白云山自然保护区蕨类植物生境因子与多样性的关系分析
2015-11-17宿秀江向魁文颜绍馗
宿秀江, 万 鑫, 向魁文, 颜绍馗
(1.湖南白云山国家级自然保护区, 湖南 保靖 416500; 2.中国科学院沈阳应用生态研究所, 辽宁 沈阳 110016;3.湖南省保靖县林业局, 湖南 保靖 416500; 4.湖南省湘西州林业局, 湖南 吉首 416000)
白云山自然保护区蕨类植物生境因子与多样性的关系分析
宿秀江1,2, 万 鑫3, 向魁文4, 颜绍馗2
(1.湖南白云山国家级自然保护区, 湖南 保靖 416500; 2.中国科学院沈阳应用生态研究所, 辽宁 沈阳 110016;3.湖南省保靖县林业局, 湖南 保靖 416500; 4.湖南省湘西州林业局, 湖南 吉首 416000)
在白云山自然保护区,调查分析了37种森林类型中影响蕨类植物分布的生境因子以及生境因子与蕨类植物多样性之间的关系。结果表明:(1)在所调查的251块样地内共发现有蕨类植物8 609株,分属16科34属90种;(2)生境因子包括森林类型、海拔、坡度、坡向、土壤腐殖质厚度以及凋落物层厚度对物种多样性和群落组成有显著影响;(3)在所调查的生境因子中,森林类型和海拔是最重要的影响因子,其中海拔对多样性的解释量高达11%。此结果表明森林类型和海拔是对蕨类分布起主要作用的环境因子,因此在自然保护区制定多样性保护策略时,应优先考虑植被类型和海拔的影响。
自然保护区; 生物多样性保护; 生境; 海拔; 林下植被
白云山自然保护区地处武陵山脉南坡中段,位于湖南省保靖县境内,始建于1998年。因生物多样性的特殊地位,2008年被湖南省批准为省级自然保护区,2013年晋升为国家级自然保护区[1]。保护区境内地貌结构独特,地势崎岖陡峭,沟壑纵横,森林密布,是蕨类植物的理想居地。因种类繁杂,目前已成为我国蕨类植物多样性研究的重要基地。然而由于本区生物多样性的丰富程度与重要性发现相对较晚,故调查和研究积累非常贫乏。直到2006年,在国家科技部自然保护区平台项目的支持下才初步整理出白云山植物名录,报道蕨类植物154种。2009—2012年保护区又进一步组织中国林业科学研究院、吉首大学等多家单位的学者完成了保护区综合考察,整理出白云山蕨类植物有45科106属396种,约占我国蕨类植物种数的18.6%[1-2]。在此基础上,2013年,龙凤菊[3]根据这些资料对白云山蕨类植物区系进行了重新整理和论述,认为其可归属于“耳蕨-鳞毛蕨植物区系”。
由于本区以往的研究工作主要重视名录的获取以及区系的归属划分,而忽视了蕨类植物的数量调查以及生境与蕨类多样性的关系研究,因此对于保护区多样性保护策略的制定,其参考价值相对较弱。近年来随着对蕨类资源利用的开发,多样性破坏严重,许多蕨类例如蛇足石杉(Huperziaserrata)、观音座莲(Angiopterisesculenta)、扇蕨(Neocheiropterispalmatopedata)以及中华水韭(Isoetessinensis)等持续出现衰减、濒危,甚至极度濒危的现象[4-6]。蕨类植物多样性保护的重要性越来越迫切,特别是在蕨类特别丰富的自然保护区,迫切需要制定蕨类保护的策略。为此我们选择了本区37种主要的森林植被类型,设置了251块样地,针对蕨类分布的种类和数量以及影响蕨类植物多样性的多个生境因子进行了调查和比较,确定影响本区蕨类多样性的主导环境因子,以期为本区蕨类植物多样性保护方案的制定提供初步的参考依据。
1 研究区概况
湖南白云山自然保护区地处中国十六个生物多样性关键地区之一——武陵山区,位于湘西北保靖县境内,华中、西南地区交汇处,东经109°16′35″—109°32′52″,北纬28°37′42″—28°50′58″,总面积20 158.6 hm2,属中亚热带山地季风湿润气候。区内年平均气温为12.4 ℃,最冷月1月,平均气温1.5 ℃,最热月7月平均气温23 ℃,≥10 ℃的天数180~220天,年有效积温3 000~4 300 ℃,年无霜期平均239天。年均降雨量1 600 mm,7—9月最多达520~560 mm,降雨日210多天,年平均相对湿度84%以上,全年有雾日平均145天,年辐射量较低。根据出版的白云山保护区科考报告,本区植被类型可划分为3个植被型组,10个植被型,12个植被亚型,59个植物群系组,83个植物群系[1],主要植被为马尾松、杉木人工林以及亚热带地带性常绿阔叶林。保护区内土壤依海拔自下到上可划分为黄红壤、黄壤、黄棕壤三大类。成土母岩主要有砂页岩、砂岩、石灰岩。
2 研究方法
2.1 野外调查
调查位置选择白云寺、吊颈湾、凌云亭、天台山等共40个自然村落,经度范围109°18′29″—109°28′15″E,纬度范围28°39′29″—28°49′46″N,海拔范围300~1 200 m,调查时间从2010年8月至2012年10月。调查时选择有代表性的森林类型,包括人工林、撂荒地、针阔混交林、常绿阔叶林,其中撂荒地为茶园与人工林在1980年破坏和采伐后自然演替形成的荒地,群落外貌为灌丛,乔木以幼树为主,属于常绿阔叶林的初始发育阶段。本次共调查251块样地,样地面积为5 m×5 m,样地选择的依据是包含蕨类植物并且属于同一群丛内的连续区域选择作为一个样地,为了尽量避免空间自相关,降低数据分析的难度,各样地之间的距离均超过500 m。调查的251块样地分属于37种植被类型,分别为:杉木人工林、杉木马尾松混交林、杉木柳杉混交林、楠竹人工林、杉木酸枣混交林、盐肤木灌丛、马尾松次生林、栲树马尾松混交林、甜槠灌丛、光叶柯灌丛、杉木鹅耳枥混交林、枫香马尾松混交林、油茶次生林、马尾松润楠混交林、杉木枫香混交林、鹅掌楸杉木灌丛、盐肤木次生林、甜槠次生林、白背绣球灌丛、润楠次生林、鹅耳枥灌丛、银木荷灌丛、冬青灌丛、猴樟次生林、枫香次生林、宜昌润楠次生林、栲树次生林、青冈次生林、柳杉人工林、枫香润楠次生林、银木荷次生林、光叶柯金缕梅灌丛、尖叶四照花灌丛、灯台树青榨槭次生林、海桐次生林、暖木次生林、凹叶厚朴灌丛。调查时记录蕨类的种类、株数、盖度并用游标卡尺测量个体的基径,并同时测定生境因子,包括海拔、坡度、凋落层厚度、腐殖质层厚度以及坡向,这些参数具体的分布范围参看表1。
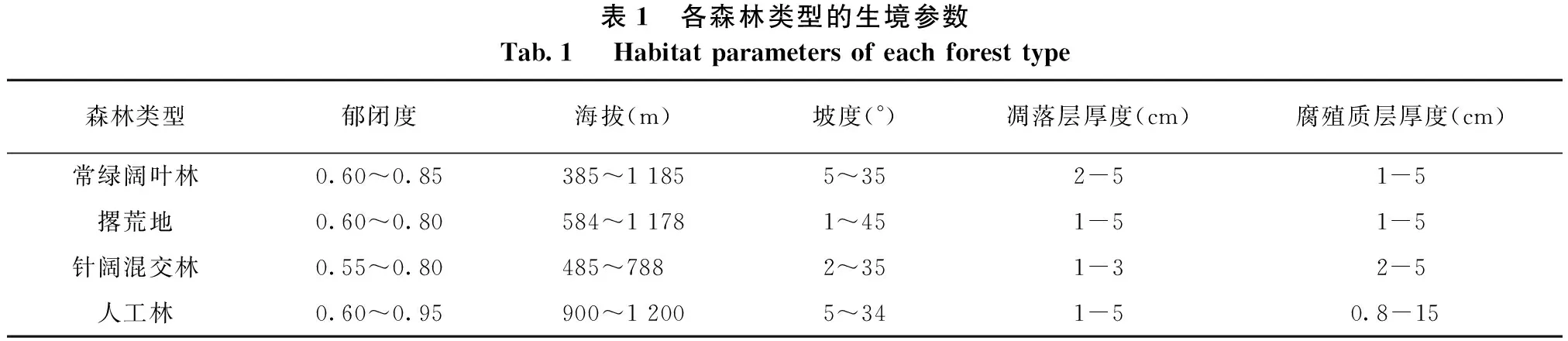
表1 各森林类型的生境参数Tab.1 Habitatparametersofeachforesttype森林类型郁闭度海拔(m)坡度(°)凋落层厚度(cm)腐殖质层厚度(cm)常绿阔叶林0.60~0.85385~11855~352-51-5撂荒地0.60~0.80584~11781~451-51-5针阔混交林0.55~0.80485~788 2~351-32-5人工林0.60~0.95900~12005~341-50.8-15
2.2 数据处理
数据处理时,群落参数包括多度、多样性和群落组成以样地(25 m2)为基本单元进行统计,其中多度使用个体计数表述,即每样地多少株丛数,多样性使用Shannon指数、Simpson指数和物种丰富度指数评估,群落组成使用NMS(多维标度排序)排序方法获取。检查生境因子的效应时,离散变量包括森林类型使用方差分析,当正态假设不能通过时改用非参数方差分析,相应多重比较使用Dunn检验,连续型变量则使用回归分析,数据分析应用R软件实现。
3 结果与分析
3.1 蕨类的基本情况
根据调查结果,251块样地共发现有林下蕨类8 609株,分属16科34属90种,其中鳞毛蕨科(Dryopteridaceae)所辖物种数最多,达32种,蹄盖蕨科(Athyriaceae)次之,18种,各科所含物种数和属数情况参看表2。此外,中日金星蕨(Parathelypterisnipponica)和假黑鳞耳蕨(Polystichumpseudomakinoi)为本地区数量上的优势种,分别占调查所获个体总数的19.52%、13.61%。黑足鳞毛蕨(Dryopterisfuscipes)和狗脊(Woodwardiajaponica)为本地区分布范围最广的种,分别占调查样地面积的22.71%和21.11%。
3.2 生境因子与多样性指数的关系分析
根据各样地间蕨类多度和多样性的统计,样地的多度从1到1 371株不等,平均34.3株,变异系数高达2.8,而多样性,其中Shannon指数0.00~2.27,平均0.97,Simpson指数0.00~0.88,平均0.50,物种丰富度指数1~12,平均4.13,三类多样性指数变异系数分别为0.58、0.51、0.60,均超过0.5。为了解释这种变异,选择森林类型、海拔、坡向、坡度、凋落层厚度以及土壤腐殖质层厚度作为生境因子,以检查蕨类多样性与生境的关系。考虑森林类型为离散变量,因此使用方差分析,结果发现森林类型对蕨类多度和多样性指数均有极显著影响(见表3)。Dunn检验进一步表明:多度(株数)和物种丰富度指数的比较,人工林、针阔混交林和常绿阔叶林均显著高于撂荒地,株数均超过50%,种数均超过25%,但人工林、针阔混交林和常绿阔叶林之间无明显差异;而Shannon多样性指数和Simpson多样性指数的比较,以撂荒地最低,针阔混交林最高,人工林和常绿阔叶林居中,该结果相对于多度和丰富度指数林型之间比较在差异上更为突出,两两比较显著性差异结果也较为复杂。另外,由于海拔、坡向、坡度、凋落层厚度、土壤腐殖质层厚度等生境因子为连续变量,因此做回归分析,结果如图1,其中多度不受这些指标中任意单一变量的影响,测定系数均趋向于0,而在多样性的分析里面,除凋落物层厚度以外,海拔、坡度、坡向以及土壤腐殖质厚度则对多样性均有一定程度的影响,其中海拔对Shannon指数和Simpson指数的解释量达11%,远高于其他生境因子。因此,这些生境因子对生物多样性的影响为海拔最强,坡度、坡向以及土壤腐殖质厚度次之,而凋落物层厚度最弱,参看图1。
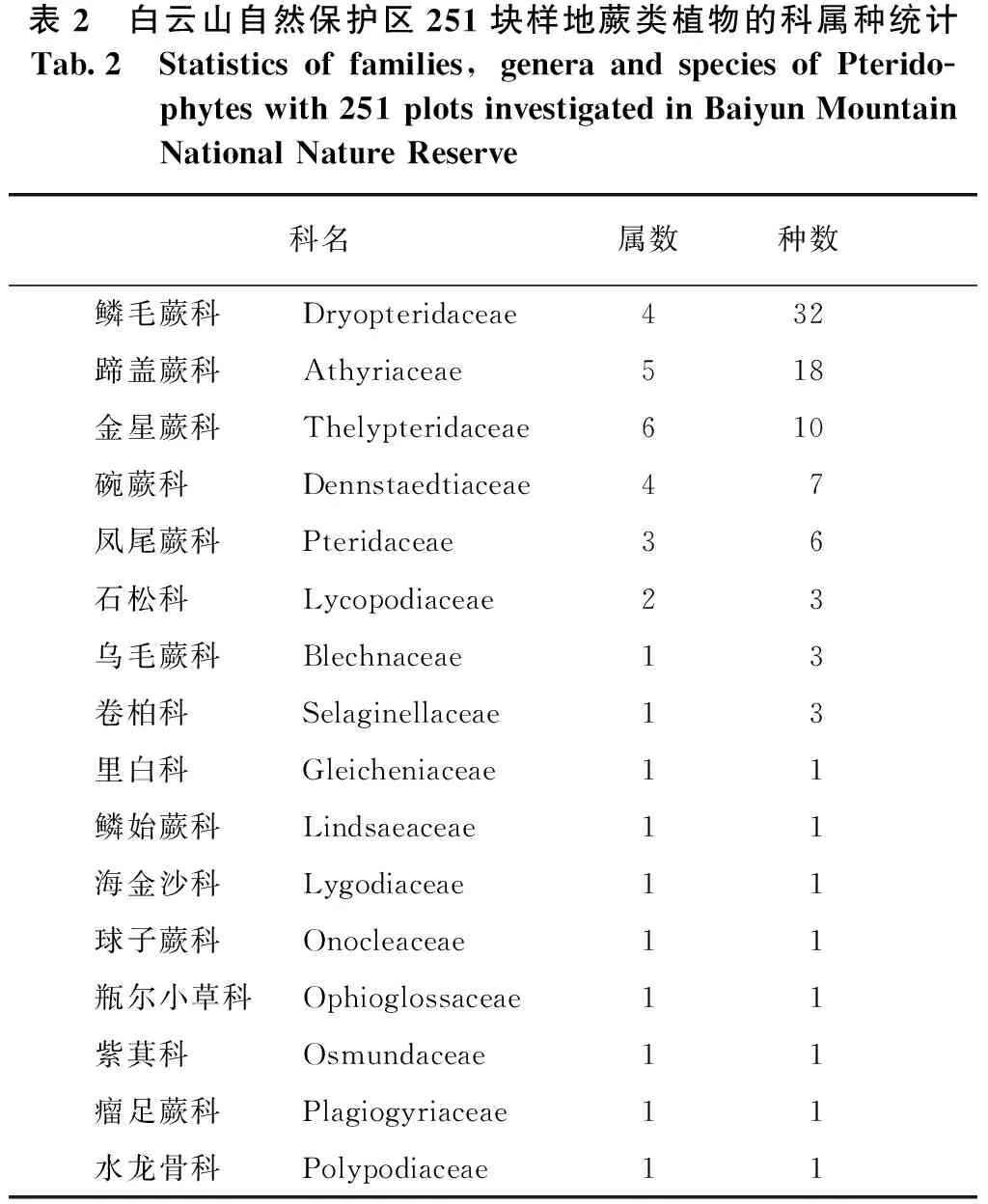
表2 白云山自然保护区251块样地蕨类植物的科属种统计Tab.2 Statisticsoffamilies,generaandspeciesofPterido-phyteswith251plotsinvestigatedinBaiyunMountainNationalNatureReserve科名属数种数鳞毛蕨科Dryopteridaceae432蹄盖蕨科Athyriaceae518金星蕨科Thelypteridaceae610碗蕨科Dennstaedtiaceae47凤尾蕨科Pteridaceae36石松科Lycopodiaceae23乌毛蕨科Blechnaceae13卷柏科Selaginellaceae13里白科Gleicheniaceae11鳞始蕨科Lindsaeaceae11海金沙科Lygodiaceae11球子蕨科Onocleaceae11瓶尔小草科Ophioglossaceae11紫萁科Osmundaceae11瘤足蕨科Plagiogyriaceae11水龙骨科Polypodiaceae11
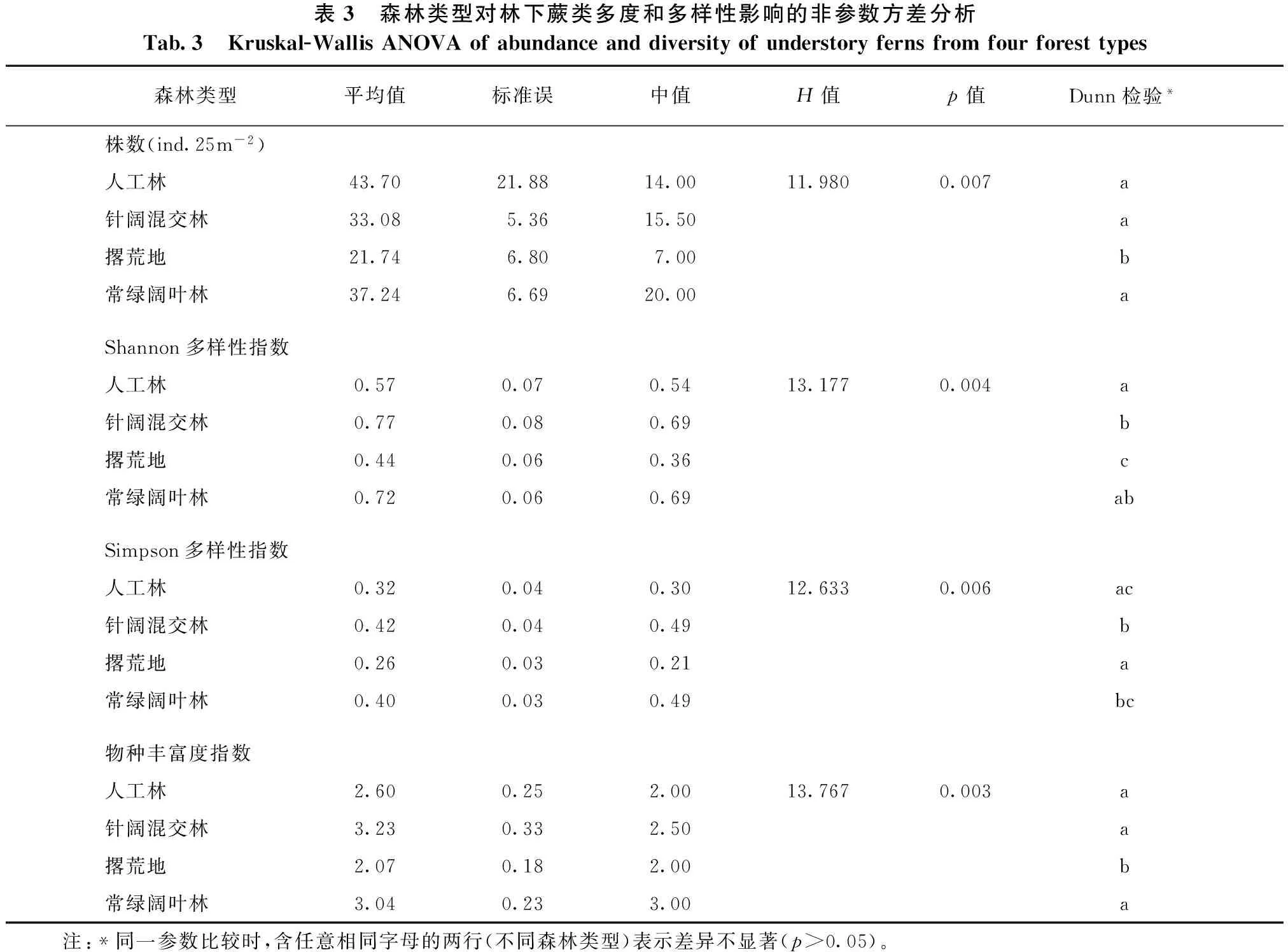
表3 森林类型对林下蕨类多度和多样性影响的非参数方差分析Tab.3 Kruskal-WallisANOVAofabundanceanddiversityofunderstoryfernsfromfourforesttypes森林类型平均值标准误中值H值p值Dunn检验*株数(ind.25m-2)人工林43.7021.8814.0011.9800.007a针阔混交林33.085.3615.50a撂荒地21.746.807.00b常绿阔叶林37.246.6920.00aShannon多样性指数人工林0.570.070.5413.1770.004a针阔混交林0.770.080.69b撂荒地0.440.060.36c常绿阔叶林0.720.060.69abSimpson多样性指数人工林0.320.040.3012.6330.006ac针阔混交林0.420.040.49b撂荒地0.260.030.21a常绿阔叶林0.400.030.49bc物种丰富度指数人工林2.600.252.0013.7670.003a针阔混交林3.230.332.50a撂荒地2.070.182.00b常绿阔叶林3.040.233.00a 注:*同一参数比较时,含任意相同字母的两行(不同森林类型)表示差异不显著(p>0.05)。

图1 生境因子与蕨类多度和多样性的关系Fig.1 Relationship between habitat parameter and ferns abundance and diversity

续图1 生境因子与蕨类多度和多样性的关系Continued Fig.1 Relationship between habitat parameter and ferns abundance and diversity
3.3 生境因子与蕨类群落组成的关系分析
根据群落组成矩阵,NMS排序能提取两显著性的排序轴映射群落组成(Stress=0.423,R2=0.155,P=0.032)。其中第一排序轴(NMS1)与森林类型无关(Kruskal-Wallis ANOVA,H=3.582,P=0.310),而第二排序轴与森林类型显著关联(Kruskal-Wallis ANOVA,H=10.323,P=0.016)。两轴与各生境因子的相关分析参看表4,其中第一轴与海拔、凋落物层厚度、腐殖质层厚度显著相关,以海拔的相关系数最大,而第二轴仅与坡度显著相关。由于排序轴反映群落组成,因此根据这种关联性,可以确定森林类型、海拔、坡度、坡向、凋落物层厚度、腐殖质层厚度对群落组成有显著影响,其中森林类型、海拔和坡度应是对群落组成变化起主要作用的影响因子。

表4 生境因子与非度量多维标度排序轴的相关性分析Tab.4 Correlationanalysisbetweenhabitatfactorsandnon-metricmultidimensionalscaling(NMS)ordinationaxes生境因子NMS1NMS2rPrP海拔0.1670.000-0.0430.258坡度-0.0350.368-0.2220.000凋落物层厚度0.1060.0050.0370.329腐殖质层厚度0.0870.0240.0180.643坡向0.0630.097-0.0320.406
4 结论与讨论
物种多样性依赖于物种的分布,而物种分布与生境因素紧密关联[7-8],本文对于蕨类生境和多样性关系的研究,主要反映以下三方面的结果:
(1)生境对蕨类的多样性有显著影响。以多样性指数为检测指标,生境因子的梯度变化引起物种分布的多样性出现差异,在所调查的生境因子中除凋落物层厚度以外,森林类型、海拔、坡向、坡度、凋落层厚度、土壤腐殖质层厚度均对多样性指数有显著影响(见图1和表3);以群落组成为检测指标,除坡向外,其余指标均对群落组成有显著影响(见表4)。一般认为物种的分布受生境、种源以及两者的联合影响[9]。在我们的研究里面,笔者认为生境条件是主要原因,而种源的作用可以忽视,主要理由有如下几点:第一,种源的扩散本身依赖于生境条件,需要借助风等载体,在白云山地区蕨类植物不但种类多,而且数量多,种源传播受限制的可能性较少,同样也有研究认为蕨类的孢子小且生产的数量巨大,能有效地减少物种传播的限制[10];第二,蕨类植物的生活史依赖于生境条件,与种子植物的受精过程发生在种子传播开始之前不同,蕨类植物的受精过程发生在孢子传播之后,两类孢子必须同时落在相邻的适宜生境才能完成受精[11],且离不开水;第三,蕨类植物分布与环境因子的关联性,其可靠程度要高于种子植物,在自然的生态系统里面,种子植物是许多动物的食物来源,动物活动能有效干扰种子的散布与授粉的过程,因此在确定物种分布时得到的环境效应实际上是环境因子与动物的联合效应,相反,蕨类植物因适口性差,对于食物网的作用轻微[12],因此动物传播对蕨类的分布影响较少,一些研究也认为蕨类植物对非生物因子的依赖要远高于种子植物[13],例如受精过程的完成需要水分的参与[11],在分析物种分布与环境因子的关系时,有些学者甚至提议使用蕨类植物作为研究材料[14]以及生长环境的评估指标[15]。
(2)各生境因子对多样性影响的相对重要性各不相同,森林类型和海拔是最主要的影响因子。生境条件包括森林类型和各种生境参数,在我们的研究里面发现森林类型是一个重要的影响因子,对多样性和群落组成均有影响。由于森林类型的形成与历史植被、演替阶段、干扰格局、附载植物的种类等有关,涉及多种人为因子和自然因子,因此是一个综合性的因素,在我们的研究里面发现人工林和阔叶林多样性差别不很明显,因此土地利用等引起的干扰格局可能起较少的作用,多样性主要的差别表现在撂荒地与其他林型的比较上,特别是撂荒地与常绿阔叶林比较非常明显,考虑到撂荒地以幼树为主,外貌呈灌丛状态,林冠层尚未真正形成,属于特殊形态的尚未发育完整的森林类型。相反,调查的人工林、针阔混交林、常绿阔叶林则林冠清晰可辨,处于完全的成熟状态,因此在本区森林类型对多样性的影响可能主要源于森林的发育阶段。在多种生境参数包括海拔、坡度、坡向、土壤腐殖质厚度、凋落物层厚度的比较中,无论是对多样性指数还是群落组成,海拔均是相对最主要的环境因子(见图1),随样地海拔的升高蕨类植物多样性出现缓慢的下降,海拔对Shannon多样性指数的解释量高达11%,而其他均低于4%。表4也显示海拔与第一排序轴的相关系数最高。张昌顺等[16]在澜沧江源区进行植物群落调查时发现影响高寒草甸植物种类组成和结构的环境因子从大至小依次为坡度、坡位、海拔和坡向,与本文研究结果海拔最大,坡度等次之稍有差别,这可能与地域以及区系的组成有关,但也同样表明了海拔和坡度确实是有重要影响的环境因子。Kluge等[14]在哥斯达黎加蕨类植物的研究中则发现蕨类的物种数随海拔的升高开始缓慢上升,然后缓慢下降,呈现一种单峰模式。在我们的研究中,蕨类植物在海拔600~800 m之间也有一个小小的峰值(见图1),但出现峰值的样地数较少,因此海拔与多样性的关系总体上仍呈现一种近似的线性关系。
(3)蕨类多样性与生境因素的关系可以认为能基本反映白云山地区生境与多样性的关系。首先,白云山是以森林类型为主的自然保护区,森林的林下层为本区蕨类分布的主要生境;其次,根据白云山251块样地的调查数据,总调查面积为0.62 hm2,约占保护区总面积的十万分之三,共记录16科34属90种蕨类,占整个自然保护区蕨类科数的35.56%,属数的32.07%,种数的22.72%,相对于十万分之三的取样面积来说,所选择的37种主要植被类型应为蕨类植物相对分布较集中的区域,因此选取样地的调查数据可基本反映大多数蕨类分布生境和多样性的关系。
综上所述,在白云山地区,蕨类的分布主要由生境条件决定,而森林类型和海拔是对蕨类分布起主要作用的环境因子,因此在自然保护区制定多样性保护策略时,应优先重视植被类型和海拔的影响。
[1] 宿秀江,刘芳,龙凤菊.湖南白云山国家级自然保护区综合科学考察报告 [M].杭州:浙江摄影出版社,2014.
[2] Flora of China Editorial Committee.Flora of China(Lycopodiaceae through Polypodiaceae)[M].Wu CY,Raven PH,Hong DY(eds.)Fl.China.Science Press & Missouri Botanical Garden Press,Beijing & St.Louis,2013,2-3:9-849.
[3] 龙凤菊.保靖县白云山蕨类植物区系初步研究 [J].湖南林业科技,2013,40(1):53-56.
[4] Kingston N,Waldren S,Smyth N.Conservation genetics and ecology of Angiopteris chauliodonta Copel.(Marattiaceae),a critically endangered fern from Pitcairn Island,South Central Pacific Ocean [J].Biological Conservation,2004,117(3):309-319.
[5] 邓亚妮,成晓,焦瑜,等.中国特有濒危植物扇蕨的生物生态学特性及其濒危机制初探 [J].生物多样性,2009,17(1):62-68.
[6] Chen YY,Yang W,Li W,et al.High allozyme diversity and unidirectional linear migration patterns within a population of tetraploid Isoetes sinensis,a rare and endangered pteridophyte [J].Aquatic Botany,2009,90(1):52-58.
[7] Bunce RGH,Bogers MMB,Evans D.,et al.The significance of habitats as indicators of biodiversity and their links to species [J].Ecological Indicators,2013,33:19-25.
[8] Warren SD,Alt M,Olson KD,et al.The relationship between the spectral diversity of satellite imagery,habitat heterogeneity,and plant species richness [J].Ecological Informatics,2014,24:160-168.
[9] Münzbergová Z,Herben T.Seed,dispersal,microsite,habitat and recruitment limitation:identification of terms and concepts in studies of limitations [J].Oecologia,2005,145:1-8.
[10] Guo QF,Kato M,Ricklefs RE.Life history,diversity and distribution:a study of Japanese Pteridophytes [J].Ecography,2003,26:129-138.
[11] Tájek P,Bucharová A,Münzbergová Z.Limitation of distribution of two rare ferns in fragmented landscape [J].Acta Oecologica,2011,37:495-502.
[12] Barrington DS.Ecological and historical factors in fern biogeography [J].Journal of Biogeography,1993,20:275-280.
[13] Ferrer-Castán D,Vetaas OR.Pteridophyte richness,climate and topography in the Iberian Peninsula:Comparing spatial and nonspatial models of richness patterns [J].Global Ecology and Biogeography,2005,14:155-165.
[14] Kluge J,Bach K,Kessler M.Elevational distribution and zonation of tropical pteridophyte assemblages in Costa Rica [J].Basic and Applied Ecology,2007,9:35-43.
[15] Larrea ML,Wernerb FA.Response of vascular epiphyte diversity to different land-use intensities in a neotropical montane wet forest[J].Forest Ecology and Management,2010,260:1950-1955.
[16] 张昌顺,谢高地,包维楷,等.地形对澜沧江源区高寒草甸植物丰富度及其分布格局的影响[J].生态学杂志,2012,31(11):2767-2774.
RelationshipbetweenPteridophytesdiversityandhabitatfactorsinBaiyunMountainNationalNatureReserve
SU Xiujiang1,2, WAN Xin3, XIANG Kuiwen4, YAN Shaokui2
(1.Baiyun Mountain National Nature Reserve, Baojing 416500, China;2.Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016, China;3.Forestry Bureau of Baojing County, Baojing 416500, China;4.Forestry Bureau of Tujia and Miao Autonomous Prefecture of West Hunan, Jishou 416000, China)
We investigated the habitat parameters affecting Pteridophytes distribution and analyzed the relationship between Pteridophytes diversity and habitat factors for 37 forest types in the Baiyun Mountain National Nature Reserve.The results showed that:(1)8 609 individuals for Pteridophytes with 90 species,34 genera and 16 families were found in all 251 plots investigated.(2)Habitat factors including forest type,elevation,slope,aspect,soil humus thickness,litter layer depth had a significant influence on Pteridophyte species diversity and community composition.(3)Among these factors inside,forest type and elevation were the most important,especially elevation explaining the Pteridophyte diversity of 11%.These results supported that forest type and elevation were the main reasons for the Pteridophytes distribution,thus we suggested priority given to vegetation type and altitude when formulating Pteridophytes diversity conservation program in the region.
nature reserve; biodiversity conservation; habitat; elevation; understory vegetation
2015-07-20
中国科学院战略性先导科技专项(XDA05050205);自然保护区生物标本资源共享子平台(2005DKA21404)。
Q 142
A
1003 — 5710(2015)05 — 0040 — 06
10. 3969/j. issn. 1003 — 5710. 2015. 05. 010
(文字编校:张 珉)
