七鳃鳗鳃黏膜免疫系统类淋巴细胞免疫应答遗传基础
2015-10-22刘欣宋雪萤张晓萍韩英伦朱婷肖蓉李庆伟
刘欣,宋雪萤,张晓萍,韩英伦,朱婷,肖蓉,李庆伟
1. 辽宁师范大学生命科学学院,大连 116081;
2. 辽宁师范大学七鳃鳗研究中心,大连 116081
七鳃鳗鳃黏膜免疫系统类淋巴细胞免疫应答遗传基础
刘欣1,2,宋雪萤1,2,张晓萍1,2,韩英伦1,2,朱婷1,2,肖蓉1,2,李庆伟1,2
1. 辽宁师范大学生命科学学院,大连 116081;
2. 辽宁师范大学七鳃鳗研究中心,大连 116081
近年来,在无颌类脊椎动物七鳃鳗体内发现了以可变淋巴细胞受体(Variable lymphocyte receptors, VLR)为基础的抗原识别机制。为揭示七鳃鳗鳃黏膜免疫系统中类淋巴细胞适应性免疫应答的遗传基础,探索无颌类与有颌类脊椎动物在适应性免疫应答机制上的进化关系,本文构建了日本七鳃鳗(Lampetra japonica)鳃囊组织免疫前后cDNA文库并进行了高通量转录组测序及分析。通过对组装得到的88 525个独立基因(Unigene)进行功能注释,分别有21 704和9769个unigene在GO(Gene Ontology)和KEGG(Kyoto Encyclopedia of Genes and Genomes)数据库得到注释。999个unigene参与免疫系统的多个通路,其中184个与高等脊椎动物TCR(T cell receptor)和BCR(B cell receptor)信号通路的51个分子具有较高的同源关系,说明七鳃鳗体内存在高等脊椎动物适应性免疫应答信号通路的相关分子。本文还发现5个VLRA、7个VLRB和4个VLRC分子,说明七鳃鳗鳃黏膜免疫组织内至少分布3种类淋巴细胞亚群。实时荧光定量PCR结果显示,Lck、Fyn和Zap70基因在免疫激发后表达量显著上调,而Syk、Btk和Blnk基因表达没有显著变化,说明七鳃鳗鳃组织受到抗原刺激后,类似T淋巴细胞的信号转导途径被激活。本研究初步证明,尽管无颌类和有颌类脊椎动物的适应性免疫系统在抗原识别机制上存在不同,但具有共同的遗传基础。研究结果为探讨七鳃鳗VLRA+、VLRB+和VLRC+淋巴细胞免疫应答信号传导过程提供了有价值的线索。
日本七鳃鳗;鳃黏膜免疫组织;转录组;免疫应答
黏膜免疫是免疫系统的一个重要组成部分,为生物体的各种黏膜提供保护,使其免受潜在病原微生物的入侵并调节机体的免疫反应[1]。鱼类生活在水环境中,其皮肤、鳃和肠道与水体直接接触,是病原侵入鱼体的门户。为免受病害侵袭,在长期进化过程中,这些组织和器官形成了黏膜相关淋巴组织(Mucosal associated lymph tissue, MALT)。尽管负责气体交换的鱼鳃由几对鳃片组成,但鳃片上排列的梳齿状鳃丝具有巨大的表面积,使其成为鱼类重要的黏膜免疫组织。Lin等[2]在大西洋鲑(Salmo salar)和黄盖鲽(Limanda limanda)鳃组织中发现大、小淋巴细胞、巨嗜细胞、中性粒细胞、嗜酸性粒细胞等免疫细胞的存在。当鱼体受抗原刺激时,巨噬细胞可以对抗原进行处理和呈递,抗体分泌细胞会分泌特异性抗体,构成了机体与外界相接触的第一道免疫防护体系[3]。从鳃淋巴细胞对有丝分裂原脂多糖(Lipopolysaccharides, LPS)和植物血凝素(Phytohaemagglutinin, PHA)的反应来看,PHA比LPS对鳃淋巴细胞的激活作用更加显著,说明分布在鱼类鳃中的淋巴细胞以T细胞为主[4]。
七鳃鳗属于脊椎动物门、圆口纲(Cyclostomata)、七鳃鳗目(Petromyzontiformes)。长久以来,七鳃鳗作为脊椎动物中最低等的鱼形动物,因其在进化上的特殊地位在比较解剖学[5]、发育神经生物学[6]、生理学[7]和免疫学[8]等研究领域备受关注。近年来,Pancer等[9,10]发现海七鳃鳗(Petromyzon marinus)和太平洋粘盲鳗(Eptatretus stoutii)存在以可变淋巴细胞受体(Variable lymphocyte receptors, VLR)为基础的抗原识别机制,提出无颌类存在一种平行于有颌脊椎动物以T细胞受体(T cell receptor, TCR)、B细胞受体(B cell receptor, BCR)识别抗原的适应性免疫系统。之后,共有3种VLR(VLRA、VLRB和VLRC)被发现,且分别存在于不同的淋巴细胞亚群上(VLRA+、VLRB+和VLRC+淋巴细胞)[11]。Guo等[12]发现在七鳃鳗的鳃囊中存在小淋巴细胞,并且VLRA+和VLRB+淋巴细胞的数量比值在鳃囊中接近1,远高于其他组织,说明VLRA+淋巴细胞主要分布在鳃囊中。VLRA+淋巴细胞在PHA刺激后的反应方式与高等脊椎动物的T细胞相似。刺激后的VLRA+淋巴细胞特异性表达分子包括转录因子GATA2/3(GATA binding protein 2/3)、原癌基因c-Rel(v-rel avian reticuloendotheliosis viral oncogene homolog)、芳香烃受体Ahr(Aryl hydrocarbon receptor)、B-细胞淋巴瘤因子Bcl11B(B-cell CLL/lymphoma 11B);趋化因子受体CCR9 (Chemokine (C-C motif) receptor 9);T细胞命运决定分子Notch1(Neurogenic locus Notch homolog protein 1);酪氨酸磷酸酶CD45;白细胞介素IL-8(Interleukin 8)受体和趋化因子受体CXCR2(Chemokine (C-X-C motif) receptor 2);促炎细胞因子IL-17和巨噬细胞移动抑制因子MIF(Macrophage migration inhibitory factor)。但是,由于七鳃鳗体内不存在以组织相容性复合体 (Major histocompatibility complex, MHC)、TCR、BCR和免疫球蛋白(Immunoglobulin, Ig)等分子为主导的适应性免疫机制,其黏膜免疫系统中类淋巴细胞适应性免疫应答的遗传基础及其与高等脊椎动物适应性免疫应答的异同目前尚未见报道。
本研究以日本七鳃鳗(Lampetra japonica)这一东亚孑遗活化石为研究对象,利用高通量测序技术对其免疫前后的鳃囊组织进行了转录组测序(RNA-seq)。通过对序列进行比较分析,搜索和识别无颌类脊椎动物参与适应性免疫应答的相关分子,揭示无颌类黏膜免疫系统类淋巴细胞适应性免疫应答的遗传基础,探究与高等脊椎动物适应性免疫系统之间在进化上的关系。
1 材料和方法
1.1材料
日本七鳃鳗捕自黑龙江省松花江流域同江地区,活体置于实验室水族箱驯养(2~5℃)。1周后选取24条健康的个体(体长35~55 cm),分为两组:对照组和实验组。实验组以复合抗原浸泡免疫1 h方式进行免疫刺激[13]。复合抗原用市售生理盐水配制,含灭活的大肠杆菌(Escherichia coli DH5α) (TaKaRa,Dalian)、金黄色葡萄球菌(Staphylococcus aureaus)(宝生物工程(大连)有限公司)和啤酒酵母菌(Saccharomyces cerevisiae)(市售)各1×109个/mL,免疫时用水族箱养殖水稀释3 000倍后使用。实验组经过2次加强免疫(每2周1次),末次免疫后3 d取其新鲜的鳃组织。对照组不用复合抗原,其余的处理方法同实验组。
1.2方法
1.2.1RNA样品提取及cDNA文库的构建和测序
使用RNAiso Reagent(TaKaRa, Dalian)提取日本七鳃鳗复合免疫实验组及对照组鳃囊总RNA。获得符合测序标准的总RNA 样本后,委托北京诺禾致源生物信息科技有限公司进行等量混合后构建转录组测序文库并用Illumina HiSeqTM2000进行双向测序(Paired-end sequencing)及分析。
1.2.2转录本的拼接
测序得到的原始序列需去除含N(N表示无法确定碱基信息)的比例大于10%的读数(Reads)、带接头的读数及低质量的读数(质量值Q≤5的碱基数占整个读数长度50%以上的读数),从而得到纯净读数(Clean reads)。采用针对短序列拼接的Trinity软件进行拼接(版本:v2012-10-05),min_kmer_cov参数设置为2,其他参数设置为默认[14]。最后,将拼接至两端不能再延长的转录本作为独立基因(Unigene)。
1.2.3转录组数据处理与注释分析
利用Blast软件(NCBI blast 2.2.28+),将Unigene在Nr蛋白序列数据库中进行序列比对。未注释的Unigene在Nt核酸序列数据库中搜索同源序列。为减少错配比例,将e值(e-value)的阈值设为小于1E-10。
运用HMMER3程序(http://pfam.sanger.ac.uk/),根据Pfam(Protein family)数据库[15]中已构建好的蛋白结构域HMM(Hidden Markov Models)统计模型,对Unigene进行蛋白质结构域的注释。
利用Blast2GO[16]和KAAS[17](http://www. genome.jp /tools/kaas/)软件,基于Nr和Pfam两部分的蛋白注释结果对Unigene分别进行Gene Ontology(GO)和KEGG(http://www.genome.jp/kegg/)通路功能注释及分类统计。
1.2.4基因差异表达分析
1.2.5实时荧光定量PCR分析
选取与TCR、BCR相邻的7个重要的信号分子(酪氨酸蛋白激酶Lck、Fyn、Tec、ZAP-70,脾酪氨酸激酶Syk,B细胞连接蛋白Blnk和布鲁顿酪氨酸激酶Btk)进行实时荧光定量PCR分析验证。鳃组织总RNA样品的提取按方法1.2.1进行。采用Primescript™RT试剂盒(宝生物工程(大连)有限公司)进行反转录。使用TaKaRa SYBR®primescript™RT-PCR(宝生物工程(大连)有限公司)试剂盒,按照产品说明书进行实时荧光定量PCR,每个样品重复实验3次。以七鳃鳗GAPDH的表达水平作为内参,对实验结果进行校准。各实验组结果用对照组结果进行归一化计算相对表达水平。PCR扩增程序为:95℃预变性5 s;95℃ 5 s,60℃ 5 s,72℃ 30 s,40个循环;最后再65℃延伸15 s。运用SPSS统计软件包的t检验模块对基因表达进行差异显著性分析,P<0.05为差异具有统计学意义。所验证的7个蛋白激酶的PCR引物序列见表1,引物由宝生物工程(大连)有限公司合成。
2 结果与分析
2.1七鳃鳗鳃转录组测序序列的转录本拼接及注释
对所构建的对照组和实验组两个cDNA文库进行双向测序分别得到的原始数据为25 947 041和28 841 887 对原始读数(Raw reads)。通过对原始读数过滤,分别得到24 382 693和27 004 299 对纯净读数(序列总碱基数分别为4.88 G和5.4 G个碱基),Q20和Q30(Phred 数值大于20、30的碱基占总体碱基的百分比)指标分别高于96%和90%,说明测序结果良好。利用Trinity软件对2个转录组双向4组纯净读数集合进行混合拼接,最终在所组装的119 004个序列重叠群(Contig)中筛选出88 525个Unigene,总长度69 291 924 bp,平均长度671 bp,N50值为1 092 bp。在这些Unigene中,序列长度在300 bp以下的有28 283个,占31.95%;300 ~ 499 bp的有25 072个,占28.32%;500 ~ 999 bp的有17 952个,占20.28%;1000~1 999 bp的有9728个,占10.99%;
长度在2 000 bp以上的有7 490个,占8.46%。按照转录组数据处理与注释分析方法对88 525个Unigene进行了功能注释,占总数29.43%的26 049个Unigene至少在一个数据库中得到注释。在PFAM和GO数据库得到注释的比例较高,分别有20 834个(24.52%)和21 704个(23.53%)Unigene;在Nr、KEGG和Nt数据库中被注释的相对较少,分别有16 879(19.07%)、9 769(11.04%)和8 084个(9.13%)。
2.2Unigene的GO分类
对21 704个获得GO注释的Unigene按照生物学过程、细胞组分和分子功能进行了分类,结果见图1。在生物学过程23个小类中,归入Unigene数量较多的是细胞过程(13 197个,9.46%)、代谢过程(10 410个,7.46%)和单有机体过程(8 329个,5.97%)。分别有3 051个(2.19%)、338个(0.24%)和44个(0.03%)Unigene归入信号传递、免疫系统过程和细胞的杀伤等与免疫相关的类别。在细胞成分的18个注释小类中,细胞(8 394个,6.02%)和细胞组件(8 344个,5.98%)归入的Unigene较多;突触(96个,0.07%)、突触组件(85个,0.06%)和细胞核(6个,0.00%)归入的Unigene较少。细胞成分注释只反映被注释分子在细胞中的位置,因此不能直观反映该分子是否参与免疫过程。在分子功能注释中,结合(12 050个,8.64%)归入的Unigene最多,其次为催化活性(7 646个,5.48%)和转运活性(1 650个,1.18%),而较少的类别有抗氧化活性(35个,0.03%)、受体调节活性(34个,0.02%)和金属伴侣活性(8个,0.01%)。GO分类结果在一定程度上反映出七鳃鳗鳃组织中所表达的基因的位置分布、所参与的生物学过程及可能行使的功能属性,为后续进行免疫相关基因的筛选和功能富集分析奠定了基础。

表1 实时荧光定量PCR扩增所用引物
2.3Uni gene的KEGG通路注释
对Unigene进行KO注释后,根据它们参与的KEGG通路进行分类,结果见表2。9769个被注释的Unigene分别属于细胞过程、环境信息加工、遗传信息加工、代谢和有机体系统这5个一级分类群下的260个标准生物学通路(表2)。在二级分类群中,信号转导中的Unigene最多,共有2 914个;其他较多的有内分泌系统(1 506个)、神经系统(1324个)、免疫系统(999个)、细胞通讯(775个)、运输和催化(642个)及信号转导分子和相互作用(576个)。
参与免疫系统各通路的999个Unigene中,146个参与细胞因子信号通路(Chemokine signaling pathway,ID:ko04062);128个参与白细胞跨血管壁迁移(Leukocyte transendothelial migration, ID: ko04670);124个参与Fc γR介导巨噬细胞吞噬作用(Fc gamma R-mediated phagocytosis,ID:ko04666);104个参与TCR信号通路(T cell receptor signaling pathway, ID:ko04660);84个参与抗体依赖细胞介导的细胞毒作用(Natural killer cell mediated cytotoxicity, ID: ko04650);80个参与BCR信号通路(B cell receptor signaling pathway,ID:ko04662);70个unigene参与Toll样受体信号通路(Toll-like receptor signaling pathway,ko04620);52个unigene参与FcεRI信号通路(Fc epsilon RI signaling pathway,ID:ko04664)。还有少量unigene参与天然免疫的分子通路,如:补体凝集反应(Complement and coagulation cascades,ID:ko04610)、RIG-I 样受体信号通路(RIG-I-like receptor signalingpathway,ID:ko04622)和NOD 样受体信号通路(NOD-like receptor signaling pathway,ID:ko04621)。以上结果显示,七鳃鳗也存在高等脊椎动物天然免疫和适应性免疫应答信号通路的相关分子,说明在一定程度上这两种平行的适应性免疫系统具有共同的遗传基础。
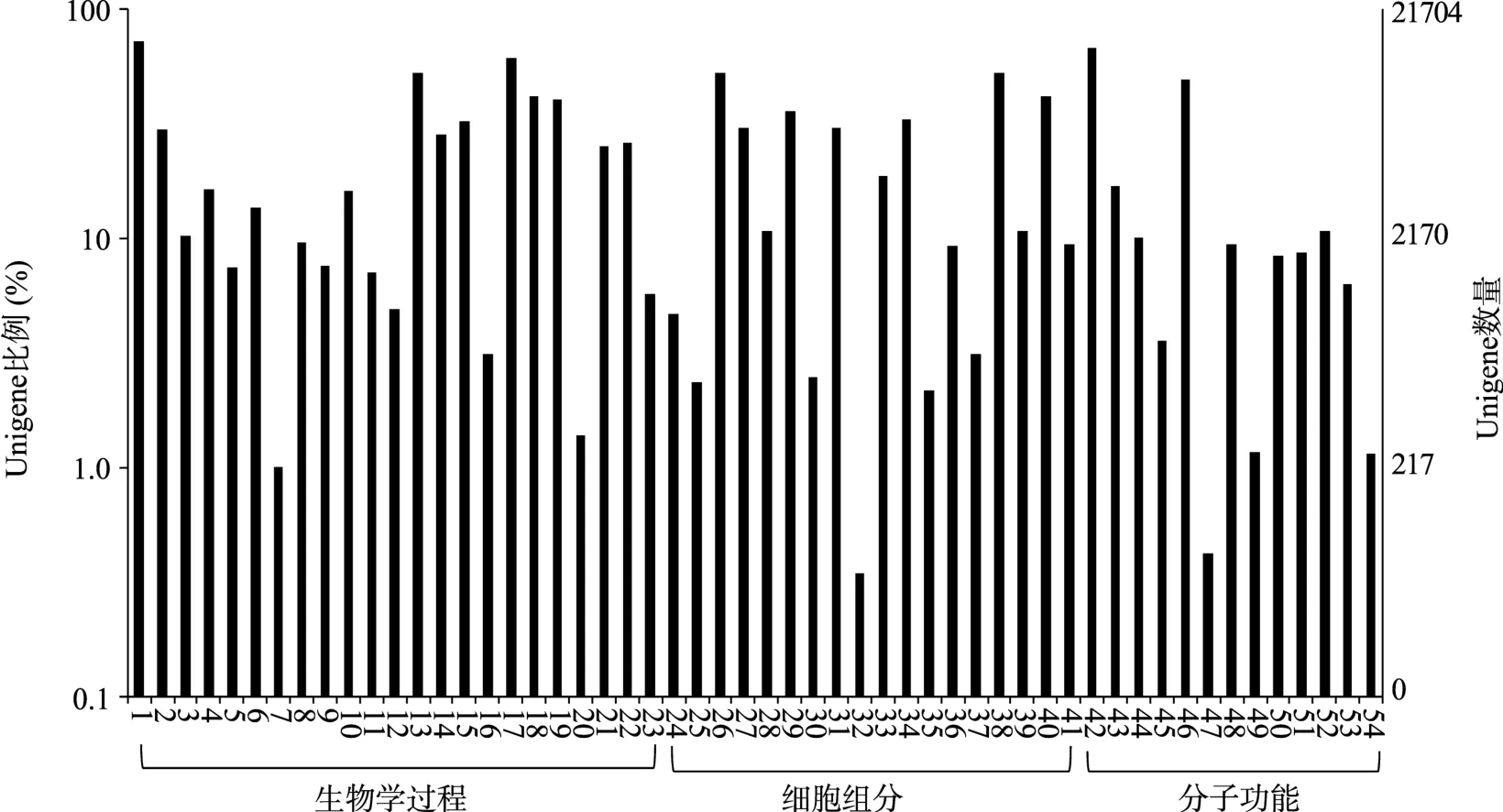
图1 Unigene的GO分类图
2.4七鳃鳗鳃黏膜适应性免疫应答分子的甄别
对注释为参与TCR和BCR信号通路的184个Unigene进行分析,发现它们与51个高等脊椎动物参与TCR和BCR信号通路的分子有较高的同源关系。对这184个Unigene进行归类比较,其中有51个Unigene序列长度较长、同源匹配e值较高(表3)。其中,编号1~23号分子在TCR和BCR信号通路中都行使功能,24~43号分子属于TCR信号通路,而剩下的8个分子属于BCR信号通路。并且,有14个Unigene在复合抗原刺激后在转录水平出现明显上调。另外,本文也从注释的Unigene中甄选到5个VLRA、7个VLRB和4个VLRC分子(表4);还有100多个Unigene在Nt数据库被注释为VLR分子,但由于序列较短不能被准确确定类别,所以本文未给出结果。3种VLR分子的发现,说明七鳃鳗鳃黏膜免疫组织内分布着所有的类淋巴细胞亚群,而这些分子的发现为将来探讨VLRA+、VLRB+和VLRC+淋巴细胞免疫应答信号传导过程提供了有价值的线索。
2.5T、B细胞信号转导通路关键蛋白激酶的验证
在筛选的参与TCR和BCR信号通路的7个蛋白激酶中,酪氨酸蛋白激酶Lck、Fyn、Tec和ZAP-70
参与T细胞信号转导通路,而B细胞连接蛋白(Blnk)、布鲁顿酪氨酸激酶Btk和脾酪氨酸激酶Syk参与B细胞信号通路。利用实时荧光定量PCR对这些蛋白激酶的mRNA在免疫前后的表达情况进行验证,发现Lck、Fyn和Zap70在免疫后表达量显著上调,而Syk、Btk和Blnk没有显著变化(图2)。结果说明,七鳃鳗鳃组织在受到抗原刺激后,类似T淋巴细胞的信号转导途径被激活。
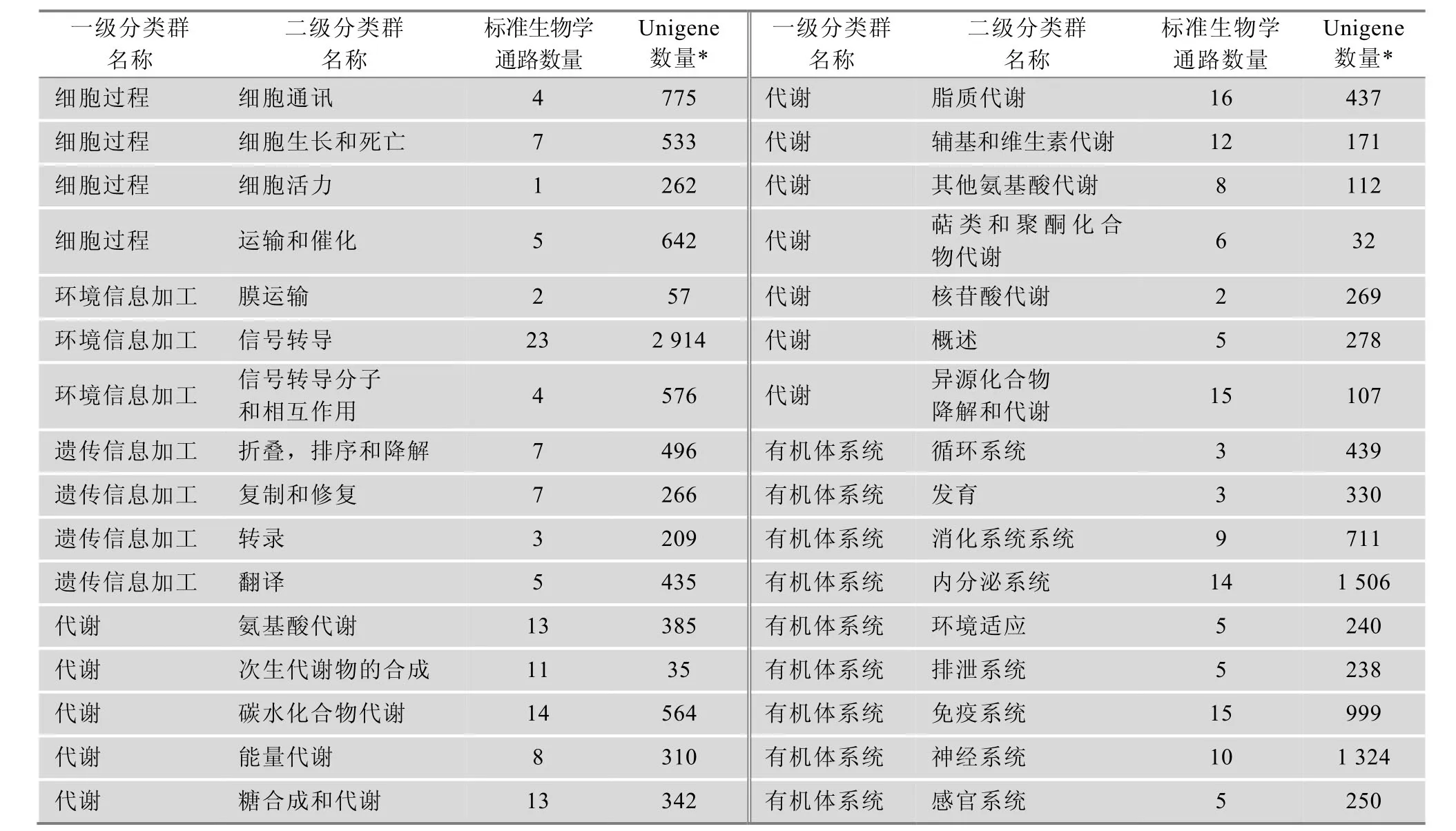
表2 KEGG通路注释结果统计表
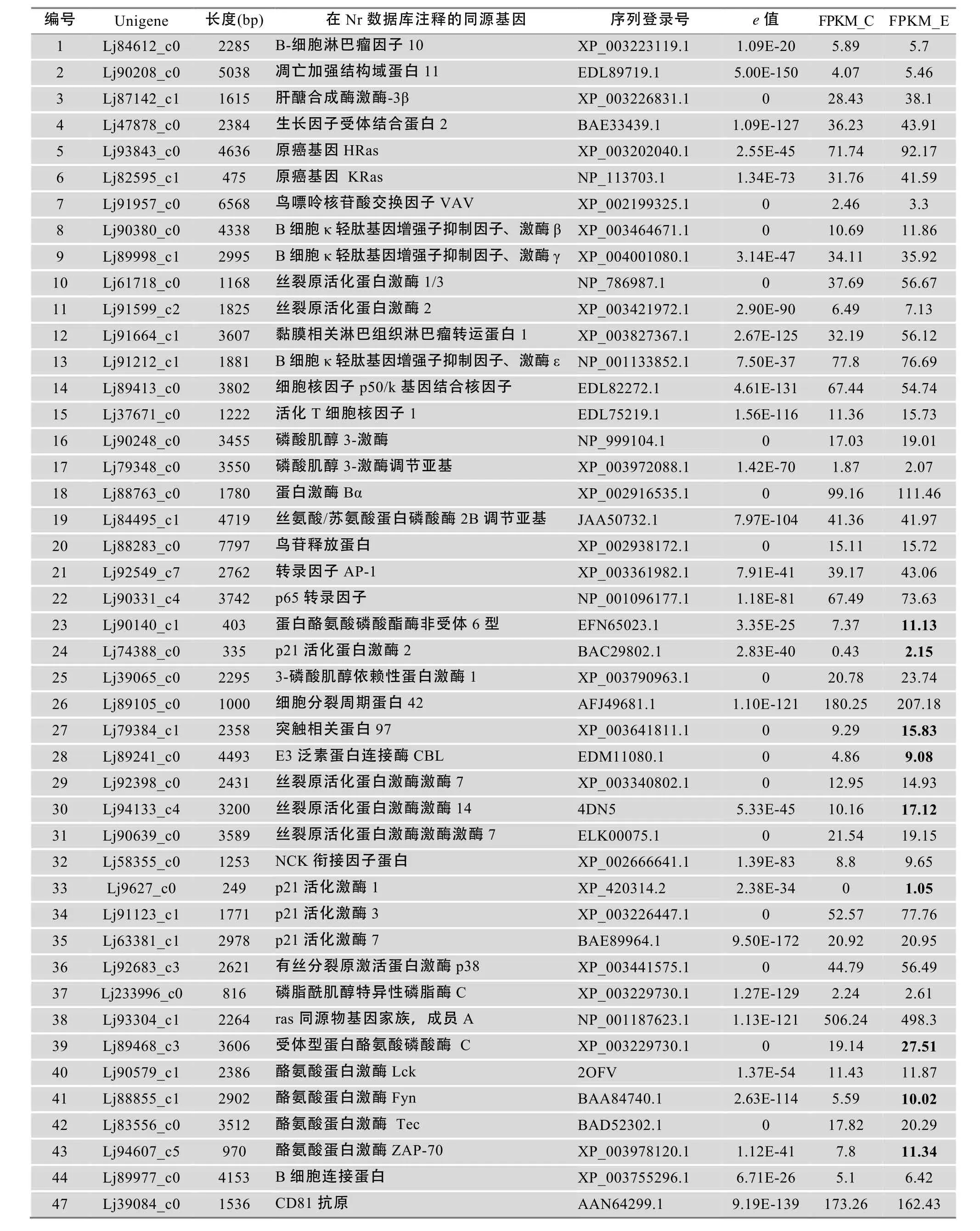
表3 与T、B细胞受体信号通路相关分子同源的信号分子

续表3
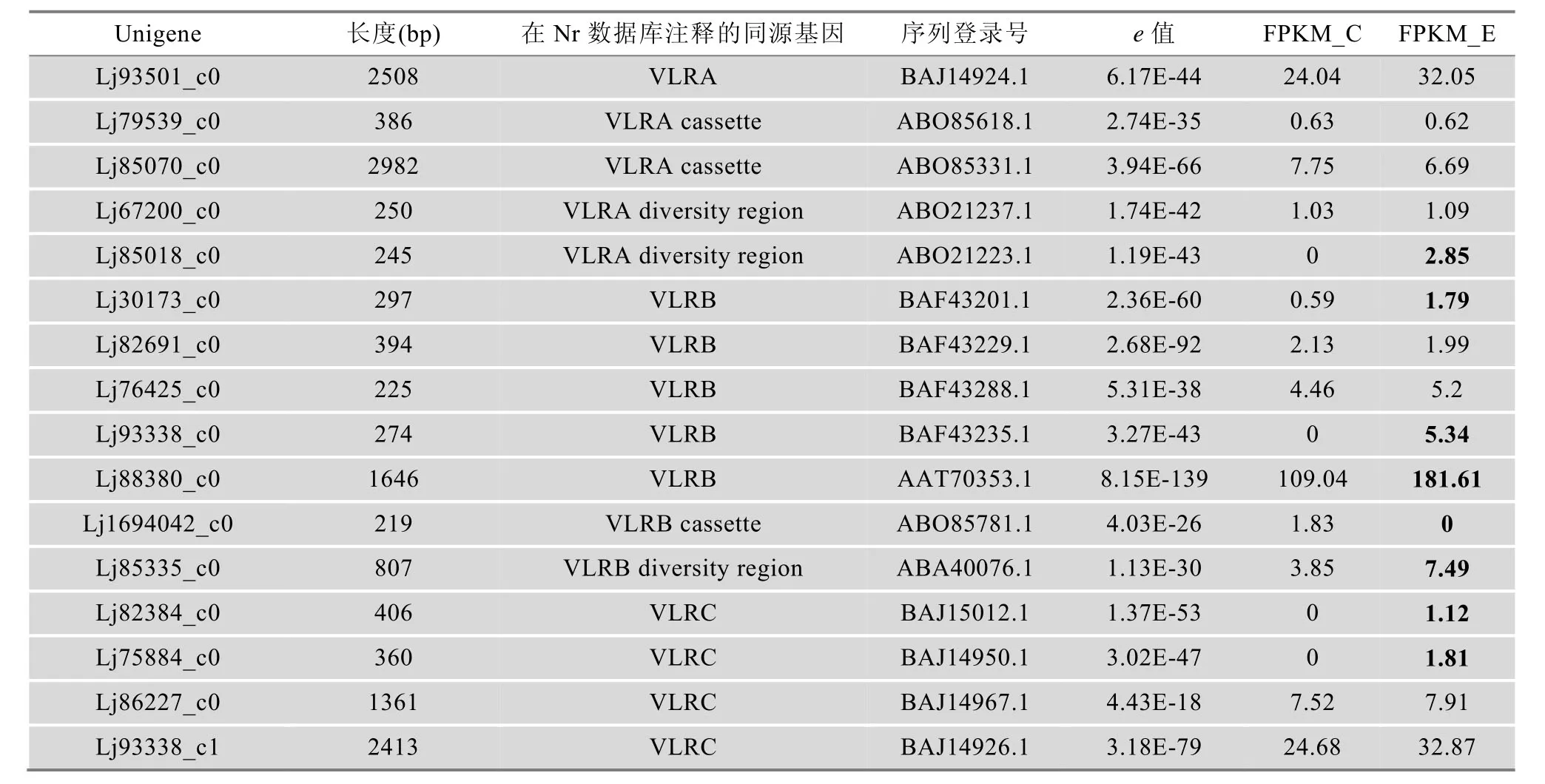
表4 鳃中表达的可变淋巴细胞受体(VLR)的类型
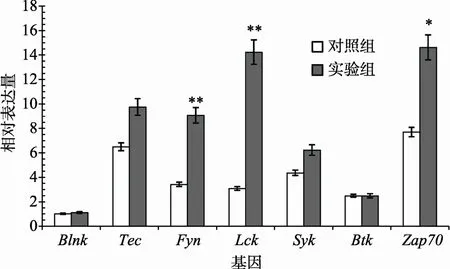
图2 7种蛋白激酶mRNA表达情况的实时荧光定量PCR验证
3 讨 论
长期以来,人们普遍认为高等脊椎动物的适应性免疫系统是由无脊椎动物的先天性免疫系统直接进化而来[24]。随着研究的深入,发现以七鳃鳗和盲鳗为代表的古老的无颌类脊椎动物存在类似脊椎动物所具有的适应性免疫现象,同时还存在许多与脊椎动物适应性免疫分子同源的分子[25]。而VLR及其重组机制的发现,挑战了先天性免疫进化学说;这种在无颌类脊椎动物中富含亮氨酸重复序列(Leucine-rich repeat)插入的重组机制,极有可能是现代适应性免疫的起源。因此,作为研究脊椎动物起源与进化的模式动物,七鳃鳗也逐渐成为探讨适应性免疫起源与进化的关键物种。本研究成功构建了日本七鳃鳗黏膜免疫重要组织鳃的cDNA文库,通过转录本的注释,探索和发现日本七鳃鳗鳃组织适应性免疫相关分子的组成及表达。本文为进一步研究七鳃鳗鳃黏膜组织各淋巴细胞亚群适应性免疫应答的机制,以及适应性免疫系统的起源与进化提供了有价值的线索。
高等脊椎动物的适应性免疫应答反应是指抗原特异性T、B细胞受抗原刺激后,从自身活化开始到增殖、分化为效应细胞,产生一系列生物学效应的全过程。参与此过程的信号通路主要有TCR和BCR信号通路。TCR是一种T细胞膜蛋白复合物,与CD3分子一起形成T细胞受体复合体来参与抗原呈递时T细胞的激活。由于TCR具有两种类型(TCRαβ和TCRγδ),T细胞也相应存在两种亚群(αβ和γδ T 细胞)[26]。与有颌类脊椎动物相比,无颌类脊椎动物的适应性免疫系统尚没有进化出完善的MHC抗原呈递机制,也没有发现TCR及BCR存在。但是,Guo等[12]发现在七鳃鳗中存在VLRA+和VLRB+两种淋巴细胞亚群,其免疫反应类似于T和B细胞;Hirano等[27]发现在七鳃鳗鳃囊中除了存在少量VLRB+类淋巴细胞外,还存在VLRA+和VLRC+两种类淋巴细胞,它们除了分别特异性表达VLRA和VLRC外,也分别表达高等脊椎动物αβ和γδ T 细胞分化相关的一些直系同源基因。因此,该研究提出VLRA+和VLRC+两种类淋巴细胞在起源上与αβ和γδ T 细胞有共同的祖先。本研究也发现在七鳃鳗的鳃囊中有VLRA、VLRB和VLRC3种受体分子的表达(表4),说明七鳃鳗鳃黏膜免疫组织内有3种类淋巴细胞亚群分布。由于鳃囊中毛细血管丰富,本文检测到相对较多数量的VLRB受体的原因可能是由于外周血中VLRB+类淋巴细胞比例较高所造成的。尽管此前的研究在七鳃鳗中发现VLRA+、VLRC+及VLRB+3种类淋巴细胞在起源上类似于有颌类脊椎动物的αβ、γδ T及B细胞,但参与这3种VLR受体信号转导的分子基础有哪些尚未涉及。本研究发现七鳃鳗鳃组织存在与高等脊椎动物T、B细胞信号通路51个信号分子同源的Unigene(表3)。其中,有23个分子为T、B信号通路所共有,另外分别有20和8个分子为T和B细胞信号通路所特有。这些结果反映出无颌类和有颌类脊椎动物适应性免疫应答尽管在机制上不同,但在一定程度上具有共同的遗传基础。
T细胞被MHC递呈的抗原肽激活后,CD3ζ链的免疫受体酪氨酸激活基序(Immunoreceptor tyrosine-basedactivation motif, ITAM)的酪氨酸残基被胞内的酪氨酸蛋白激酶Lck磷酸化,然后募集其他含有SH2(Scr homology 2)结构域的酪氨酸蛋白激酶(Fyn和Zap-70等),并诱导一系列细胞内信号级联反应[26]。BCR依靠B细胞外膜的免疫球蛋白受体分子识别抗原,然后将细胞外信号通过跨膜的CD79α/CD79β异二聚体胞内结构域的ITAM基序传导进胞内。后者通过募集激酶Lyn、Syk及PI3K和接头分子GAB1和BLNK等启动细胞内信号传导过程[28]。本文发现在28个注释参与T和B细胞信号
通路的分子中存在与TCR(酪氨酸蛋白激酶Lck、Fyn、Zap-70和Tec)以及BCR(B细胞连接蛋白Blnk、脾酪氨酸激酶Syk和布鲁顿酪氨酸激酶Btk)邻近的上游信号分子,通过实时荧光定量PCR方法验证了它们在日本七鳃鳗鳃囊组织中的转录表达情况(图3)。这些上游信号分子的发现与验证,说明七鳃鳗各淋巴细胞亚群与高等脊椎动物相比除了跨膜信号转导途径不同以外,它们胞内部分的信号传导机制可能是保守的。另外,本文还发现Lck、Fyn和Zap70等几个T细胞信号转导关键激酶同源分子在免疫激发后显著上调,说明七鳃鳗鳃黏膜免疫系统在受到抗原刺激后主要启动的是类似T细胞的免疫应答信号转导途径。
VLR分子C末端具有富含脯氨酸和苏氨酸的茎(Stalk)区和一段疏水尾(Hydrophobic tail)。VLRA 和VLRC 可以通过糖基磷脂酰肌醇(Glycosyl- phosphatidyl-inositol, GPI) 修饰, 切掉疏水尾后锚定到类淋巴细胞膜表面[29]。因此,分布于类淋巴细胞外膜表面的VLR分子一定需要跨膜蛋白分子的配合才能向胞内传递信号。由于在无颌类脊椎动物中没有发现TCR和CD3分子构成的T细胞受体复合体及BCR和CD79构成的B细胞抗原受体复合物,因此无颌类脊椎动物VLR信号跨膜传导的协同作用分子尚不清楚。Suzuki等[30]在盲鳗(E. burgeri)类淋巴细胞中发现了无颌类成对抗原受体样分子—APARs(Agnathan paired receptors resembling Ag receptors)。APARs中的一种(APAR-B)胞内区存在免疫受体酪氨酸抑制基序(Immunoreceptor tyrosine based inhibitory motif, ITIM);还有一类含有ITAM基序,被命名为NICIR(novel ITAM-containing immunogloblinsuperfamily receptors)[31]。但是APARs与各种VLR分子跨膜信号传导的联系目前还没有实验证据,未来尚需要进行更深入的研究和探索。
[1] Holmgren J, Czerkinsky C. Mucosal immunity and vaccines. Nat Med, 2005, 11(4 Suppl): S45-S53.
[2] Lin SH, Davidson GA, Secombes CJ, Ellis AE. A morphological study of cells isolated from the perfused gill of dab and Atlantic salmon. J Fish Biol, 1998, 53(3): 560-568.
[3] Press CMcL, Evensen Ø. The morphology of the immune system in teleost fishes. Fish Shellfish Immunol, 1999,9(4): 309-318.
[4] Lin SH, Ellis AE, Davidson GA, Secombes CJ. Migratory,respiratory burst and mitogenic responses of leucocytes isolated from the gills of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol, 1999, 9(3): 211-226.
[5] Forey PL, Janvier P. Agnathans and the origin of jawed vertebrates. Nature, 1993, 361(6408): 129-134.
[6] Braun CB. The sensory biology of the living jawless fishes: a phylogenetic assessment. Brain Behav Evol,1996, 48(5): 262-276.
[7] Baker ME, Funder JW, Kattoula SR. Evolution of hormone selectivity in glucocorticoid and mineralocorticoid receptors. J Steroid Biochem Mol Biol, 2013, 137: 57-70.
[8] Cooper MD, Alder MN. The evolution of adaptive immune systems. Cell, 2006, 124(4): 815-822.
[9] Pancer Z, Amemiya CT, Ehrhardt GRA, Ceitlin J, Gartland GL, Cooper MD. Somatic diversification of variable lymphocyte receptors in the agnathan sea lamprey. Nature,2004, 430(6996): 174-180.
[10] Pancer Z, Saha NR, Kasamatsu J, Suzuki T, Amemiya CT,Kasahara M, Cooper MD. Variable lymphocyte receptors in hagfish. Proc Natl Acad Sci USA, 2005, 102(26):9224-9229.
[11] Kasamatsu J, Sutoh Y, Fugo K, Otsuka N, Iwabuchi K,Kasahara M. Identification of a third variable lymphocyte receptor in the lamprey. Proc Natl Acad Sci USA, 2010,107(32): 14304-14308.
[12] Guo P, Hirano M, Herrin BR, Li JX, Yu CL, Sadlonova A,Cooper MD. Dual nature of the adaptive immune system in lampreys. Nature, 2009, 459(7248): 796-801.
[13] 毛芝娟, 楼丹, 杨波, 吴建军, 吴雄飞, 川合研儿, 陈昌福. 用灭活哈维弧菌菌苗浸泡免疫大黄鱼的研究. 华中农业大学学报, 2004, 23(3): 326-330.
[14] Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, Adiconis X, Fan L, Raychowdhury R, Zeng QD, Chen ZH, Mauceli E, Hacohen N, Gnirke A, Rhind N,di Palma F, Birren BW, Nusbaum C, Lindblad-Toh K,Friedman N, Regev A. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat Biotechnol, 2011, 29(7): 644-652.
[15] Finn RD, Bateman A, Clements J, Coggill P, Eberhardt RY,Eddy SR, Heger A, Hetherington K, Holm L, Mistry J,Sonnhammer ELL, Tate J, Punta M. Pfam: the protein families database. Nucl Acids Res, 2014, 42(D1):D222-D2230.
[16] Götz S, García-Gómez JM, Terol J, Williams TD, Nagaraj SH, Nueda MJ, Robles M, Talón M, Dopazo J, Conesa A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucl Acids Res, 2008, 36(10):3420-3435.
[17] Moriya Y, Itoh M, Okuda S, Yoshizawa A, Kanehisa M. KAAS: an automatic genome annotation and pathway reconstruction server. Nucl Acids Res, 2007, 35(Suppl. 2):W182-W185.
[18] Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol, 2009, 10(3): R25.
[19] Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform, 2011, 12: 323-339.
[20] Trapnell C, Williams B A, Pertea G, Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ, Pachter L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotech, 2010, 28(5): 511-515.
[21] Robinson MD, Oshlack A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol, 2010, 11(3): R25.
[22] Wang LK, Feng ZX, Wang X, Wang XW, Zhang XG. DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics, 2010,26(1): 136-138.
[23] Storey JD, Tibshirani R. Statistical significance for genomewide studies. Proc Natl Acad Sci USA, 2003,100(16): 9440-9445.
[24] Laird DJ, De Tomaso AW, Cooper MD, Weissman IL. 50 million years of chordate evolution: seeking the origins of adaptive immunity. Proc Natl Acad Sci USA, 2000, 97(13):6924-6926.
[25] Pancer Z, Cooper MD. The evolution of adaptive immunity. Annu Rev Immunol, 2006, 24: 497-518.
[26] Call ME, Pyrdol J, Wiedmann M, Wucherpfennig KW. The organizing principle in the formation of the T cell receptor-CD3 complex. Cell, 2002, 111(7): 967-979.
[27] Hirano M, Guo P, McCurley N, Schorpp M, Das S, Boehm T, Cooper MD. Evolutionary implications of a third lymphocyte lineage in lampreys. Nature, 2013, 501(7467):435-438.
[28] Seda V, Mraz M. B-cell receptor signalling and its crosstalk with other pathways in normal and malignant cells. Eur J Haematol, 2015, 94(3): 193-205.
[29] Herrin BR, Alder MN, Roux KH, Sina C, Ehrhardt GRA,Boydston JA, Turnbough CL Jr, Cooper MD. Structure and specificity of lamprey monoclonal antibodies. Proc Natl Acad Sci USA, 2008, 105(6): 2040-2045.
[30] Suzuki T, Shin-I T, Fujiyama A, Kohara Y, Kasahara M. Hagfish leukocytes express a paired receptor family with a variable domain resembling those of antigen receptors. J Immunol, 2005, 174(5): 2885-2891.
[31] Haruta C, Suzuki T, Kasahara M. Variable domains in hagfish: NICIR is a polymorphic multigene family expressed preferentially in leukocytes and is related to lamprey TCR-like. Immunogenetics, 2006, 58(2-3):216-225.
(责任编委: 胡松年)
Genetic basis of immune response of lymphocyte-like cells in the mucosal immune system of Lampetra japonica
Xin Liu1,2, Xueying Song1,2, Xiaoping Zhang1,2, Yinglun Han1,2, Ting Zhu1,2, Rong Xiao1,2,Qingwei Li1,2
1. School of Life Science, Liaoning Normal University, Dalian 116081, China;
2. Lamprey Research Center, Liaoning Normal University, Dalian 116081, China
In recent years, the antigen recognition mechanism based on variable lymphocyte receptors (VLRs) was found in agnathan lamprey. To illuminate the genetic basis of immune response of lymphocyte-like cells in the mucosal immune system of lamprey and explore the evolutionary relationship of adaptive immune responses between thejawless and jawed vertebrates, we constructed cDNA libraries of lamprey (Lampetra japonica) gills before and after stimulation, and then performed high-throughput transcriptome sequencing and analysis. Through functional annotation of 88 525 assembled unigenes, 21 704 and 9769 unigenes were annotated in Gene Ontology (GO) and Kyto Encyclopedia of Genes and Genomes (KEGG) databases, respectively. Among 999 unigenes involved in multiple pathways of immune system, 184 unigenes were highly homologous to 51 TCR (T cell receptor) and BCR (B cell receptor)signalling molecules in higher vertebrates, indicating that molecules involved in adaptive immune signalling pathways in higher vertebrates also exist in lampreys. In addition, identification of five VLRA, seven VLRB and four VLRC molecules suggest that at least three types of lymphocyte subsets are distributed in lamprey gill mucosal immune tissues. The results of real-time fluorescence quantitative PCR showed that the expression levels of Lck, Fyn and Zap70 were up-regulated after immune stimulation while those of Syk, Btk and Blnk were not changed significantly, indicating the activation of TCR-like signal transduction pathway after antigen stimulation in lamprey gill tissues. Our studies preliminaryly proved that two parallel adaptive immune systems in jawless and jawed vertebrates have common genetic basis, and also provided valuable clues to the exploration of signalling processes of VLRA+,VLRB+, and VLRC+lymphocyte-like cells in response to antigens.
Lampetra japonica; mucosal immune tissues; transcriptome;adaptive immune response
�到的Unigene序列作为参考序列(
equence),利用软件Bowtie[18]将每个样品的纯净读数比对(Mapping)到参考序列上,运行参数设为默认值。利用RSEM(RNA-Seq by Expectation Maximization)软件[19]对Bowtie的比对结果进行统计,进一步得到了每个样品比对到每个Unigene上的读数数量(Read count),并对其进行FPKM转换,估算基因的表达水平[20]。基因差异表达的输入数据为基因表达水平分析中得到的读数数量数据。采用TMM(Trimmed mean of M values)[21]对读数数量数据进行标准化处理,利用DEGseq[22]进行差异分析,筛选阈值为q<0.005且|log2(Sample1/Sample2)|>1[23]。
2015-03-16;
2015-05-19
国家重点基础研究发展计划(973计划)课题(编号:013CB835304)和国家自然科学基金项目(编号:31271323)资助
刘欣,教授,硕士生导师,研究方向:细胞生物学。E-mail:liuxin@lnnu.edu.cn
李庆伟,教授,博士生导师,研究方向:细胞生物学。E-mail:liqw@263.net
10.16288/j.yczz.15-108
网络出版时间: 2015-8-289:25:48
URL: http://www.cnki.net/kcms/detail/11.1913.R.20150828.0925.002.html
