若尔盖湿地公路建设生态恢复中植物地上生物量研究
2015-10-28陈蓓马睿秦纪洪等
陈蓓 马睿 秦纪洪等

摘要:在若尔盖湿地段公路建设区,草地生态系统恢复和重建采取了草皮切割贮存并在主体工程完工后将草皮回覆创面进行生态快速恢复的模式,以期实现有效保持水土、构建生态公路的目标。对恢复群落和原生背景群落的地上生物量进行研究,以评价这种生态恢复技术的可行性与有效性。通过对群落种类组成基本性质以及群落生物量与组成植物群落中的种类密度分析,结果表明,优势种对群落性质和环境具有控制性影响,切割草皮与背景样方相比,植物种类有所减少,但优势种组成基本未发生变化;对地上生物量结构波动与变化规律研究结果表明,生物量随季节的变化而变化,在8月中下旬分别达到它们的峰值生物量;对群落的结构分析表明,切割样地有其特定的群落结构和功能。
关键词:若尔盖湿地;公路建设区;人工恢复植被;地上生物量
中图分类号:S157;X171.4 文献标志码:A 文章编号:0439-8114(2015)19-4676-06
DOI:10.14088/j.cnki.issn0439-8114.2015.19.006
Abstract:Grassland ecosystem restoration and reconstruction of highway construction in the Zoige wetland was studied combining artificial turf cutting technology with relocating technology. In this study, the aboveground biomass of the restoration community and the native community was determined to evaluate the feasibility and effectiveness of this technique. By analyzing the species composition of the community and the species density in the plant community, the results showed that the dominant species had a controlling influence on the community nature and the environment. The plant species of turf cutting plots decreased when compared with the control, while the composition of the dominant species were not changeable in turf cutting plots and the control. The results of the research on the fluctuation and variation of the aboveground biomass structure showed that the biomass varied with the seasons. The peak biomass was reached in late August. The structure analysis of the community showed that the cutting plots had specific community structure and the function.
Key words:Zoige Wetland; highway construction zone; ecological restoration; aboveground biomass
国道213线郎川段(郎木寺—川主寺)改扩建工程从若尔盖国家级湿地保护区以及若尔盖大草原通过,全线3/4路段位于海拔3 200~3 800 m的高寒地区,有着种群丰富且保护较好的高寒自然生态系统。鉴于若尔盖高寒草甸草原和高原湿地地处青藏高原东南缘的生态脆弱区的环境敏感性,以及作为长江与黄河上游重要水源地、高原湿地生物多样性重点区域的生态重要性,在设计与建设时充分考虑了有效的生态与工程措施以降低工程对草原与湿地生境的破坏。高寒草甸草粘层形成时间漫长,一旦破坏不但植被难以恢复,而且草粘层丧失会直接导致高寒草甸土壤侵蚀与沙化,因此在工程建设时采用了工程前将施工地表的草皮切块堆放贮藏,主体施工完成后将切块堆放的草皮覆盖工程中破坏的道路两侧地表,从而实现地表植物群落快速恢复和有效保持水土、构建生态公路的目标。这些措施的实施及其效果评价,对于高寒区域工程施工后的生态恢复重建与水土保持具有重要意义。鉴于若尔盖高寒草甸草原和高原湿地的生态脆弱性和生态重要性,如何在公路建设时减小对脆弱的草原、湿地生境的破坏,同时对工程中破坏的植被进行快速恢复和有效保持水土,是郎川公路改扩建工程急需解决的问题之一。
前期的初步试验中表明,通过收集开挖地面的草皮,回覆移植加强湿地和草原创面的生态恢复是可行的[1,2],在川西北高原湿地的工程建设中采取草皮移植快速恢复工程破坏生境的措施在一定程度上取得了成效,前期的研究表明草皮恢复后的植物群落结构稳定且成效与割取草皮厚度、面积大小与面积相关[3],对恢复群落和原生背景群落的相似性指数(Jacard指数和Sorenson指数)开展对比研究发现[4]:取样面积越大、与背景越相似的群落,恢复效果越好;对β多样性指数进行研究表明,β多样性测度指标以物种的个体数作为测度指标将导致较大的误差,而以相对盖度、重要值作为测度指标时则得到较合理的结果。当然,全面评估这种生态恢复技术,还需要对恢复群落和原生背景群落的地上及地下生物量进行研究[5],地上生物量是初级生产力的重要组成部分和表现形式,本研究将初级生产力作为一种评价生态功能的指标,以评价在高寒地带工程建设中通过利用原生草皮剥离后再覆盖进行生态恢复技术的可行性与有效性。
1 材料与方法
1.1 国道213郎木寺—川主寺段概况
试验点地位于郎川路线中北段,属若尔盖县,32°11′N-33°56′N,102°35′E-103°03′E,平均海拔2 900~3 850 m,具青藏高原大陆季风气候特征。冬季(11月至次年4月)主要受西北风冷气团控制,寒冷干燥、日照多而强、日温差大。夏季(5~10月)受西南季风控制,降水明显增多而进入湿季。多年平均气温1.1 ℃,最热月(7月)多年平均气温10.7 ℃,最冷月(1月)多年平均气温-10.7 ℃;极端最高气温24.6 ℃,极端最低气温-33.7 ℃。年平均降雨量656.8 mm,最大年降水量862.9 mm,历年单日最大雨强65.3 mm/d,历年平均蒸发量1 233.2 mm,历年平均相对湿度78%。草地年均日照实际时数2 417 h,年平均风速2.4 m/s。
1.2 样地选择与调查方法
野外调查地点为郎川公路若尔盖花湖地区草皮移栽试验样地,离公路50 m,样方规格如表1。选取不同切割规格(面积和厚度)的草皮回覆道路建设破坏的地表后群落恢复情况的样方6个,背景原生草甸样方1个,分别进行物种调查、密度、高度、盖度、频度的测定,在公路两侧样地(面积为100 cm×100 cm 随机取各植物地上部分50株(个别种为100株),将莎草科和禾草科植物的每一分蘖枝作为一株,测定其生物量(烘干重),并计算出各植物种群平均单株生物量。地上生物量与地下生物量测定同步进行,齐地剪取,样方面积50 cm×50 cm,3次重复。地上净生产量按植物枯黄前(8月底)测定得最高生物量为依据。全部生物量均以烘干记重。
1.3 物种重要值指标
重要值计算公式为:IV=(相对高度+相对盖度+相对密度+相对频度+相对生物量)/5。
2 结果与分析
2.1 样地群落种类组成的基本性质
在样地植物群落研究中,对物种进行逐个登记,主要包括物种调查、密度、高度、盖度、频度、地上生物量(鲜重和干重)的测定,得到一份所研究群落的植物种类名录。群落中的主要植物种群有:羊茅(Festuca ovina Linn)、早熟禾(Poa pratensis)、垂穗披碱草(Elymus nutans)、嵩草(Kobresia)、老鹳草(Geranium pylzowianum)、风毛菊(Saussurea supera)、金莲花(Trollius buddae)、棘豆(Oxytropis sp.)、钝裂银莲花(Anemone sp.)、鹅绒委陵菜(Potentilla anserina)、毛茛(Ranunculus japonica)、薹草(Carex sp.)、川藏蒲公英(Taraxacum maurocarpum)、马先蒿(Pedicularis kansuensis)等。
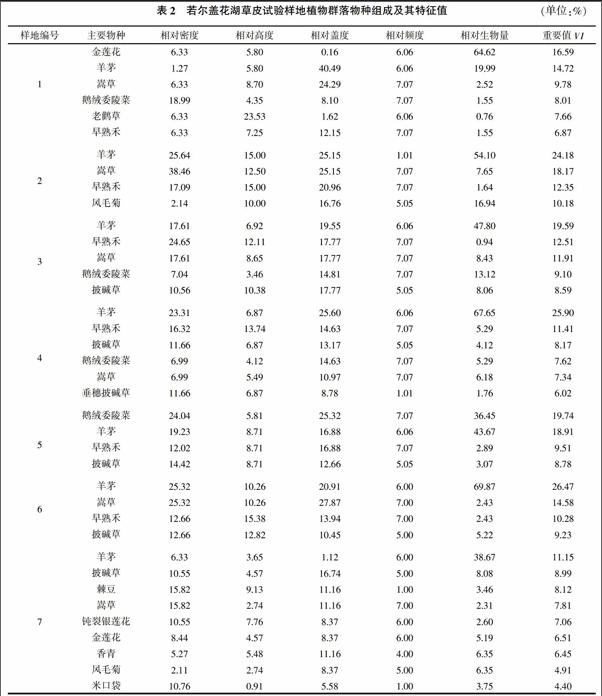
表2列出了每个样地中重要值大于该组平均值的物种及其特征值。调查发现,该区草皮的主要植物有32种,隶属8科28属,在若尔盖花湖草皮试验样地中,植物群落多以耐寒多年生禾本科和莎草科为主,人工移栽草皮上的植物群落除禾本科外,蓼科、菊科和毛茛科、豆科等类群的物种增加,群落的优势种和建群种以羊茅、早熟禾、披碱草和嵩草为主。
本研究样地中优势种以羊茅为主,它们个体数量多、盖度大、生物量高、生活能力强,即优势度大。优势种对群落性质和环境具有控制性影响,如果除去了优势种,群落将发生根本的变化,并影响其他物种生存。亚优势的个体数量和作用都仅次于优势种,但是在决定群落性质和控制群落环境方面仍起着一定作用的植物种。本研究样地亚优势种主要有披碱草、嵩草、早熟禾、金莲花等。伴生种与优势种相伴存在,但不起主要作用。样地中常见的有鹅绒委陵菜、棘豆、车前草、钝裂银莲花、香青、高原毛茛等,它们在群落中数量较少,是常见的伴生种。偶见种是指那些在群落中出现频率很低的种类,多半是由于种群本身数量稀少的缘故。样地中出现的偶见种有狼毒(Stellerachamae jasme)、黄帚囊吾(Ligularia virgaurea)、龙胆(Gentiana spp.)、金露梅(Potentilla fruticosa)等。偶见种可能是偶然的入侵种,也可能是群落中的衰退残遗种。有的偶见种对生态环境的变化具有一定的指示意义。
羊茅在6个试验样地中重要值最大(其平均值为20.13%),为该群落的主要组成成分;次优势种依次是嵩草、早熟禾、披碱草、鹅绒委陵菜、老鹳草、金莲花、垂穗披碱草、棘豆、钝裂银莲花、香青(Anaphalis sp.)等;其余21种植物的重要值均小于种的平均值(7.15%),在整个群落组成中占次要地位,而且多为植株矮小、有毒有害的杂草类,如狼毒。7#样地作为背景样方具有最多的植物种类(23种),优势种是羊茅和披碱草,均为优良牧草;次优势种为棘豆、嵩草、钝裂银莲花、金莲花等。经过切割移栽的草皮与背景样方相比植物种类有所减少,除禾本科和莎草科以外的植物及一年生植物增加,杂草增加,但优势种组成基本未发生变化。
对物种的特征值分析表明,羊茅在6个试验样地的相对密度为1.27%~25.64%,与背景样方(6.33%)相比大部分值均偏高;相对盖度变化范围为19.55%~40.49%,明显高于背景样方的1.12%;相对生物量变化范围为36.45%~69.87%,也高于背景样方的38.67%。其他的主要优势种也具有类似的变化,说明切割的草皮仍然以优良的牧草为主,但切割草皮上优势种的基本指标均高于背景样方。此外,一些偶见种也出现在试验样地中,如鹅绒委陵菜,在背景样方为偶见种,但在各个草皮试验样方内则变成常见种。这可能由于切割的草皮具有边缘效应,草皮边缘的植物长势会好于大片的原生植被。
2.2 群落生物量与组成植物群落中的种类密度分析
群落生物量直接与组成植物群落中的种类密度相关,植物密度关系到植物进行光合作用叶面积的大小。样地中嵩草和羊茅的密度最高,其次是垂穗披碱草、早熟禾等。这些植物多成丛生长,数量虽多,除垂穗披碱草植株较高大外,其余多生长较低矮。薹草、棘豆、马先蒿的植株密度较低,但它们植丛较明显,而且在群落中分布不太均匀。移栽草皮群落与原生群落物种特征值有差异,区内植物群落优势种和建群种以羊茅、披碱草和嵩草为主(表2),羊茅为该群落的主要优势种,其他植物在整个群落组成中占次要地位,且多为有毒有害的杂草类。
切割移栽的草皮与背景样方相比植物种类有所减少,除禾本科和莎草科以外的植物及一年生植物和杂草增加,但优势种组成基本未发生变化。草皮样地除禾本科和莎草科以外的植物和一年生植物增加,表明人为干扰的植物群落在结构与功能方面发生了退化。各物种的特征值与背景样方相比大部分值均偏高,表明切割草皮仍然以优良的牧草为主。另外,样地形状和边界与多样性密切相关,长方形样地(即样带或样条)优于面积相等的正方形样地,长方形样地可能会反映更多的变异情况。因此,还需进一步对不同形状的样地进行比较研究,确定切割草皮的形状、面积或周长对草皮移栽后的恢复状况的影响。
2.3 地上生物量结构波动与变化规律研究
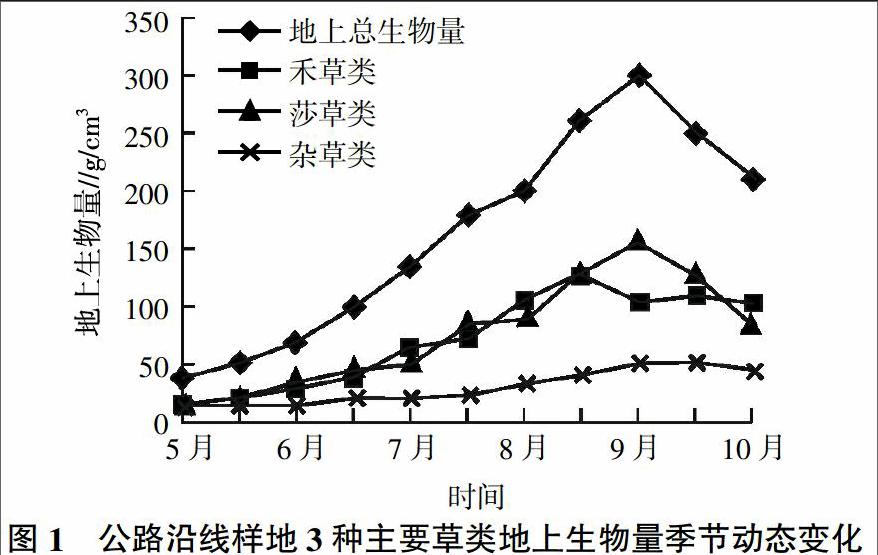
为了便于地上生物量的测定,将组成草甸的植物分为3大类:禾草类、莎草类和杂草类。禾草类主要包括禾本科的羊茅、早熟禾、洽草、垂穗披碱草等;莎草类包括嵩草、薹草等;除上述两类外,其他所有双子叶植物均归于杂草类。
禾草类、莎草类、杂草类生物量随季节的变化而变化(图1)。总体来讲,在8月中下旬分别达到它们的峰值生物量。禾草类先于其他草类达到峰值,而莎草类、杂草类及其地上生物总量均在9月中旬达到峰值。从上述现象分析,地上生物量峰值出现在7月底至8月上旬之后。不同植物类群,因它们的生态-生物学特性各异,禾草类在8月中下旬普遍果实成熟而进入枯黄期;杂草类结实较晚,莎草类枯黄较迟,故峰值生物量出现较禾草类晚,峰值生物量出现后,因气温降低,生物量逐渐下降,直到枯黄时停止。
地上生物量的累积动态,还可以从5~9月生物量的生长强度来说明(表3)。生长强度是指在某一生长期内,平均单位时间内生物量的生长率。从表3可以看出,在相同生长期内,不同的草类生长强度不一,如在刚返青后的4月25日至5月30日期间,禾草类的生长强度[0.538 9 g/(m2·d)]高于杂草类[0.444 4 g/(m2·d)]和莎草类[0.405 6 g/(m2·d)]。表3还说明,同一类植物在不同生长期生长强度不一,如禾草类,在4月25日至5月30日返青期,生长强度为[0.538 9 g/(m2·d)],到5月31日至6月15日拔节期间为[1.067 5 g/(m2·d)],而在7月16至7月30日孕穗期和8月16日至30日成熟期间,生长强度达到最大,分别为[2.231 3 g/(m2·d)]和[2.410 0 g/(m2·d)]。各类草生物量所占总生物量的百分比也随着生长季节而变化。禾草、莎草类因返青较早,它们分别占总生物量的38.8%和29.2%,杂草占32%。随着禾草类、杂草类生长发育,它们的比例不断上升,而莎草类则相应下降;枯黄前,禾草类为52.24%,莎草为14.16%,而杂草为33.6%。枯黄前禾草比例较高,因为禾草类植株较高(一般为30~40 cm),从拔节、孕穗、抽穗到子粒成熟,生物量相应增加较快。杂草类植株较少,但植株平均个体单枝重量较高,特别是到了7月中旬以后,一些杂草类相继开花,植株生长旺盛,生物量增加较快。莎草类植物生长稠密而低矮,优势种嵩草高5~8 cm,属于寒冷中生和冷旱中生植物,返青早,6月中旬已进入盛花期;生长早期,生物量较高,后来随着禾草类、杂草类迅速生长,相比之下,它们占总生物量的比率相应下降。
不同切割草皮样地间的地上生物量差异见表4,背景样方为34.65 g/cm3,在切割草皮中样地1地上生物量最大,达51.53 g/cm3,其次是样地4、样地5样地6,分别为34.00、27.71、25.58 g/cm3;而样地3、样地2的地上生物量最小,分别为18.3 g/cm3和10.67 g/cm3。造成这种差异的主要原因是不同样地植物种类、年龄结构、密度、环境因子及人为活动干扰的不同。
2.4 群落的结构分析
2.4.1 垂直结构 草地群落的垂直结构主要指群落的分层现象,也称为群落的成层性。草地有其特定的群落结构,比较简单,并与其功能相联系。
第一层高20~10 cm,由紫羊茅、嵩草、高原毛茛、圆穗蓼、珠芽蓼、龙胆科、马先蒿、高原紫菀、花锚、鹅鹳草、草玉梅、莎草、发草等组成。
第二层高10~5 cm,由垂穗披碱草、早熟禾、金莲花、钝裂银莲花、棘豆、蒲公英、黄帚橐吾、狼毒、车前草、老鹳草等组成。
第三层高5 cm以下,由鹅绒委陵菜、风毛菊、香青等组成。
群落的垂直结构不仅表现在地上部分,地下的根系也有明显的成层性,不同种类的根系可分布在不同的土层深度。但是,最大根量分布在土壤的表层,这与土壤养分的分布是一致的。在本研究样地中,表层土壤有大量草根且盘结紧实形成草毡层,而10~20 cm土壤中有少部分疏松的根系,20~30 cm土壤中则只有很少量根系。
群落的垂直结构具有提高有限资源的时空利用范围和利用效率的意义,因此有利于提高群落的生产力水平;减少物种为争夺养分和空间资源的竞争,使群落拥有更高的的物种多样性;成层性提高了生物群落的稳定性和对环境的改造作用,增强了抵御外界干扰的能力;成层性的复杂程度是生态环境的一种良好指示,一般层次越复杂,极端环境中的生物群落越简单。
2.4.2 水平结构 群落的水平结构是指群落的水平配置状况或水平格局。本研究中的草皮形成小的群落,生长发育好,有时还可以生长一些越带分布的植物,并形成一定数量的斑块。每一个斑块是一个小群落,它们彼此组合形成群落的镶嵌性水平结构。自然界中群落的镶嵌性是绝对的,而均匀性是相对的。
2.4.3 时间结构 群落的外貌常随着时间的推移而发生周期性的变化,这是群落结构的另一个重要特征,即时间结构。在研究区域草原群落中,一般在5~8月水热充沛,植物繁茂生长,百花盛开,色彩丰富;秋末植物开始干枯休眠,呈红黄相间的秋季季相。冬季则是一片枯黄,或被白雪覆盖。
3 小结与讨论
地上生物量是初级生产力的重要组成部分和表现形式,本研究我们将初级生产力作为一种评价生态功能的指标。Pfistere等[6]利用植物地上生物量来度量群落生产力,地上生物量是总生物量的一部分,但它所占的比例不是一成不变的,因为在不同环境中植物会有不同的资源分配对策。对人工恢复高寒草甸的植物群落多样性与地上生物量的研究表明,物种丰富度依次增加,地上生物量依次降低。植株的大小关系到物种对水分、光照的竞争能力,和对其他种的生存能否起到抑制作用。嵩草、羊茅、垂穗披碱草、早熟禾等这些植物多成丛生长、数量多,除垂穗披碱草植株较高大外,其余多生长较低矮。
在高寒环境中,水分成为限制性因素,根系竞争会增强[7],植物分配在根系生长的资源所占比例上升。所以在受到扰动时,地上生物量占总生物量的比例下降,这种下降趋势在物种丰富的群落中更明显,是因为这些群落有更强烈的种间根系竞争。所以对于不同物种多样性的群落,地上生物量占总生物量的比例不同。从地上生物量变化得出的结果无法反映总生物量的变化。在这项研究中,物种丰富的群落在受到草皮切割扰动时,地上生物量占总生物量比例可能会低于物种少的群落,前者生产力的降低值被高估了。
随物种多样性上升,群落生产力呈升高趋势。Tilman等[8]综合该项试验多年的数据发现:生产力随多样性上升而升高,多样性的这种正效应逐年增强;混合群落发生超产的比例以及超产的强度逐年提高,这表明生态位互补的作用随时间推移而强化。
参考文献:
[1] 陈 蓓,孙 辉,秦纪洪,等.朗川公路若尔盖湿地段人工移栽草皮的群落生态学研究[J].水土保持研究,2007,14(2):68-71.
[2] 陈 蓓,孙 辉,秦纪洪,等.高寒地区道路工程的生态效应及其生态恢复研究进展[J].四川环境,2009,28(2):102-105.
[3] 马 睿,孙 辉,陈 蓓,等.若尔盖郎川公路建设区有毒有害植物扩散及生态安全性评价[J].世界科技研究与发展,2006,28(3):70-75.
[4] 陈 蓓,马 睿,秦纪洪,等.若尔盖湿地公路建设生态恢复中植物群落多样性与相似性研究[J].水土保持研究,2013,20(4):115-119.
[5] 杨兆平,欧阳华,宋明华,等.青藏高原多年冻土区高寒植被物种多样性和地上生物量[J].生态学杂志,2010,29(4):617-623.
[6] PFISTERER A B,SCHMID B. Diversity-dependent production can decrease the stability of ecosystem functioning[J]. Nature, 2002,416:84-86.
[7] WILSON MV, SCHMIDA A. Measuring beta diversity with presence-absence data[J]. J Ecol, 1984,72:1055-1064.
[8] TILMAN D. Functional diversity[A]. Encyclopedia of Biodiversity (Vol.3)[C]. New York: Academic Press, 2001.
