不同饵料组合对马来沙水母生长的影响
2015-10-10陈华增齐继光王云忠杨翠华刘光兴
陈华增,齐继光,王云忠,杨翠华,刘光兴,王 玮
(1.青岛水族馆,山东 青岛 266003;2.中国海洋大学环境科学与工程学院,山东 青岛 266003)
马 来 沙 水 母Sanderia malayensis Goette(1886)又称天草水母,属刺胞动物门(Cnidaria)钵水母纲(Scyphomedusae)旗口水母目(Semaeostomeae)游水母科(Pelagiidae)沙水母属(Sanderia),主要分布于我国的东海、南海以及日本、马来西亚、新加坡和菲律宾等国家的海域,属暖水性种类[1]。马来沙水母伞体扁平,直径6~10 cm,具32个缘瓣和l6 条触手,外伞部具刺胞疣,口腕两侧缘有许多褶皱,4个间辐位赤褐色生殖腺呈马蹄形,其口腕舒展似丝带,轻柔飘逸,具有极佳的观赏性。
水族馆是集收集、研究、饲养和展示诸功能于一体的机构,其社会责任是通过呈现自然界的多样性和奇妙性,引导人们认识和保护自然环境。随着国内外水族行业发展日渐兴盛,至2009年7月,国内水族馆已近70 家,而日本水族馆在1994年时已有230 家之多,与其相比,我国的水族馆发展仍有较大空间[2]。
由于人类对海洋发展与保护认知的不断提高,马来沙水母的生物学价值、经济价值亦逐渐显现。可持续地展现马来沙水母价值的前提不仅靠自然捕捞,更应通过科学饲养,从而实现其长期性和稳定性。为此,国内外学者进行了大量的研究。例如:日本学者在20世纪70年代研究了马来沙水母的水螅体及其繁殖行为[3-4];Laetitia 等[5]研究了马来沙水母不同类型的无性繁殖模式,建立了钵水母无性繁殖分类方式的标准;郭嘉瑱等[6]观察了马来沙水母各个发育阶段的形态特征;郭玲玲等[7]探讨了光照强度、光质和光周期对马来沙水母幼体生长的影响。但关于马来沙水母食性的研究较少。马来沙水母的食性复杂,可摄食枝角类、桡足类、鱼虾幼体等浮游动物及水中有机颗粒等[8-9]。对人工养殖而言,由于受到饵料来源、成本等因素的影响,马来沙水母的饵料来源不多,难以像在自然环境下一样摄取到多种食物,这在一定程度上影响了其正常生长,进而限制该水母的室内大规模繁殖饲养。因此,研究通过比较不同饵料对马来沙水母生长性状的影响,以期为马来沙水母的养殖提供数据支持。
1 材料与方法
1.1 试验材料
试验用马来沙水母幼体为青岛水族馆人工培育所得,伞径为2.29±0.24 cm,体质健康、无损伤、游动力强。
试验缸为封闭式循环水系统,各试验组间相对独立。试验缸水体200 L,盐度32,水体温度20±0.5℃,使用150 W 金属卤素灯每天照明8 h。通过蛋白分离器、紫外杀菌灯和陶瓷滤环等设施控制试验缸的水质。通过调节系统分压阀来控制试验缸水流的大小,使水母始终悬浮在缸中自由游动。
1.2 试验方法
1.2.1 试验饵料的分组和投喂 整个试验分为6 组,分别投喂颗粒饲料(PD)、糠虾Mysinae(MS)、丰年虫Chirocephalus(FS)、海月水母Aurelia aurita +颗粒饲料(MP)、海月水母+糠虾(MM)和海月水母+丰年虫(MF);其中,MP、MM、MF 是指上午投喂海月水母块(活体海月切成伞径一半大小的块,用镊子投喂到马来沙水母口腕处),下午依照不同的试验组分别投喂颗粒饲料(MP 组)、糠虾(MM 组)和丰年虫(MF 组)于水母口腕处。每组设3个平行,每缸投放马来沙水母25 只。试验用颗粒饲料为某品牌海水鱼苗专用微颗粒饲料(粒径0.2~0.3 mm),糠虾(体长0.6~0.8 cm)和丰年虫(体长0.5~0.6 cm)为冰冻品,作饵料用的海月水母由青岛水族馆自主培育所得。各试验组每天投喂两次,时间分别为8︰30、15︰30。投喂量以水母胃囊呈饱满状态为准,根据水母的生长变化,调整每天的投喂量。试验时间(T)为30 d,每5 d 测量一次生长指标。
1.2.2 指标测定 试验测定的马来沙水母生长指标有口腕长(OL)、伞径(DM)和体重(W)。口腕长为水母口腕的纵向长度,体重定义为从试验缸取出的马来沙水母沥水后的湿重。每组取3 只,其口腕长和伞径用游标卡尺测量,体重用电子称(0.1 g)测得。根据测得的试验起始和最终生长指标数据,计算各试验组马来沙水母的特定生长率(SGR)、伞径与口腕的日均生长率(η)[10],其计算参考下列公式(1):
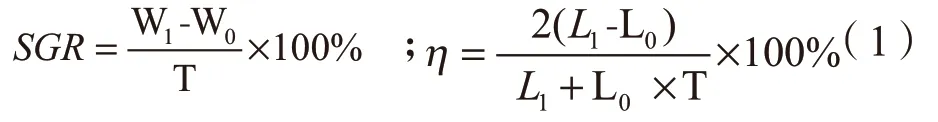
式中:W1为最终体重;W0为初始体重;L1为最终伞径或口腕长(cm);L0为初始伞径或口腕长(cm);T 为试验时间(d)
1.2.3 数据统计分析 采用SPSS13.0 软件对试验数据进行统计分析,并采用Ducan’s 多重比较方法进行显著性检验。
2 结果与分析
2.1 不同饵料组合对马来沙水母口腕长影响
由图1 可知,各饵料组对马来沙水母口腕长的影响明显,试验结束时MF 和MP 组水母口腕长度最大,且两者间差异不显著(P>0.05,下同);口腕长均值最小的是MM 和MS 组,两组间差异不显著。MM 和MS 组水母在饲养10 d 时出现口腕生长的峰值,且与其他试验组有显著性差异(P<0.05,下同)。
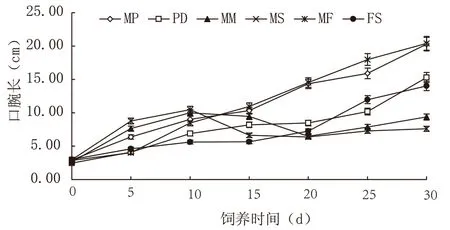
图1 不同饵料组合对马来沙水母口腕长的影响
2.2 不同饵料组合对马来沙水母伞径大小的影响
试验过程中,各试验组水母伞径均有增长(见图2)。饲养30 d 时,MP 组马来沙水母伞径最大,且与其他饵料组差异显著,其次为MF 组,剩余4 组表现不理想(各组间差异不显著)。MM 组在饲养10 d 时,出现伞径的峰值,与另外5个饵料组表现出显著性差异,随后变化不大。
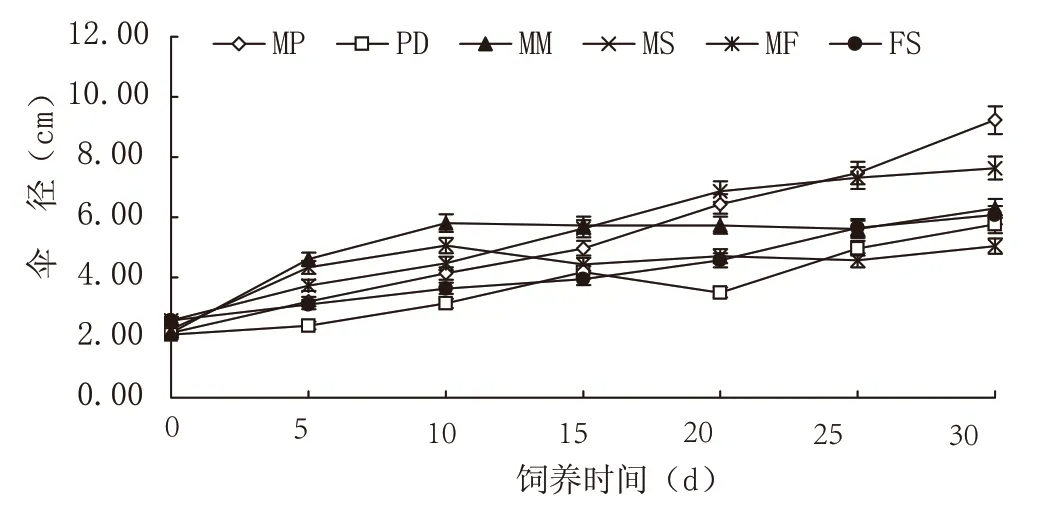
图2 不同饵料组合对马来沙水母伞径的影响
2.3 不同饵料组合对马来沙水母体重的影响
随着试验的开展,各组水母体重间差异日趋明显(图3)。饲养15 d 后,MP 组水母体重增长迅速,饲养30 d 时,MP 组水母体重达到40.3 g,与其余5个试验组差异显著;其次为MF 组,其水母体重最终达到24.13 g,增长明显;MS 组的水母体重增长值最小,但与PD 组、MM 组及FS 组差异不显著。MM 组体重在10 d 时达到峰值,随后试验期内变化较小。

图3 不同饵料组合对马来沙水母体重的影响
2.4 不同饵料条件下马来沙水母的生长情况
从表1 中可以看出,不同饵料条件饲养30 d 后,MP 组马来沙水母的特定生长率达到132%;其次是MF 组,SGR 值为77%;而MS 组的SGR 值最小,仅16%。MP 组水母的伞径日均生长率最高,为4.14%;其次为MF 组(3.32%)。6个饵料组合中MF 组水母的口腕长日均生长率值最高,为5.02%;MP 组排第二(4.98%);MS 组的ηDM 和ηOL 值在6 饵料组中都最小。

表1 不同饵料组合对马来沙水母生长的影响
2.5 MP 组口腕长、伞径与体重的相关性分析
从表2 中可以看出,口腕长、伞径与体重均具有高度线性正相关,Pearson 相关系数分别为0.941、0.960,且差异极显著(P <0.01,下同)。通过回归分析建立30日龄马来沙水母体重的回归方程:W=18.579×DM-5.454×OL-24.649,其中W 为水母体重(g),DM 为水母伞径(cm),OL 为水母口腕长(cm)。多元回归关系的显著性检验显示该方程达极显著水平。同时,伞径和口腕性状对体重的决定系数R2=0.956,R2值较大。
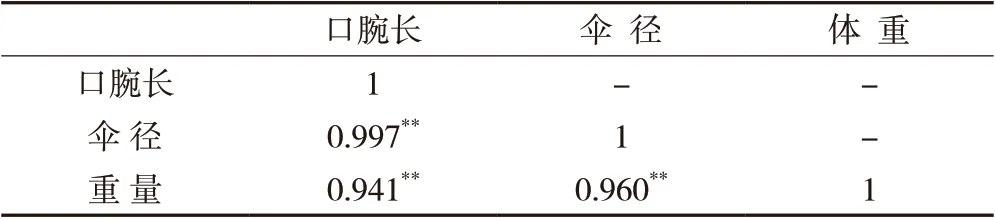
表2 口腕长、伞径和体重间的相关分析
3 讨 论
在自然界中,大型水母的主要饵料是海洋动物的卵和幼虫等浮游生物[11]。当饵料丰富且其他条件适宜时,水母生长迅速,个体数量也极速增加[12]。但受条件限制,水族馆难以模仿自然环境为水母提供多样性饵料,从而导致与自然界中的个体相比,饲养水母的伞径、重量及体色等性状会有一定差异。
鉴于培养海月水母的时间成本和经济成本较高,实际生产中难以做到全天投喂海月水母,所以研究未设海月水母饵料组,而只比较了投喂颗粒饲料、糠虾、丰年虫、海月水母+颗粒饲料、海月水母+糠虾和海月水母+丰年虫6种饵料组合对马来沙水母口腕长、伞径和体重3个生长指标的影响。在为期30 d 的试验过程中,马来沙水母各项生长指标均有明显增长,但最终试验效果差异较大。试验结束时,海月水母+颗粒饲料组的马来沙水母伞径平均值为9.23 cm,体重达到40.3 g,此两项指标均与其余5个试验组差异显著,而糠虾组水母的平均伞径为5.03 cm,体重仅有6.23 g。通过比较各试验组体重特定生长率,海月水母+颗粒饲料组水母的特定生长率高达132%,而伞径和口腕长的日均生长率数据进一步说明该饵料组合对马来沙水母生长的作用显著优于其他5 组饵料。出现这样结果的原因有3个方面:第一,杨为东[13]在研究不同饵料对咖啡金黄水母(Chrysaora melanaster)幼体伞径的影响时发现,饵料为海月水母的试验组水母在试验结束时平均伞径最大,笔者在日常饲养中亦发现,投喂海月水母的马来沙水母、彩色金黄水母(Chrysaora colorata)、咖啡金黄水母生长情况一直良好,个体伞径大、口腕长、形象美观,这在一定程度上可证明这一研究结果;第二,研究选用的饲料是海水鱼苗专用微颗粒饲料,营养价值较高[14-15],相对其他非海月水母饵料而言,可较大程度地满足马来沙水母的营养需求;第三,在整个试验周期内,海月水母+颗粒饲料组的水母摄食情况良好,未出现饵料摄入一段时间后部分被排出的现象,说明此饵料组合能够被充分消化吸收。
与其他4 组饵料组合不同,糠虾组和海月水母+糠虾组的马来沙水母在饲养10 d 时,口腕长、伞径和体重出现拐点,饲养后期20 d 的变化较小。试验中观察发现饲养10 d 后,部分水母口腕变得平滑,对饵料的吸附能力下降,出现此现象的原因尚需进一步研究;另外还发现,饲养10 d 后,水母虽摄入糠虾,但之后又会将部分饵料排出胃囊,降低水母对饵料的消化吸收率,从而直接影响了水母的生长。这种现象在咖啡金黄水母、海月水母和半球美螅水母(Clytia hemisphaerica)试验中也出现过[13,16-18](但具体发生机制目前尚不明确。且有研究显示,饵料摄入不足会导致海蜇出现生长停滞甚至个体缩小的负增长现象[19]。摄食量不足与生长停滞形成恶性循环链,该研究中糠虾组和海月水母+糠虾组饲养10 d 后出现的状况与之相似。
试验数据表明,海月水母+颗粒饲料组的马来沙水母生长速度快、个体大且形象美观,符合水族馆对该水母的饲养研究预期,因此笔者对该组水母生长指标进行了相关性分析。30日龄的马来沙水母,其口腕长与体重、伞径与体重、口腕长与伞径均表现出极显著正相关,相关系数分别为0.941、0.960和0.997。伞径、口腕长对体重的回归关系为极显著水平,而且R2值较大,说明回归方程显著存在,伞径和口腕长对体重有较大的影响。通过对所选指标进行相关性分析并建立回归方程,在尽量减少人为因素对水母的生长影响情况下,可根据部分性状的数据对主要性状进行估测,为马来沙水母的大规模繁殖饲养以及育种亲体筛选等工作提供参考。
为实现马来沙水母生长条件的最优化,综合比较3个生长指标在整个试验期的变化,得出以下投饵方案:马来沙水母伞径在2~5 cm 时,选用海月水母+糠虾饵料组合,即上午投喂海月水母,下午投喂糠虾;伞径超过5 cm 后,选用海月水母+颗粒饲料组合。因此,在马来沙水母的饲养过程中,管理人员要根据不同的生长阶段选择合适的饵料,以确保水母的正常生长。
[1]高尚武,洪惠馨,张士美. 中国动物志:无脊椎动物(第二十七卷)[M]. 北京:科学出版社,2002. 203.
[2]张先锋,王士莉. 走进水族馆[M]. 北京:北京科学技术出版社,2010. 16-27.
[3]Uchida T,Sugiura Y. On the ephyra and postephyra of a semaeostome sedusa, Sanderia malayensis Goette[J]. Fac Sci Hokkaido Univ Ser VI,Zool. 1975,19(4):879-881.
[4]Uchida T,Sugiura Y. On the polyp of the scyphomedusa, Sanderia malayensis and its reproduction[J]. Fac Sci Hokkaido Univ Ser VI,Zool. 1978,21(2):279-285.
[5]Laetitia A,Gerhard J. New insights into reproductive traits of scyphozoans:special methods of propagation in Sanderia malayensis Goette,1886(Pelagiidae, Semaeostomeae) enable establishing a new classification of asexual reproduction in the class Scyphozoa[J]. Mar Biol.,2009,156(7):1411-1420.
[6]郭嘉瑱,王海铭,王文章. 马来沙水母各发育阶段形态[J]. 齐鲁渔业,2012,29(1):12-15.
[7]郭玲玲,赵 文,杨为东,等. 光照对马来沙水母幼体生长的影响[J]. 大连海洋大学学报,2012,27(1):18-21.
[8]马喜平,凡守军. 水母类在海洋食物网中的作用[J]. 海洋科学,1998,(2):38-41.
[9]熊 瑛,王云龙,汤建华,等. 黄海南部大型水母暴发区中小型浮游动物生态特征[J]. 生态学杂志,2009,28(10):2063-2068.
[10]李晓东,刘铁钢,张长新,等. 几种生物饵料对海蜇幼蜇生长的影响[J]. 水产科学,2003,22(3):4-6.
[11]洪惠馨. 水母与海蜇[J]. 生物学通报,2002,37(2):13-16.
[12]Brodeur R D,Sugisaki H,Hunt G L. Increases in jellyfish biomass in the Bering Sea:implications for the ecosystem [J]. Marine Ecology Progress Series,2002,233:89-104.
[13]杨为东,郭玲玲,赵 文,等. pH、盐度和不同饵料对咖啡金黄水母幼体生长与存活的影响[J]. 大连海洋大学学报,2011,26(3):223-226.
[14]赵金柱,艾庆辉,麦康森,等. 微粒饲料替代生物饵料对大黄鱼稚鱼生长、存活和消化酶活力的影响[J]. 水产学报, 2008,32(1):91-97.
[15]于海瑞,麦康森,马洪明,等. 微颗粒饲料与冰冻桡足类对大黄鱼稚鱼生长、存活和体成分的影响[J]. 水生生物学报,2012,36(1):49-56.
[16]Riisgard H U,Caroline V M. Clearance rates of ephyrae and small medusae of the common jellyfish Aurelia aurita offered different types of prey [J]. Journal of Sea Research,2011,65:51-57.
[17]陈渝萍. 半球美螅水母摄食生态的初步研究[D]. 厦门:厦门大学,1995.
[18]王 楠,李超伦. 水母伞径和饵料密度对海月水母(Aurelia sp. 1) 捕食率的影响[J]. 海洋与湖沼,2012,43(3):480-485.
[19]刘顶海,张继红,陈四清,等. 海蜇池塘养殖生长规律的研究[J]. 湖南农业科学,2011,(17):122-125.
