积分别为0.676 4 m3和0.510 2m3,前者是后者的1.33倍。总之,试验林整体生长水平高,内部分化大[12]。经方差分析,各主要生长指标组合间、重复间都存在极显著差异(见表1)。
3.2 杂种优势
根据表2、表3结果,杂交组合平均材积为0.6061m3,半同胞家系平均材积为0.5946m3,半同胞家系生长水平整体上要低于全同胞家系;将杂交组合材积与母本半同胞、父本半同胞及父母本半同胞材积中亲值作简相关分析,其相关系数分别为0.2089、0.1485、0.3896,杂交组合与父母本半同胞中亲值间存在着显著的正相关关系;交配组合超亲优势的变幅很大,其变化在-25.96%~32.51%之间;正反交效应十分明显,靖6与靖31杂交,其正反交杂种优势相差达31.72%。
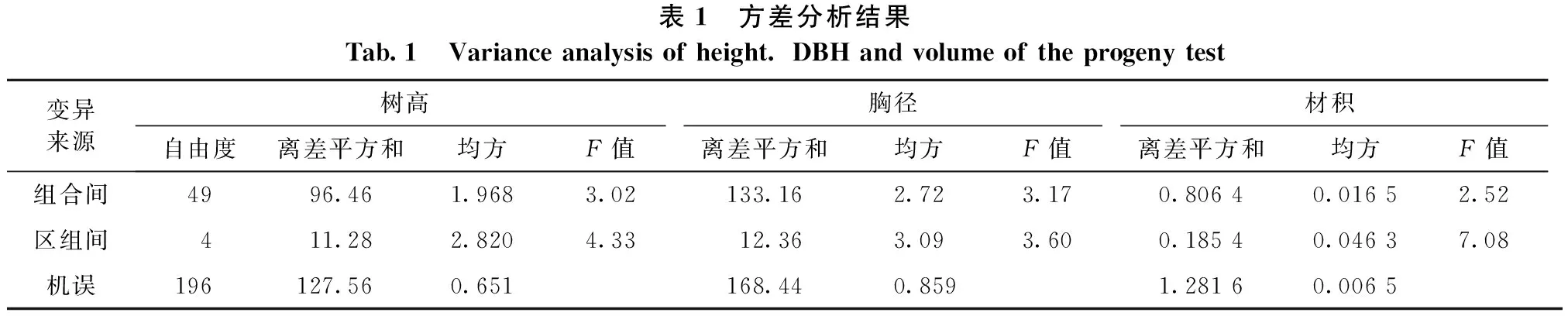
表1 方差分析结果Tab.1 Varianceanalysisofheight.DBHandvolumeoftheprogenytest变异来源树高胸径材积自由度离差平方和均方F值离差平方和均方F值离差平方和均方F值组合间4996.461.9683.02133.162.723.170.80640.01652.52区组间411.282.8204.3312.363.093.600.18540.04637.08机误196127.560.651168.440.8591.28160.0065

表2 杉木全双列杂交材积杂种优势分析Tab.2 HeterosisinvolumeofwoodofprogeniesoriginatingfromdiallelmatingofChinesefir亲本平均单株材积(m3)杂种优势(%)母本(P1)父本(P2)正交反交正反交差正交反交靖2 靖6 0.59920.6673-0.06802.1013.69靖2 靖7 0.67860.59220.086415.620.90靖2 靖130.56120.51050.0507-1.36-13.02靖2 靖160.65230.60790.04459.291.83靖2 靖230.71860.65880.059822.4312.25靖2 靖310.68290.53100.15202.48-20.33靖2 靖350.65180.62150.030211.055.89靖6 靖7 0.49230.4993-0.0072-15.83-14.62靖6 靖130.57020.5936-0.0237-2.511.51靖6 靖160.62120.61900.00214.063.71靖6 靖230.57240.55210.0203-4.10-7.50靖6 靖310.49350.7048-0.2114-25.965.76靖6 靖350.52650.6095-0.0831-10.004.22靖7 靖130.61630.46210.154311.71-16.25靖7 靖160.57110.6624-0.0911-4.3010.97靖7 靖230.61470.52740.08739.75-5.83靖7 靖310.56900.5852-0.0162-14.6312.03靖7 靖350.61070.48191.28978.97-14.04靖13靖160.60350.6170-0.01361.123.39靖13靖230.65660.63990.016917.2614.24靖13靖310.61160.60500.0066-8.23-9.22靖13靖350.69930.64100.058324.7714.35靖16靖230.55730.51730.0400-6.63-13.34靖16靖310.66820.62750.04980.26-7.21靖16靖350.58970.6865-0.0968-1.1915.03靖23靖310.66890.6791-0.01030.361.91靖23靖350.63430.7421-0.107813.2532.51靖31靖350.64330.56390.0792-3.48-15.38平均 0.6061
3.3 自交效应
8个自交系中,4个自交系的种子有发芽能力,4个自交系的种子无发芽能力;4个种子有发芽能力自交系的造林后3个有衰退效应,1个有优势效应(见表3)。有衰退现象自交系的材积最大衰退达29.47%;有优势效应自交系的材积长量超过了半同胞亲本,高于试验林平均水平。
3.4 配合力
3.4.1 配合力方差分析和遗传参数估算 经配合力方差分析,树高、胸径、材积的一般配合力、特殊配合力和正反交效应的差异都达到了显著或极显著水平(见表4),表明这些效应都存在着遗传变异。表4结果还说明:树高、胸径的不同配合力效应遗传方差比基本接近,都在30%~38%,3种配合力的重要性相当;材积的不同配合力遗传方差比以特殊配合力的最大,占44.40%,材积主要受特殊配合力支配。从遗传能力的相对大小比较,树高遗传力较大,胸径和材积遗传力相近且较小。

表3 自交衰退分析Tab.3 Growthcomparisionofopen-pollinatedprogeniesandselfings亲本自由授粉自交系自交衰退树高(m)胸径(cm)材积(m3)树高(m)胸径(cm)材积(m3)树高(%)胸径(%)材积(%)靖2 21.026.90.597618.823.70.4215-10.48-11.90-29.47靖6 20.427.10.590921.227.30.62053.920.745.01靖1320.725.60.535019.825.10.4945-4.35-1.95-7.57靖1621.627.80.654020.926.60.5820-3.24-4.32-11.01靖7 19.825.50.5102自交系种子不发芽靖2321.027.00.6020自交系种子不发芽靖3121.928.10.6764自交系种子不发芽靖3520.926.80.5907自交系种子不发芽平均20.926.90.5946
3.4.2 一般配合力效应 计算8个亲本树高、胸径、材积的一般配合力效应,并进行差异性比较,结果列表5。

表4 配合力方差分析及遗传参数估算Tab.4 Varianceanalysisofcombiningabilitiesandestimationofgeneticparameters变因树高胸径材积均方F值方差比(%)均方F值方差比(%)均方F值方差比(%)GCA2.315.7830.382.442.6837.550.02383.0127.44SCA1.032.5836.121.331.4631.870.01351.7144.06REC0.822.0533.501.421.5630.580.01481.8728.50机误0.400.910.0079遗传力Vg=58.35 Va=22.15Vg=39.78 Va=17.54Vg=41.88 Va=16.63 I注:GCA、SCA、REC分别为一般、特殊、正反交配合力;Vg、Va分别为一般、个体遗传力。

表5 一般配合力效应值及差异性比较Tab.5 Evaluationofgeneralcombiningabilitiesandsignificancecomparisons名次树高(m)胸径(cm)材积(m3)亲本Gi亲本Gi亲本Gi1靖310.562**靖2 0.800*靖230.02332靖230.538*靖350.708*靖2 0.02273靖350.323靖230.527靖350.01954靖130.026靖160.521靖310.01495靖2 0.017靖310.109靖160.00886靖16-0.125靖13-0.350靖13-0.00937靖6 -0.660**靖6 -0.820靖6 -0.0343**8靖7 -0.676**靖7 -1.495**靖7 -0.0455**
表5结果表明,树高、胸径、材积的一般配合力效应值排序有差异,即按不同性状选择一般配合力高的无性系,其筛选结果不尽相同。以3个性状总体衡量,亲本靖2、靖23和靖35有显著的正向效应,是3个优良的杂交亲本;靖6和靖7有显著的负向效应,是2个较差的杂交亲本。不同亲本之间比较,其效应值的差异是显著的,材积一般配合力效应值最大相差300%。
3.4.3 特殊配合力效应 特殊配合力反映的是一个交配群体中特定的交配组合与子代总平均值的离差。以表2数据计算材积的特殊配合力效应值(见表6),并进行差异显著性检验。

表6 杂交组合特殊配合力效应值Tab.6 Thespecialcombiningabilityeffectvalueofhybrid-ization母本(P1)父本(P2)正交反交平均靖2 靖6 -0.1080.9540.423靖2 靖7 1.130-0.2170.457靖2 靖13-0.700-1.490-1.095-*靖2 靖160.7200.0280.374靖2 靖231.7530.8211.287**靖2 靖311.197-1.1710.013靖2 靖350.7120.2400.476靖6 靖7 -1.774-1.665-1.719-**靖6 靖13-0.560-0.195-0.377靖6 靖160.2350.2010.218靖6 靖23-0.525-0.842-0.683靖6 靖31-1.7551.538-0.108靖6 靖35-1.2410.053-0.594靖7 靖130.159-2.244-1.043-*靖7 靖16-0.5460.8770.166靖7 靖230.134-1.227-0.546靖7 靖31-0.578-0.326-0.452靖7 靖350.072-1.936-0.932-*靖13靖16-0.0410.1700.065靖13靖230.7870.5270.657靖13靖310.086-0.0170.034靖13靖351.4530.5440.998*靖16靖23-0.761-1.384-1.072-*靖16靖310.9680.3340.651靖16靖35-0.2561.2530.499靖23靖310.9791.1381.058*靖23靖350.4402.1201.280**靖31靖350.580-0.658-0.039
表6结果表明: 特殊配合力效应值与0比较,组合靖2与靖23、靖23与靖35、靖23与靖31、靖13与靖15的材积特殊配合力显著大于0,为正向显著类型;组合靖6与靖7、靖2与靖13、靖16与靖23、靖7与靖13、靖7与靖35的材积特殊配合力显著小于0,为显著负向类型;其他组合均为不显著类型。不同组合相互比较,材积性状特殊配合力效应值的差异亦是显著的。
3.4.4 正反交效应 3个性状各组合的正反交效应值及差异显著性检验结果表明:亲本正反交效应也同样具有正负向的差异,以材积为例,正向显著类型占7.14%,负向显著类型占3.57%,不显著类型占89.3%。正交靖7×靖13、靖2×靖31,反交靖31×靖6具有显著的正向显著性。各组合间相互比较,其差异也是显著的。
3.5 亲本及杂交组合的综合评价
靖2、靖23和靖35有较高的一般配合力效应值,用它们组配,能产生较多生长量突出的组合,是3个较好的育种亲本。如靖35的正、反交效应方差大,可望产生生长量突出的正交或反交组合,正交靖7×靖35、靖31×靖35和反交靖35×靖23、靖35×靖16生长突出;靖23的特殊配合力方差大,可望产生生长突出的正、反交组合,靖23与靖2、靖13、靖3、靖35组合的正、反交生长均突出。
靖31、靖16、靖23、靖2、靖6、靖35等半同胞家系速生丰产,是6个优良的1.5代种子园建园材料。本试验中靖31、靖16、靖23、靖2、靖6、靖35的31年生平均单株材积生长量分别达0.6764m3、0.6540m3、0.6020m3、0.5976 m3、0.5909m3、0.5907m3,分别比当地种源(对照)大48.0%、43.1%、31.7%、30.8%、29.3%、29.2%[13]。
靖23与靖35组合、靖23与靖31组合,其正交和反交材积都大于靖州种源(对照)30%以上,且自交种子不发芽或自交有优势,是2组建立双系种子园的理想材料[14]。
19个杂交组合,包括靖35×靖23、靖2×靖23、靖31×靖6、靖13×靖35、靖35×靖16、靖2×靖31、靖31×靖23、靖2×靖7、靖23×靖31、靖16×靖31、靖6×靖2、靖16×靖7、靖23×靖2、靖13×靖23、靖2×靖16、靖2×靖35、靖31×靖35、靖35×靖13、靖23×靖13,相对于对照材积增益分别达62.4%、57.2%、54.2%、53.0%、50.2%、49.4%、48.6%、48.5%、46.4%、46.2%、46.0%、44.9%、44.2%、43.7%、42.7%、42.6%、40.8%、40.3%、40.0%。可从这些优良组合中选择优良单株作为营建第2代种子园的材料[15]。
4 结论与讨论
(1) 全双列交配设计是林业先进国家进行子代测定的设计方法之一。选择较多亲本进行全双列杂交,工作量虽较大,但有利于更好地研究自交效应和正、反交效应,挖掘更多的优良组合。
(2) 杂交组合的材积与亲本半同胞的中亲值间达到了显著的正相关,杂交后代的表现可根据其父母本半同胞家系生长水平作出大概估计。
(3) 本试验中,8个杉木自交系中有4个自交系种子无发芽能力,占50%的;4个种子有发芽能力的自交系中,有3个自交系林分生长明显衰退。种子园小区中保证一定数量的无性系可减少自交种子不发芽或自交系生长衰退的机率。对于个别自交优势系(如靖6),可建立单系种子园充分利用自交优势。
(4) 各全同胞家系间存在真实的遗传变异,且3种配合力效应都达到显著或极显著差异水平,说明杉木育种在注意一般配合力的同时,不可忽视特殊配合力和正、反交效应。
(5) 树高、胸径的3种配合力相对重要性接近,遗传方差比都在30%~38%;材积主要受特殊配合力支配,遗传方差比为44%。世代传递能力,以树高遗传力较大,胸径、材积遗传力相近且较小。
(6) 评选出优良杂交亲本3个,1.5代种子园优良亲本6个,双系种子园优良亲本2组,同时评选出19个优良组合,从中可选择优株建立第2代杉木种子园。
[1] 丘进清.杉木种子园技术综述[J].南京林业大学学报,2006,22(5):103-106.
[2] 郑仁华,施季森.福建省杉木良种繁育现状与对策[J].林业科技开发,2004,18(2):3-7.
[3] 孙鸿有,郑勇平,翁春媚.杉木种子园种子品质性状变异及遗传参数[J].浙江林学院学报,2005,22(1):61-65.
[4] 阮梓材,胡德活,陈仲.杉木种子园建园亲本的综合选择[J].广东林业科技,1994.21(2):14-19.
[5] 吴琼美,樊汝汶,肖石海.杉木传粉生物学的观察[J].南京林业大学学报,1995,19(1):10-18.
[6] 官康林,黄坚钦,何福基.杉木种子园球果的出籽率和饱籽率[J].浙江林学院学报,1997,14(2):127-133.
[7] 施季森.杉木交配设计后代遗传分析,种子园技术[M].北京:北京科技出版社,1992.
[8] 陈岳武.配合力分析在杉木数量遗传研究中的应用,陈岳武论文选集[D].南京:南京林业大学,1989.
[9] 沈熙环.林木育种学[M].北京:中国林业出版社,1990.
[10] 沈熙环.创造条件向高世代育种发展.种子园优 质高产技术[M]..北京:中国林业出版社,1994.
[11] 陈岳武.杉木优树选择方法的研究.陈岳武论文选集[D].南京:南京林业大学:1989.
[12] 余荣卓.杉木种子园子代长期测定结果与早晚期选择相关性分析[J].福建林学院学报,1998(4):1-4.
[13] 孙鹏.杉木家系选择及效益分析[J].四川林业科技,1999[J].26(2):11-14.
[14] 徐清乾,许忠坤.杉木杂交组配与两系种子园建立技术研究[J].湖南林业科技,2004.31(6):18-20.
[15] 徐清乾,许忠坤,程政红.杉木第二代种子园建立技术研究.湖南林业科技,2002,29(4):16-19.
AnalysisofChinesefirfulldiallelprogenygeneticvariation
XU Qingqian1, WU Chengtian2, XU Zhongkun1*
(1.Hunan Academy of forestry, Changsha 410004, China;2.Jingzhou Paiyashan State-Owned Forest Farm, Jingzhou 418400, China)
The eight clones of Chinese fir complete diallel crosses of 32-year-old forest's tree height, DBH and volume for test the advantages of hybrid and inbred fading effect analysis, and the combining ability variance and combining ability effect value were calculated. The result show that: true genetic variation exists in three traits, general combining ability, special combining ability, positive and negative cross effect variance all reached significant level, and volume to special combining ability of the genetic variance ratio (GVC) is maximum. Selected three excellent clones for hybridization and two growps of excellent clones for Bi-clonal seed orchard, and 19 combination of cross superior trees for second generation.
Chinese fir; clone; full line crossing; combining ability; genetic variance
2015-08-25
农村领域国家科技计划项目“湖南杉木三代育种技术研究与示范”(2012BAD01B020102)。
徐清乾(1963-),男,湖南省浏阳市人,研究员,主要从事林木育种研究。
* 为通讯作者。
S 791.27.01
A
1003 — 5710(2015)06 — 0007 — 06
10. 3969/j. issn. 1003 — 5710. 2015. 06. 002
(文字编校:唐效蓉)
