不同林龄橡胶凋落物叶分解特性与有机碳动态研究
2015-10-21金龙吴志祥杨川管利民赖华英
金龙 吴志祥 杨川 管利民 赖华英

摘 要 通过网袋法,研究一年内6 a、12 a、20 a、28 a共4组不同林龄橡胶凋落物叶分解及有机碳动态变化的过程。结果表明:(1)不同林龄橡胶凋落物叶均表现为慢-快-慢的分解趋势,经1 a的分解,4组林龄组橡胶凋落叶残留率分别为28.33%、30.53%、31.59%和33.89%。(2)不同林龄橡胶凋落物叶分解系数为1.376 2~1.608 8,平均值为1.498 2,大小顺序为6 a>12 a>18 a>28 a,分解50%和95%所需时间分别为0.43、0.45、0.47、0.50 a和1.86、1.96、2.03、2.18 a。(3)林龄显著影响橡胶凋落物叶初始N含量、P含量、木质素/N比、C/P比与木质素/P比,但对C含量、木质素含量及C/N比影响不明显。(4)初始C/N比极显著影响橡胶凋落物叶分解速率,而初始P含量对凋落物分解影响不显著(5)4组林龄组橡胶凋落物叶有机碳浓度是一个先上升后下降的过程,研究结束时,4组林龄橡胶凋落物叶有机碳分别释放了77.58%、79.41%、75.89%和75.02%,其动态变化过程显著符合一元多项式模型,决定系数(R2)在0.88以上。
关键词 橡胶林;不同林龄;凋落物;分解速率;有机碳
中图分类号 S718 文献标识码 A
森林凋落物是森林生态系统的重要组成部分,同时也是地表及地下分解者的主要食物来源,其生产和分解过程是森林生态系统物质循环和能量流动的重要环节之一[1]。另外,森林凋落物还在改善土壤理化性质、提升林地生产力、减少水土流失及保温增湿等方面起着重要作用[2-4]。从广义上讲,森林凋落物主要包括枯叶、枯枝、死亡根系和倒木等部分[5],其中叶是主要成分,约占凋落物总量的60%~80%,甚至更高[1-6]。森林凋落物及其分解一直都是土壤学、生态学、环境科学及林学等学科的研究重点,至今已有100多年的历史,自德国生物学家Ebermayer于1876年首次报道森林凋落物及其在养分循环中的作用以来,国外就陆续开展了生态系统内凋落物的总量、组成成分、凋落节律、凋落物分解与养分释放及分解模型等方面的研究[7-9]。而中国有关凋落物研究的时间较短,直到20世纪80年代,中国学者才开始对凋落物的分解进行研究,主要涉及生物因素和非生物因素对凋落物分解的影响、养分动态变化研究[10-11]。20世纪90年代后,伴随对全球气候变化的广泛关注,有关凋落物分解的研究被结合在了对全球碳收支平衡的调节控制上[12-13]。近10年来,随地下生态学研究不断深入及地下生态过程在响应全球变暖过程中的重要性,凋落物对地下生态过程调控机制也越来越受到关注[13]。但由于影响凋落物分解的因素复杂众多,凋落物中各种化学组分及其相互作用对凋落物分解的影响目前仍不十分不清楚[6, 13],尤其是在气候变化的背景下,C、N养分及其交互作用对凋落物分解的调控机制还需更深入的研究[13]。因而,尽管已经有大量森林凋落物研究报道的资料[14-15],但全球仍在持续深入研究各类型森林凋落物的特征及其分解对气候的响应机制。
橡胶林生态系统是中国热区一种最典型的人工林生态系统,从第九届中国橡胶基础研究研讨会上获悉,截至2012年,中国植胶面积达到了111万hm2。在全球广泛开展森林凋落物研究背景下,中国关于橡胶林凋落物的研究也取得了一定的进展。杨曾奖等[16]研究了27 a橡胶林间作砂仁模式下凋落物的组成及分解状况;任泳红等[17]研究了橡胶多层林凋落物的节律及分解变化;曹建华[18]报道了PR107品种橡胶3.0 m×7.0 m 栽培模式下9个年龄段(4~28 a)凋落物量及养分的释放状况;赵春梅等[19]研究了不同林龄橡胶枯落物的分解及氮素释放过程等。Philip等[20]研究了15 a橡胶枯落物的分解与养分的释放状况。但对橡胶这类人工林凋落物的研究范围仍较窄,主要集中在橡胶林凋落物量及养分释放动态的研究,而有关养分的研究也主要集中在N、P、K、Ca、Mg等营养元素的释放规律上,而对橡胶凋落叶基质质量与其分解速率的关系,分解过程中有机碳动态及其释放量的研究还鲜有报道。基于此,本研究通过野外分解袋法研究了不同林龄(6 a、12 a、20 a 和28 a)橡胶凋落叶分解过程及其有机碳动态变化的过程。旨在为准确评估橡胶林碳源/汇功能提供参考依据,同时也为林地养分管理、提高林地生产力等方面提供了理论依据。
1 材料与方法
1.1 试验地概况
不同林龄橡胶凋落物叶分解实验的试验样地设在海南儋州中国热带农业科学院试验农场3队内,地点位于19°31′47"N,109°29′30"E,平均海拔144 m,属于热带季风气候,年平均气温为20.5~28.5 ℃,最冷月平均气温为16.5~17.6 ℃,全年日平均气温≥10 ℃的积温为8 500~9 100℃;全年旱雨两季分明,5~10月为雨季,11月~翌年4月为旱季,年平均降雨量为1 607~2 000 mm,其中7、8、9共3个月降雨量占全年降雨量的70%以上;年平均相对湿度为83%;实验区地形为缓坡丘陵(相对高度差<10 m),土壤为花岗岩母质化所形成的砖红壤,土层厚度约为100 cm,pH值4.52~5.86。
该实验区为第二代胶园,其中,分解实验样地林下隔行间作有用于压青肥的天堂鸟,其他植被有从当年生草本到多年生小草本,包括弓果黍(Cyrtococcum patens)、飞机草(Chromolaene odorata)、地胆头(Elephantopus scaber)、 含羞草(Mimosa pudica)等。
1.2 凋落物分解实验
凋落物分解采用网袋法,同时为保证土壤性质和气候环境的均匀一致性,本研究以试验场内6 a、12 a、20 a 和28 a共4个林段(栽培品系分别为RR525、热研7-33-97、热研7-33-97和PR107)橡胶树的新鲜枯落叶作为实验材料,于2013年12月橡胶树开始落叶后,分别收集4个不同年龄段新鲜凋落物叶,待样品风干后,准确称取(50.00±0.50)g直接裝入大小为45 mm×35 mm的网袋(孔径大小为1.2 mm×1.2 mm)内,于2014年1月10日分别随机放入6~12林段(林龄为12 a,3 m×7 m常规种植模式)的地表,每林龄15袋,3个重复,共计60袋,并用枯落物将其覆盖。2014年3月10日开始取第一次样,然后每隔2个月取一次样,共计5次,每次每个林龄各取回3袋,轻轻除去分解袋上附着的泥沙,并去除袋内根系等杂物,烘干至恒重后称重,计算凋落物的分解速率;同时实验室采用重铬酸钾法测定样品的有机碳含量[21],采用靛酚蓝比色法测定全N[22],采用钼锑抗比色法测定全P[22],采用溴乙酰-乙酸比色法测定木质素[23]。
1.3 质量残留率与有机碳释放速率的测定
凋落物分解过程中凋落物残留率和有机碳分释放速率采分别采用以下公式进行计算:
枯落物残留率:Di=(Wi/W0)×100%,式中,Di为第i月的残留率(%),Wi为第i月所取样品的剩余重,W0为落叶初始质量。
有机碳释放速率=[(初始浓度×初始质量)-(t时刻浓度
×t时刻质量)]/(初始浓度×初始质量)×100%。
1.4 基于哑变量的Olson指数衰减模型
目前研究凋落物分解的模型有多种,其中以Olson指数衰减模型应用最为广泛,形式为:
Mt=M0e-kt,式中,M0是初始质量,Mt为某一时间t时的质量,k为分解速率常数。
目前针对不同林龄生长模型的预测时都是单独建立模型,这样建立的模型会造成整体和分量之间的不相容,增大了模型误差。而采用哑变量(dummy variable)可把各林龄用定性代码来表示,从而整合成一个模型来构建,这样既减少了工作量又提高了模型精度。哑变量的定义为: 对于等级性(定性)数据x,用变量δ(x,i)表示成:
这种方法叫做定性因子(0,1)化展开,因此称变量δ(x,i)为哑变量。一个定性变量(m个等级)对应一个向量δ(x,#)=[δ(x,1),…,δ(x,m)]。哑变量只取0或1,于是一个定性变量就变成取0或1的数值向量,便可用数值方法处理[24-25]。
本研究所收集的数据来自4个林龄,第i个林龄编号为Si,把定性数据Si转化为(0,1)数据:
则可将Olson指数衰减模型改写成哑变量形式如下:
y=(a1S1+a2S2+a3S3+a4S4)e[-(k1S1+k2S2+k3S3+k4S4)t]
式中,a1、a2、a3、a4是待定参数,S1、S2、S3和S4分别是6 a、12 a、20 a和28 a共4个林龄的代码,k1、k2、k3和k4分别为4个林龄的分解系数,t为分解时间。
1.5 数据分析
采用Microsoft office excel 2007、origin9.0等软件对所有取样点数据进行整理、分析、计算、作图。
2 结果与分析
2.1 不同林龄橡胶凋落物叶初始质量参数
凋落物叶化学组分是影响其分解的重要内在因子,不同林龄橡胶林凋落物叶初始质量参数见表1。不同林龄(未考虑品系因素,另有说明,下同)橡胶凋落叶有机碳含量介于49.29%~51.86%,平均含量为51.58%。橡胶凋落物叶中N、P浓度随林龄的增长而呈波动变化,其中N浓度表现为28 a、12 a显著低于6 a和20 a;P浓度表现为6 a、28 a显著低于12 a和20 a。木质素含量、C/N比、木质素/N比和木质素/P比随林龄变化均呈升高-降低趋势,C/P比呈降低-升高趋势,平均值分别为23.69%、24.52、10.97和154.62;方差分析结果表明,木质素浓度、C/N比在不同林龄间差异不显著;木质素/N比为6 a显著低于12 a、20 a、28 a;木质素/P比为6 a、12 a 显著低于20 a、28 a;C/P比为6 a、28 a显著低于12 a和20 a。综上所述,橡胶凋落物叶的品质随林龄的增加有所降低。
2.2 凋落物叶分解残留率的月变化
通过分解袋法,计算样品的残留率来研究橡胶凋落叶的分解速度。由表2可知,4组不同林龄橡胶凋落物叶的残留量随时间的推移不断降低,月质量损失率存在明显的季节变化,表现为先升高后降低的趋势,分解高峰出现在3~7月,分解高峰平均月分解速率为12.16%。经过将近1 a的分解,4组林龄橡胶凋落物叶最终残留率随林龄的增加表现出升高趋势,分别为28.33%、30.53%、31.59%和33.89%,但不同林龄间分解速率差异不显著(p>0.05)。说明林龄对橡胶凋落物叶分解速率有一定的影响,随林龄的增加分解难度增加。
2.3 含有哑变量Olson指数衰减模型拟合
本研究利用含有哑变量的Olson指数衰减模型进行拟合,拟合结果为:
Y=(93.469S1+95.348S2+95.014S3+92.042S4)e[-(1.608 8S1+1.532 3S2+1.475 4S3+1.376 2S4)t]
式中S1、S2、S3和S4分别是6 a、12 a、20 a和28 a共4个林龄的代码;t为分解时间。6 a、12 a、20 a和28 a共4個林龄的拟合参数为93.469、95.348、95.014、92.042;分解系数(K)分别为1.608 8、1.532 3、1.475 4和1.376 2。4个林龄的拟合结果均达到极显著水平(p<0.01),决定系数在95%以上,拟合效果较好。比较不同林龄橡胶凋落物叶的分解系数,不同林龄之间K值的大小顺序为6 a>12 a>20 a>28 a(表3)。根据该模型计算分解50%、95%所需的时间,结果显示4林龄分解50%所需时间分别为0.43、0.45、0.47、0.50 a,分解95%所需时间为1.86、1.96、2.03、2.18 a,该值要略大于任泳红等[17]和曹建华等[18]所报道的1.33 a和12个月(表3)。
2.4 凋落物叶初始质量与分解系数的相关性
在相同分解条件下,凋落物分解速率主要受凋落物化学成分的影响,且不同分解阶段影响的因子不同,前期主要受养分含量、水溶性碳化合物和结构碳化合物含量的强烈影响,而后期则更多地受木质素及纤维素/木质素比的支配[26]。通过分析不同林龄橡胶凋落物叶分解系数与各主要化学成分之间的关系(表4),分解系数与初始C/N比呈极显著负相关,与C、N、C/P比含量呈正相关,与木质素、P、木质素/N比、木质素/P比呈负相关,其中与木质素、木质素/N的相关性在0.7以上。可见,在同一气候环境下,C/N比是引起不同林龄橡胶凋落物叶分解速率差异的最为主要原因。
2.5 凋落物叶分解过程中有机碳浓度的变化
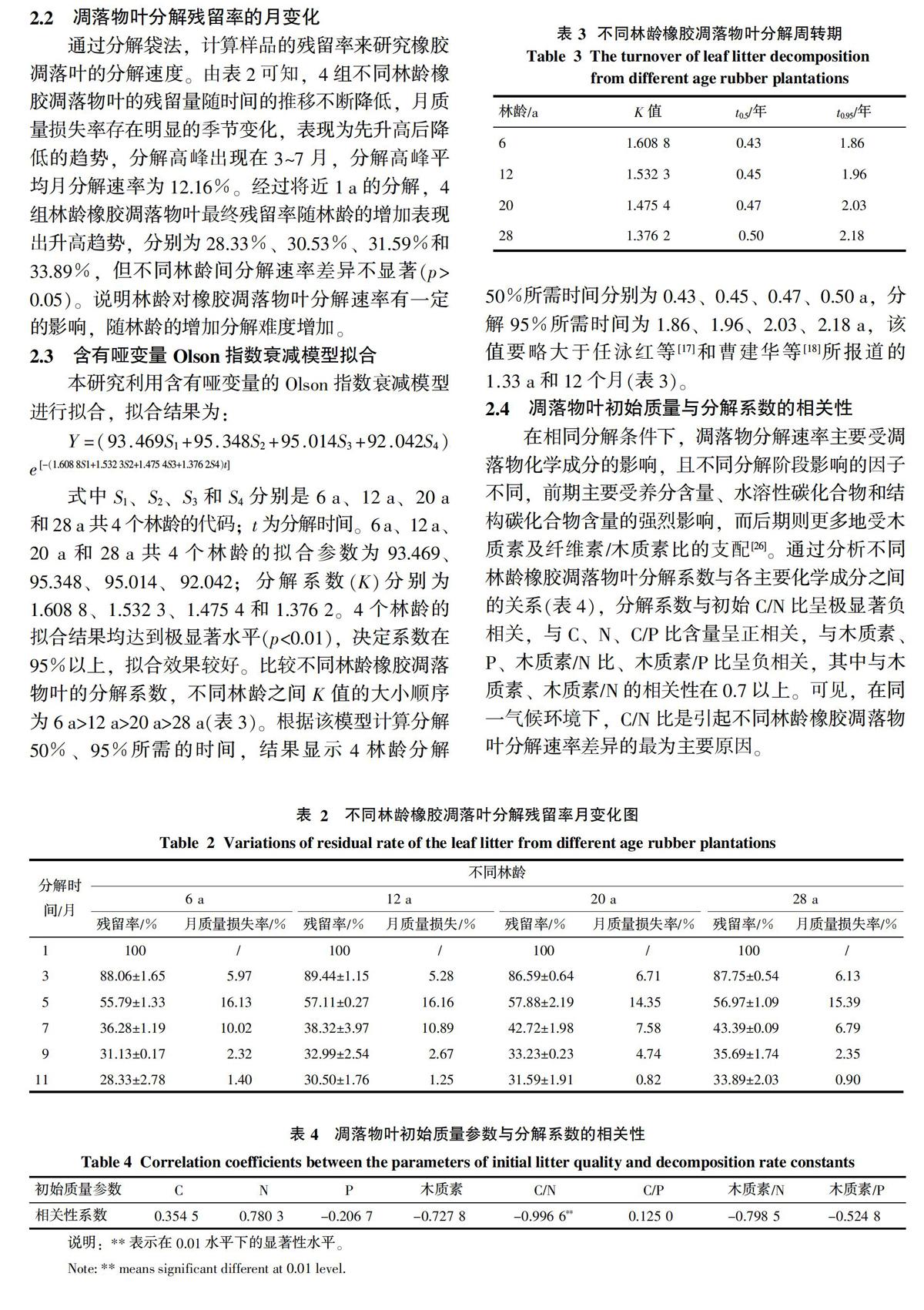
由图1可知,橡胶凋落物叶分解过程中,有机碳浓度表现出一定的变化趋势:4组不同林龄橡胶凋落叶有机碳浓度在分解初期(1~3月)有一个上升的过程,且上升幅度在不同林龄间存在差异,其中,上升幅度以12 a最高,以20 a最低,相对初始浓度分别增加了10.46%和4.81%,6 a和28 a则分别增加了7.27%和6.18%;分解开始后的3~7月,4组林龄橡胶凋落叶有机碳浓度均表现为明显的快速下降过程;7月后,除12 a在9~11月有一个突降外,其余3林龄组有机碳浓度趋于平稳,取样结束时,4组林龄橡胶凋落物叶有机碳浓度分别为39.66%、35.01%、38.24%和36.33%,较初始浓度分别下降了10.46%、16.85%、11.87%和12.96%。
2.6 凋落物叶分解过程中有机碳释放速率的动态变化
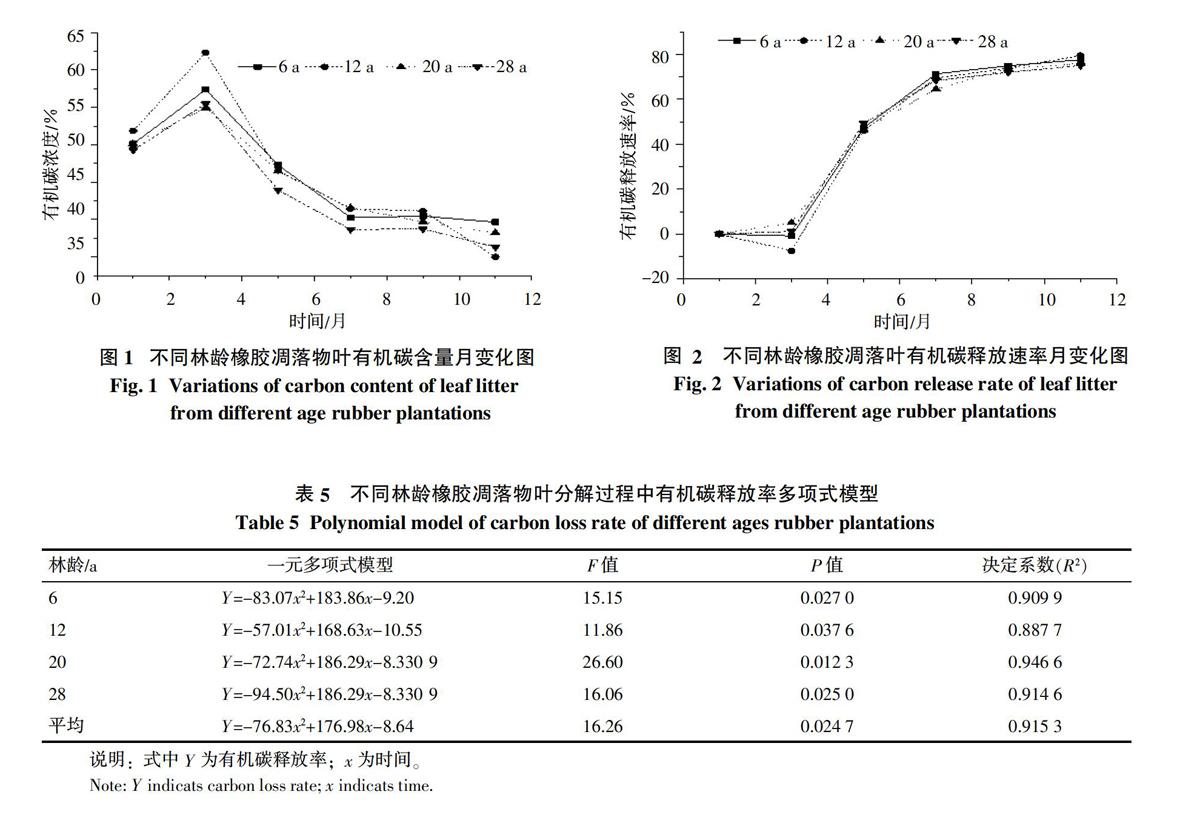
由图2可知,4组不同林龄橡胶凋落物叶有机碳释放速率呈现以下特征:在分解初期(1~3月),有机碳释放速率慢,甚至表现为净积累过程,如6 a和12 a橡胶凋落叶,净积累量分别为0.84%和7.48%;分解开始后的3~7月,为有机碳快速释放阶段;7月后,释放速率开始放缓,整个过程中,4林龄橡胶凋落物叶有机碳释放速率较为一致;至11月最后一次取样时,总有机碳量分别释放了77.58%、79.41%、75.89%和75.02%。
本研究运用6种不同模型对各林龄橡胶凋落物叶分解过程中有机碳释放速率随时间变化过程进行拟合,结果以一元多项式拟合方程最为理想(表5)。由表5可知,4组不同林龄橡胶凋落物叶在分解过程中,有机碳释放速率随时间的变化均显著(p<0.05)符合一元多项式模型,决定系数在0.88以上。因此,用一元多项式方程对橡胶凋落叶有机碳释放速率进行模拟可取得较好的效果。
3 讨论与结论
3.1 不同林龄橡胶凋落物叶初始养分分析
N、P是植物体的重要组成部分,参与生命物质的合成。随林龄的增加,橡胶凋落叶初始N含量、P含量均呈现出显著性波动变化,其中N表现为6 a>20 a>12 a>28 a,P表现为12 a>20 a>28 a>6 a,总体养分及化学组分别随林龄的增加有所降低。N、P等养分在不同林龄橡胶间的波动变化,不仅与土壤养分供应有关,还与养分转移及在各器官之间的分配有关[27]。橡胶树在幼龄期(6 a)由于生物量小,对N、P等养分总需求量低,且主要用于生物量的增长,因而单位体积土壤能提供相对充足的养分;进入开割期后(12 a、 20 a),虽然有一部分养分伴随胶乳流失,但这种刺激可能会激发对养分的吸收,同时由于凋落物量也基本进入稳定期,再加上人为施肥等措施能够及时补充橡胶正常生长所需。对于老龄(28 a)即将更新树,地上生物量大、人工扶管措施减弱,前期土壤肥力流失严重,因此导致对N、P等养分吸收竞争激烈,难度增大。植物体的元素化学组分主要来自于根系对土壤养分的吸收,因此,土壤质量的不同必然引起橡胶生长状况的差异,而这种差异可能首先就反映在植物中心部位-叶上。另外,元素转移在植物体内普遍存在,橡胶叶在掉落前,养分在不同林龄橡胶植物体内的转移差异,也会影响到凋落叶的养分含量。而对于P为何6 a显著性最低,是因为其余3林龄均为割胶林段,而割胶刺激可能激发了橡胶树对P的吸收。研究结果还表明,林龄间的N、P养分显著性差异还导致了木质素/N、木质素/P的显著性差异。
3.2 不同林龄橡胶凋落物叶分解速率分析
4组林龄橡胶凋落物叶随时间变化均呈慢-快-慢的分解过程,周转时间随林龄增大逐渐增长。在1 a的分解期内,橡胶凋落物叶分解速率是雨季分解快,旱季分解慢,前期分解快,后期分解慢,分解高峰出现在3~7月,这与前人研究的结论基本一致[19]。分解前期(1~2月)为一年中最冷月份,温度低,降雨量少,微生物活性低,因淋溶作用和微生物代谢作用引起的凋落物质量损失少;夏季(雨季)高温多雨,生物代谢和淋溶作用增强,因而凋落物分解速度快速增大;分解后期(7~11月),随易分解物质的消耗殆尽,难分解物质大量积累,微生物可食用物质减少,凋落物叶上的生物开始出逃或则死亡,分解速度也开始减慢。在同一分解环境下,橡胶凋落叶品质是影响其分解速度的一个最为重要的因素。橡胶凋落叶品质随林龄增加总体有所降低,其中,作为预测凋落物分解速度的C/N比[26],随林龄的增大逐渐增大。因此,不同林龄橡胶凋落叶品质差异是导致其分解快慢最为重要的因素。
经含有哑变量Olson指数衰减模型预测,4组林龄橡胶凋落叶彻底分解所需时间在1.86 a以上,高于任泳红等[17]和赵春梅等[19]同类报道的1.33 a和12个月。这可能与林下微气候环境、土壤生物及土壤肥力不同有关。本实验林下由于间作用于胶园压青的天堂鸟,其宽大密集的叶片对地表水热通量起到一定的拦截,相比较凋落物质量,温度和降水在凋落物分解中起主导作用[8],因此,水、热等环境因子的微小变化,必将对凋落物的分解产生重大影响。另外,地表间作天堂鸟可能会引起地表生物数量、群落结构及生物活性的改变,这也是影响凋落物分解速率差異的一个重要原因。
3.3 橡胶凋落物叶质量与分解速率的关系
橡胶凋落物叶初始C/N比极显著影响分解速率,而初始P含量及C/P比对分解速率影响不显著,均表现为弱相关性。已有研究结果发现,凋落物分解速率随初始N、P浓度增加而增加,随初始木质素浓度、C/N比和木质素/N比的增大而减小[26]。还有研究结果表明,受氮沉降的影响,P在热带地区凋落物分解中的重要性要大于N[28]。本研究结果发现,N、C/N比对橡胶凋落叶分解的重要性要强于P及C/P比,随C/N比增加,分解速度逐渐降低,且C/N比与分解系数呈极显著相关,可用来作为预测橡胶凋落物叶分解的指标。至于P及C/P比为何在橡胶凋落叶分解中没有表现出相当的重要性,甚至P表现为弱相关性。分析原因,可能是因为橡胶凋落物叶初始C/P比值(318.84~380.22)远低于大多数凋落物分解的临界C/P比值(1 000~1 300),而凋落物分解的临界C/P比值就是分解微生物的临界C/P比值[26],因此,橡胶凋落物叶中较为丰富的P可能不是微生物分解的限制因子。另外,P的行为还与凋落物类型有关[29]。唐仕姍等[30]总结了中国不同森林类型凋落物研究成果后发现,中国主要森林类型的凋落物初始P浓度与分解系数的关系不显著,其中,初始P浓度介于0.1~2.7 g/kg,且主要分布在小于1 g/kg的范围内。本研究结果还发现,碳组分/养分与分解系数的相关性优于单独养分,相关性绝对值大小前4位为C/N>木质素/N>木质素>木质素/P,说明凋落物碳组分与N、P等养分的相互作用在调控橡胶凋落物叶分解过程中起着关键性作用。
3.4 橡胶凋落叶有机碳动态分析
4组林龄橡胶凋落物叶有机碳相对含量总体呈先上升后下降,最后趋于平稳的态势,其变化过程与凋落物的分解速率基本一致。有机碳相对含量是指有机碳绝对量与其干物质量的比值,有机碳浓度的上升下降过程,不仅体现了有机碳绝对量的释放过程,同时也反映出了干物质的损失过程。凋落物分解前期主要为可溶性物质的淋溶损失过程,后期则为有机物的代谢损失过程[26]。前期分解过程中,由于作为有机碳主要成分的纤维素、木质素还未分解,有机碳损失量小于干物质损失量,因此有机碳浓度表现为上升过程;随着纤维素、半纤维素等有机物大量分解,有机碳浓度也开始快速降低;最后残留下的木质素等难分解的物质,限制了微生物的利用,导致干物质损失速率开始降低,有机碳浓度变化也基本趋于平稳。同时,通过对比前人的研究结果,发现橡胶凋落叶有机碳动态变化过程与李正才等[31]报道的北亚热带6种人工林有机碳动态变化比较一致,与刘强等[32]报道的变化趋势有所不同。另外,由于热带的水热条件优于亚热带,橡胶凋落叶分解1 a后的有机碳浓度变化量高于亚热带人工林树种。同时,由于凋落物类型、分解环境及参与分解的生物类群、数量及活性差异,其释放量低于尖峰岭热带雨林等样地。
3.5 橡胶凋落物叶有机碳释放模型分析
4组林龄橡胶凋落物叶有机碳释放速率显著符合一元多项式模型,与凋落物分解指数衰减模型不同。橡胶凋落物叶有机碳释放速率模型与干物质损失模型不同,说明有机碳释放过程与凋落物分解过程不完全相同,这是因为对每种凋落物而言, 凋落物有机碳的分解与凋落物干物质降解的过程不同[31];但对于橡胶凋落叶这一特定类型的凋落物,其有机碳分解快慢的顺序与其干物质分解快慢的顺序总体上较为一致, 说明橡胶凋落物有机碳的释放和其干物质的分解机制存在一定的关系。和同类报道相比,与李正才等[31]、陈莉莎等[33]报道的有机碳指数释放模型也不同,这可能是因为区域气候环境及凋落物种类差异,导致凋落物分解速率及有机碳释放过程中的调控机制不同有关。
本研究的不足之处主要是在分析林龄对橡胶凋落物叶分解及养分影响时,未考虑橡胶品系间的差异可能引起分解速率及化学组分含量的变化。但根据前人的研究结果,相同林龄不同品系的橡胶枯落物分解速率一致,养分含量也没有显著性差异变化[34-35];相反,同一品系不同林龄橡胶枯枝、枯叶及鲜枝、鲜叶养分含量存在显著差异[18],因此,可推定不同林龄橡胶凋落物叶品质及分解速率差异主要由林龄效应引起。
参考文献
[1] 于恩娜, 王金贵, 初宝顺. 凋落物及其在森林生态中的作用[J]. 现代农业科技, 2009, 23(2): 286-288.
[2] 薛 立, 何跃君, 屈 明,等. 华南典型人工林凋落物的持水特性[J]. 植物生态学报, 2005, 29(3): 415-421.
[3] 肖 洋, 陈丽华,余新晓. 北京密云麻栎人工混交林凋落物养分归还特征[J]. 东北林业大学学报, 2010, 38(7): 13-15.
[4] 杨丽萍,苏海鹏. 橡胶林凋落物研究进展[J]. 热带农业工程,2011, 35(4): 40-43.
[5] 赵鹏武, 宋彩玲, 苏日娜,等. 森林生态系统凋落物研究综述[J]. 内蒙古农业大学学报(自然科学版), 2009, 30(2): 292-299.
[6] 宋曰欽, 翟明普,贾黎明. 三倍体毛白杨地上凋落物对年龄的响应[J]. 东北林业大学学报, 2010, 38(3): 17-19, 22.
[7] Bray J R, Gorham E. Litter production in forests of the word[J]. Advances in Ecological Research, 1964, 2(1): 101-157.
[8] Aerts R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems:a triangular relationship[J]. Oikos, 1997, 79(3): 439-449.
[9] 林成芳, 高 人, 陈光水,等. 凋落物分解模型研究进展[J]. 福建林业科技, 2007, 34(3): 227-233.
[10] 王 斌, 杨校生. 不同气候区4种典型地带性植被凋落物比较研究[J]. 世界林业研究, 2009, 29(S): 44-48.
[11] 贾开心, 郑 征,张一平. 西双版纳橡胶林生物量随海拔梯度的变化[J]. 生态学杂志, 2006, 25(9): 1 028-1 032.
[12] 彭少麟,刘 强. 森林凋落物动态及其对全球变暖的响应[J]. 生态学报, 2002, 22(9): 1 534-1 544.
[13] 宋 飘, 张乃莉, 马克平,等. 全球气候变暖对凋落物分解的影响[J]. 生态学报, 2014, 34(6): 1 327-1 339.
[14] Talbot J M, Lehtonen A. Needle and branch biomass turnover rates of Norway spruce(Picea abies)[J]. Canadian Journal of Forest Research, 2004, 34(12): 2 517-2 527.
[15] 徐李亚, 杨万勤, 李 晗,等. 高山森林林窗对凋落物分解过程中水溶性氮和磷的影响[J]. 水土保持学报, 2014, 28(3):214-221.
[16] 杨曾奖, 郑海水, 周再知,等. 橡胶间种砂仁模式下凋落物的特征[J]. 广东林业科技, 1997, 13(4): 19-24.
[17] 任泳红, 曹 敏, 唐建维,等. 西双版纳季节雨林与橡胶多层林凋落物动态的比较研究[J]. 植物生态学报, 1999, 23(5):418-425.
[18] 曹建华, 陶忠良, 赵春梅,等. 不同树龄橡胶树枯落物养分归还比较[J]. 热带作物学报, 2011, 32(1): 1-6.
[19] 赵春梅, 曹建华, 李晓波, 等, 橡胶林枯落物分解及其氮素释放规律研究[J]. 热带作物学报, 2012, 33(9): 1 535-1 539.
[20] Philip A, Philip V, George E S, et a1. Leaf litter decomposition and nutrient release in a fifteen year old rubber plantation[J]. Indian Journal of Natural Rubber Research, 2003, 16(1/2): 81-84.
[21] 中国科学院南京土壤研究所. 土壤理化分析[M]. 上海: 上海科学技术出版社, 1978.
[22] 鲍土旦. 土壤农化分析(第三版)[M]. 北京:中国农业出版社,2000: 30-34.
[23] 李 靖, 程 舟, 杨晓伶,等. 紫外分光光度法测定微量人参木质素的含量[J]. 中药材, 2006, 29(3): 239-241.
[24] 王 欣, 高明达, 杨 飞,等. 不同林龄华北落叶松人工林叶凋落物分解及养分动态比较[J]. 东北林业学报, 2012, 40(10): 55-60, 66.
[25] 李忠国, 孙晓梅, 陈东升,等. 基于哑变量的日本落叶松生长模型研究[J]. 西北农林科技大学学报:自然科学版, 2011, 39(8): 69-74.
[26] Xu X, Hirata E. Decomposition patterns of leaf litter of seven common canopy species in a subtropical forest: N and P dynamics [J]. Plant and Soil, 2005, 273(2): 279-289.
[27] 邹积鑫, 曹建华, 蒋菊生,等. 橡胶树无性系PR107不同树龄叶片养分含量比较研究[J]. 热带农业科学, 2009, 29(10): 9-12.
[28] Hastwell G. Nutrient cycling and limitation: hawaii as a model system[J]. Austral Ecology, 2005, 30(3): 609-610.
[29] Moro M J, Domingo F. Litter decomposition in four woody species in a mediterranean climate: weight loss, N and P dynamics[J]. Annals of Botany, 2000, 86(6): 1 065-1 071.
[30] 唐仕姍, 楊万勤, 殷 睿,等. 中国森林生态系统凋落叶分解速率的分布特征及其控制因子[J]. 植物生态学报, 2014, 38(6): 529-539.
[31] 李正才, 徐德应, 杨校生, 等. 北亚热带6种森林类型凋落物分解过程中有机碳动态变化[J]. 林业科学研究, 2008, 21(5):675-680.
[32] 刘 强, 彭少麟, 毕 华,等. 热带亚热带森林凋落物交互分解的养分动态[J]. 北京林业大学学报, 2005, 27(1): 24-32.
[33] 陈莉莎, 张金池, 庄家尧, 等. 苏南丘陵区3种林分凋落叶客置分解特征[J]. 浙江农林大学学报, 2013, 30(5): 681-688.
[34] 谢学方. PR107和热研7-33-97橡胶人工林胶园土壤养分平衡研究[D]. 海南: 海南大学, 2011.
[35] 王丽华,林钊沐. 不同产量水平橡胶品种幼树的氮磷钾营养差异研究[J]. 广东农业科学, 2009, 36(3): 31-35.
