贵州3处保护区生物廊道景观格局与连通性浅析
2015-09-16粟海军喻理飞吴际通
粟海军,喻理飞,吴际通,2
(1.贵州大学喀斯特生态环境研究中心,贵州 贵阳 550025;2.贵阳市森林资源管理站,贵州 贵阳 550005)
贵州3处保护区生物廊道景观格局与连通性浅析
粟海军1,喻理飞1,吴际通1,2
(1.贵州大学喀斯特生态环境研究中心,贵州 贵阳 550025;2.贵阳市森林资源管理站,贵州 贵阳 550005)
以黔东武陵山至苗岭山系间的梵净山国家级自然保护区、佛顶山省级自然保护区及雷公山国家级自然保护区为研究对象,选定生物廊道宽度为2 km,以3个自然保护区现有的边界为基础,向外扩展平均约50 km为缓冲区并彼此相连形成可供成为生物廊道的互通区域,对这一区域景观特征及廊道连通性进行研究。结果表明:①三者景观多样性指数(SHDI)为0.97,景观斑块分布较均匀;②在景观尺度水平上,三者间不同类型斑块与不同因子斑块结合度(COHESION)均较高,达99以上,表明三者间连通性良好;③根据连通性路径分析结果,佛顶山北段至梵净山区域连通性良好,连通因子占廊道面积达88.90%,佛顶山南段至雷公山区域,连通因子占86.12%,生物廊道连通因子主要为林地构成,阻隔因子主要为耕地及建设用地。研究结果揭示了3个保护区内及其之间生物廊道的景观斑块特征与生物物种扩散的景观连通性,可为构建科学有效的生物廊道与区域自然保护区群提供参考。
景观格局;生物廊道;连通性;自然保护区;贵州省
生物廊道(Biological Corridor或Bio-corridor)一般指连接破碎化生境并适宜生物生活、移动或扩散的通道[1-3],是目前国际上公认的连接自然保护热点区域与构建自然保护区群的最为有效的形式[4-8]。贵州黔东及黔东南地区是贵州生物多样性最丰富的区域之一,覆盖了贵州生态功能区划中的“黔东北梵净山森林生态系统生物多样性关键区域”及“黔东苗岭东段雷公山常绿阔叶林生物多样性关键区域”[9],包含梵净山与雷公山2个国家级自然保护区及“人与生物圈保护区”。而佛顶山省级自然保护区正处于这2大区域之间,成为承接武陵山脉与苗岭山脉的重要生物廊道结点[10]。因此,为揭示保护区群构建对于区域生物多样性保护的重要性,探索大区域尺度上保护区群建立的生物廊道效应,本文通过研究保护区群尺度上的景观格局和生物廊道扩散传递的连通性,以理清3个重要区域生物扩散传递条件、通道和主要阻隔因素,以及提高连通性的对策。
1 研究地概况
贵州佛顶山省级自然保护区位于黔东北部石阡县境内,以中亚热带常绿阔叶林森林生态系统及其珍稀动植物物种为保护对象。佛顶山自然保护区地理位置为27°15′—27°25′N、107°56′—108°12′E,总面积12634.54 hm2,森林覆盖率为87%,为一沿北东向南西延伸的高大脊状山岭,是夹持于北东向断裂带间的一个巨大构造透镜体组成的地垒式山地,沿着北东向区域性大断裂与其北东面的梵净山一脉相承。山脉主峰海拔1869.3 m,是贵州东部仅次于梵净山的第二大高山。佛顶山倚武陵山脉主峰梵净山(2583 m)而成姊妹峰,同时又介于梵净山与西南面苗岭山脉的雷公山(2350 m)之间[10]。
梵净山国家级自然保护区于1978年建立,1986年晋升为国家级保护区并列入国际“人与生物圈”保护区网络成员。面积41900 hm2,森林覆盖率90%。主要保护对象是以黔金丝猴、珙桐(Davidiainvolucrata)等为代表的珍稀野生动植物及原生森林生态系统。雷公山自然保护区于1982年建立,2001年晋升为国家级自然保护区,总面积47300 hm2,森林覆盖率达83%。是以保护台湾杉(Taiwaniacryptomerioides)等珍稀生物为主的亚热带山地森林生态系统类型的自然保护区,也是长江水系与珠江水系的分水岭[11-12]。
介于两者间的佛顶山,距梵净山、雷公山分别为79、108 km。距离虽不远,但依《中国自然地理》[13]区划,梵净山与佛顶山均属华中区西部山地高原亚区黔桂湘低山丘陵省中的武陵山地区域(VI44),而雷公山属同一亚区下的贵州高原省苗岭丘原区域(VI42);也分属不同气候区,梵净山和佛顶山属北亚热带贵州区,雷公山为中亚热带川鄂湘黔区;在中国植被区划上,梵净山、佛顶山属中亚热带常绿阔叶林南部亚地带——贵州山原,栲类、青冈林、石灰岩植被区,雷公山属三江(都江、浔江、融江)流域山地,栲类、木荷林、石灰山植被区[13-14],因此三地生物多样性的交叉融汇具有重要意义。
在山体垂直特征差异上,山体高度以梵净山最高、山脚海拔最低,相对高度最大;雷公山次之;佛顶山最小。佛顶山处于武陵山主峰梵净山向苗岭主峰雷公山过渡的一个山体,山体主要分布于海拔1500 m以下(表1)。三者低于1500 m高度的主体分别占62.75%、96.62%、71.59%。

表1 佛顶山与梵净山、雷公山山体特征比较
生物廊道的宽度依物种而异,但较宽的廊道适于大多数物种[15],本文选定生物廊道宽度为2 km。为分析三者间最优连通的生物廊道,考虑到三者间的相距距离,以3个自然保护区现有的边界为基础,向外扩展平均约50 km为缓冲区并彼此相连形成可供成为生物廊道的互通区域(图1),总面积35328.81 km2,包括黔东及黔东南地区的江口、松桃、印江、石阡、施秉、黄平、台江、镇远、凯里、雷山、麻江等县。本文主要对这一区域景观特征及廊道连通性进行研究。

图1 梵净山、佛顶山及雷公山扩展范围及其生物廊道示意图
2 研究方法
利用2010年土地利用现状数据及森林资源二类调查数据,将研究区分为林地、草地、水体、建设用地和耕地5大地类景观类型(图1)。
为确定景观格局分析的最佳尺度,利用ArcGIS 9.3软件的矢量面转栅格工具(Polygon to Raster)将矢量数据转为栅格数据,栅格单元大小依次设为25、50、100、200、300、400、500、600、700、800、900、1000、1500 m。利用FRAGSTATS 3.3计算景观特征指数,由于景观指数随着空间粒度大小(grain size)变化而变化[16- 17],因此需要定量化分析各景观指数在不同尺度上的基本不变区域,然后确定不同指数的公共稳定区域,在第一公共区域中选择中等偏上的粒度作为景观格局分析的最佳尺度[18-19]。
1) 斑块平均大小(Mean patch size):代表一种平均状况,可以指征景观的破碎程度。公式为:MPS=A/N,式中:A为整个景观或某一景观类型的面积(hm2);N为斑块数目。
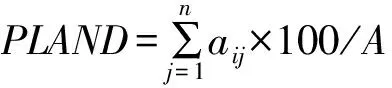
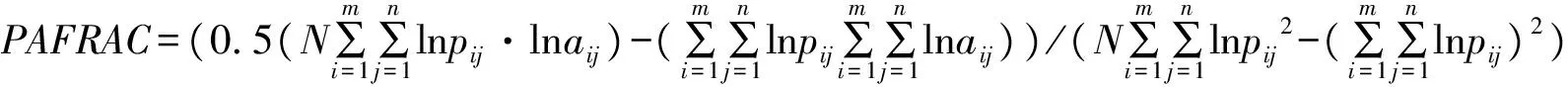

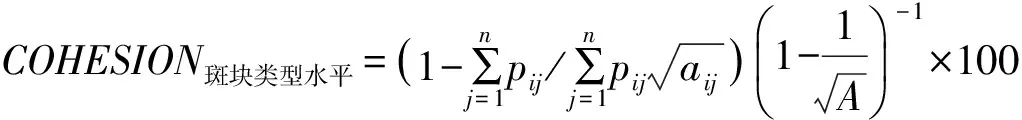
3 结果与分析
3.1 景观格局指数随尺度变化特征与景观尺度确定
总体上看,景观指标(图2)随尺度增加呈现不同的趋势:平均斑块大小指数呈先逐渐递减后增加的趋势,其基本不变区域是100~200、800~900 m;周长面积比分维数指数呈迅速下降后逐渐稳定趋势,其基本不变区域是100~300、400~600、700~1500 m;斑块结合度指数呈逐渐下降趋势,其基本不变区域是200~300、800~900 m;香农多样性指数呈波动变化,其基本不变区域为50~200、400~600、700~900 m。在选择尺度时,若想体现研究区域的景观特征信息,选择相对稳定的后一尺度域是为适宜尺度的较好取值范围,因此,确定本研究中景观类型图进行景观格局分析的适宜粒度为200 m。

图2 景观格局指数的尺度效应图
3.2 生物廊道及缓冲区域的景观构成分析
根据前述面积界定,研究区总面积达350余万hm2,其中林地占到58.36%,耕地占26.12%,草地占15.16%,建设用地和水体分别仅占0.25%和0.12%,因此林地构成景观的基质。
平均斑块大小反映了不同斑块类型的破碎程度,由表2可知,林地的MPS最大,显著大于其它斑块类型,说明林地斑块最完整(249.06 hm2),其次为草地(77.60 hm2),而耕地(26.15 hm2)、建设用地(18.93 hm2)和水体(9.73 hm2)都比较破碎。
周长面积比分维数是反映斑块类型形状复杂程度的指数,值域范围为1≤PAFRAC≤2,值等于1时代表形状最简单的正方形或圆形,值等于2时代表形状最复杂的斑块类型[20]。林地、草地和耕地的分维数最高,均为1.44;其次为水体,为1.38;最小的为建设用地,为1.34,说明林地、草地和耕地的形状相对比较复杂;水体和建设用地形状较简单,比较容易受人类活动的影响。
香农多样性指数(SHDI)与景观类型数以及类型的面积百分比有关,与均匀度指数存在显著正相关关系,而与优势度指数存在显著负相关关系[21]。与浙江天目山保护区[22]及福建戴云山保护区[23]等南方保护区景观多样性相比,梵净山、佛顶山、雷公山景观多样性指数仅达0.97,相比而言景观类型分布较为均匀,但SHDI值从>1开始,值越大则多样性越丰富,说明除森林斑块外的其他类型斑块也多有存在。生境斑块的多样性与复杂性,可能有利于某类物种的迁徙、移动与占居建群,但建设用地、水体等斑块的存在无疑对大多数物种的生存和扩散具有阻碍作用[5,24]。
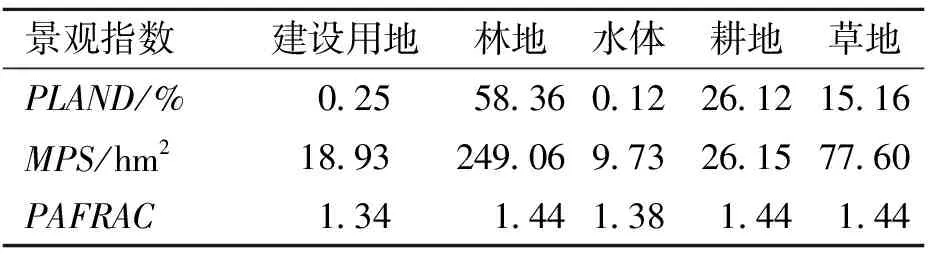
表2 梵净山、佛顶山、雷公山景观格局指标
3.3 生物廊道景观格局连通性
连通性指生物廊道在空间上如何连接,它是生物廊道结构的量度指标。连通性较高的景观格局有利于生物的迁徙扩散,但就某一种生物而言,其迁徙扩散不仅取决于生境斑块结构的连通性,更取决于种群状况、扩散方式及与之相适应的生境内部质量条件等[1,25]。基于物种扩散的复杂性,暂无法确定三者间生物廊道针对某一物种的连通性、连接性及适宜性等问题,而主要通过斑块结合度指数讨论景观尺度上的一般连通性(表3)。
在梵净山—佛顶山—雷公山区域(F—F—L)中,以佛顶山为中心,分为南北两段,其北段为佛顶山—梵净山区域(F—F)、南段为佛顶山—雷公山区域(F—L),从景观水平看,F—F—L整体、F—F、F—L之间斑块结合度均在99.27以上,比人为干扰较大的武汉森林保护区[26]高,而与结合度较好的东北三江平原流域[27]相近。说明3个山体之间景观连通性较好,三者间易于形成良好的生物廊道,而居于中间的佛顶山可成为连通梵净山与雷公山之间生物传播的重要连续点和通道。

表3 研究区不同类型斑块结合度指数
*:F—F—L为佛顶山—梵净山—雷公山区域;F—F为佛顶山—梵净山区域;F—L为佛顶山—雷公山区域。下同。
就斑块类型对于植物种群的扩散而言,林地、灌丛与草地生境类型为其主要扩散生境;耕地因人为干扰频繁不利于植物建群生长,建设用地也不适于植物扩散。对一般野生动物而言,林地、草地、水体、部分耕地均可成为其活动区域,也是其传播植物繁殖体的主要区域,但对于大中型兽类及两爬类,则要求较大面积的森林与人为干扰较少的优良生境,因此大面积连通的森林生态系统的存在,较有利于动物的移动迁徙。从斑块类型水平来看,林地斑块结合度最高,连通性也最好;其次为草地和耕地;最差的为建设用地和水体。因此林地的高连通性为生物的传播提供了重要条件。
基于此,林地、草地、水体斑块类型构成3个山体间的生物扩散的连通因子,将斑块合并,构成生物可能较自由传播的通道。建设用地、耕地因构成生物传递的阻隔因子,将建设用地、耕地斑块合并,构成了生物自由传播的阻隔屏障。经计算,梵净山—佛顶山—雷公山区域(F—F—L)整体连通因子为99.95,阻隔因子为88.26,整体景观结合度水平达99.80。梵净山—佛顶山—雷公山区域F—F—L无论是其北段(F—F)还是南段(F—L),景观水平和斑块类型水平的连通性因子的结合度指数都高于90,景观连通性很高,阻隔因子的结合度虽较高,但仍较低于连通因子。可见其结果与前述斑块结合度分析较为一致;3个保护区间具备良好的生物廊道植被生境连通基础。
3.4 F—F—L区域生物传递的通道途径分析
根据图1分析佛顶山北段区域,即从佛顶山至梵净山区域,其东面林地、草地、水体斑块等连通因子分布较多,且连接成较大的连通性斑块。西面林地、草地、水体斑块较多被建设用地和耕地等阻隔性因子所分离,尤其是思南、印江、石阡境内各乡镇,大量建设用地和耕地影响生物的传递。采用斑块连通性最短距离作图,最短距离约79.12 km,路径是沿武陵山脉向北延伸,经铜仁市江口县闵孝、官和乡后进入黔东南州施秉县龙田、上寨乡后又进入铜仁市石阡县坪山进入佛顶山保护区。以2 km廊道宽度计算,连通性因子所占廊道面积达88.9%,以林地类型为主,占80.74%;阻隔因子为耕地类型,占11.10%(表4)。

表4 2 km宽度的廊道中各地类面积及所占比例
研究区南段,即从佛顶山至雷公山区域,其东面林地、草地、水体斑块等连通因子分布较多,且连接成较大的连通性斑块。西面林地、草地、水体斑块较多被建设用地和耕地等阻隔性因子所分离,尤其是凯里市、麻江县境内各乡镇,大量建设用地和耕地影响生物的传递。廊道最短距离为108.69 km,路径是向北延伸到苗岭雷公山,经黔东南州施秉县马溪、城关、杨柳塘、双井乡(镇),台江县革一、施洞、老屯、台拱、台盘、南宫乡(镇)进入雷公山国家级自然保护区。整个廊道中连通因子面积占86.12%,其中林地占绝大部分,其次为草地,水体极少;阻隔因子仅为耕地,占13.88%。可见加强这些路径的植被恢复与生态环境建设,可以实现构建距离最短、阻隔最小而具备较好植被生境条件的生物廊道,这对于指导区域自然保护区群与生物廊道建设具有重要现实意义[4-5,28]。
4 结论与讨论
不同生物物种迁徙移动、传播扩散及占居新地建群发展所依赖的生物廊道存在很大差异,但对于大多数动物物种而言,具有良好隐蔽与食物条件的植被带是生物廊道构成的重要条件,而对于很多种子植物而言,在空间上连续的母树分布,也有利于植物种子的传播与种群的扩散[1,4,6,28]。因此,在缺乏针对具体物种种群监测数据的情况下,研究景观尺度上的廊道特征与连通性,对指导区域生物廊道建设,建立自然保护区群具有重要的现实意义[24]。
佛顶山与梵净山同属武陵山脉和同一个生物地理单元,而雷公山则属苗岭山脉和另一个生物地理单元。佛顶山处于梵净山与雷公山之间,客观上成为2个重要生物多样性热点区域的生物廊道结点。
将林地、草地、水体作为生物传递的连通性因子,建设用地、耕地作为阻隔性因子,采用斑块连通性最短距离建立带宽2 km的廊道,佛顶山与梵净山之间最短的景观廊道约80 km,连通因子面积占廊道的88.90%,阻隔因子占11.10%;佛顶山与雷公山之间最短的景观廊道约110 km,连通因子面积占廊道的86.12%,阻隔因子占13.88%。表明研究区域的景观连通性较高,有利于生物的传递与交流,而廊道的阻隔因子主要是耕地和建设用地,且耕地占据主导作用,因此通过合理科学的退耕还林规划与生境营造,可有针对性地增加廊道的连通性[25]。可见,根据连通性分析结果,可得到生物廊道的最佳通道途径,有利于指导加强现存最佳通道的保护与开拓和建设新的生境通道。
生物廊道的建立对于珍稀濒危种的保护,以及构建区域生物多样性保护区群都具有重要作用[24]。依据景观格局特征进行一般性的连通性分析,能够从宏观尺度上把握生物廊道连通状况,但某一物种的生物廊道连通性却与该物种的扩散生物学特性及生境需求有关[2,8],因此,深入系统的生物廊道研究与构建,还需要开展科学的样方设置与长期生物监测研究才能实现。
[1]李正玲,陈明勇,吴兆录.生物保护廊道研究进展[J].生态学杂志,2009,28(3):523-528.
[2]Saunders D A,Hobbs R J,Margules C R.Biological consequences of ecosystem fragmentation:a review[J].Conservation biology,1991,5(1):18-32.
[3]Simberloff D,Cox J.Consequences and costs of conservation corridors[J].Conservation Biology,1987,1(1):63-71.
[4]Rouget M,Cowling R M,Lombard A T,et al.Designing Large-Scale Conservation Corridors for Pattern and Process[J].Conservation Biology,2006,20(2):549-561.
[5]姜明,武海涛,吕宪国,等.湿地生态廊道设计的理论、模式及实践:以三江平原浓江河湿地生态廊道为例[J].湿地科学,2009,7(2):99-105.
[6]Falcy M R,Estades C F.Effectiveness of Corridors Relative to Enlargement of Habitat Patches[J].Conservation Biology,2007,21(5):1341-1346.
[7]张饮江,金晶,董悦,等.退化滨水景观带植物群落生态修复技术研究进展[J].生态环境学报,2012(7):1366-1374.
[8]Prendergast J R,Quinn R M,Lawton J H.The gaps between theory and practice in selecting nature reserves[J].Conservation biology,1999,13(3):484-492.
[9]贵州省环境保护厅,贵州省发展与改革委员会.贵州省生态功能区划[Z].贵阳,2005.
[10]喻理飞,李明晶,谢双喜.贵州佛顶山自然保护区科学考察集[M].北京:中国林业出版社,2000.
[11]周正贤.梵净山研究[M].贵阳:贵州人民出版社,1990.
[12]张华海,张旋.雷公山国家级自然保护区生物多样性研究[M].贵阳:贵州科技出版社,2007.
[13]中国科学院中国自然地理编辑委员会.中国自然地理动物地理[M].北京:科学出版社,1979.
[14]吴征镒.中国植被[M].北京:科学出版社,1980.
[15]朱强,俞孔坚,李迪华.景观规划中的生态廊道宽度[J].生态学报,2005,25(9):2406-2412.
[16]冯永玖,刘艳,周茜,等.景观格局破碎化的粒度特征及其变异的分形定量研究[J].生态环境学报,2013(3):443-450.
[17]Urban D L.Modeling ecological processes across scales[J].Ecology,2005,86(8):1996-2006.
[18]赵文武,傅伯杰,陈利顶.景观指数的粒度变化效应[J].第四纪研究,2003,23(3):326-333.
[19]张东水,兰樟仁,邱荣祖.闽江口湿地遥感影像最佳景观观察尺度的选择[J].遥感信息,2006(4):30-32.
[20]张会儒.天然林资源动态时空分析评价技术[M].北京:中国林业出版社,2006.
[21]布仁仓,胡远满,常禹,等.景观指数之间的相关分析[J].生态学报,2005,25(10):2764-2775.
[22]吴兆艳,汤孟平,洪海峰,等.浙江天目山自然保护区森林景观格局分析[J].西南林业大学学报,2011(5):62-66.
[23]梁美霞.基于景观格局分析的戴云山自然保护区景观生态建设[J].福建林业科技,2013(1):127-132.
[24]陈利顶,傅伯杰,刘雪华.自然保护区景观结构设计与物种保护:以卧龙自然保护区为例[J].自然资源学报,2000,15(2):164-169.
[25]陈利顶,傅伯杰.景观连接度的生态学意义及其应用[J].生态学杂志,1996,15(4):37-42.
[26]滕明君,周志翔,王鹏程,等.基于RS/GIS的武汉市九峰城市森林保护区景观结构特征及规划对策[J].长江流域资源与环境,2010(1):79-85.
[27]罗春雨.三江平原挠力河流域景观多样性分析[J].国土与自然资源研究,2009(2):58-60.
[28]Kaiser J.Building a case for biological corridors[J].Science,2001,293(5538):2199.
Landscape Patterns and Connectivity of Bio-corridors between ThreeNature Reserves in Guizhou Province
SU Hai-jun1,YU Li-fei1,WU Ji-tong1,2
(1.KarstEco-environmentResearchCentreofGuizhouUniversity,Guiyang550025,Guizhou,China;2.ManagementStationofForestResourcesofGuiyangCity,Guiyang550005,Guizhou,China)
The bio-corridors with 2km width between Fanjishan National Nature Reserve,Fodingshan Provincial Nature Reserve,and Leigongshan National Nature Reserve,which are located between Wuling Mountain Chain and Miaoling Mountain Chain in the east and southeast of Guizhou province,China,were studied on the characteristics of landscape and the connectivity of bio-corridors,and the 50 km-expanded buffer area outside of respective Nature Reserves as well.The results showed:①among these three Reserves,theSHDIwas 0.97,which indicated that patches of landscape distribute evenly;②at the scale of landscape,theCOHESIONsto different types and factors were over 99 high to all Reserves,which indicated that these three Reserves connect well;③according to the connectivity analysis results,88.9% area of bio-corridor from the north of Fodingshan to Fanjingshan is connected well,and 86.12% from the south of Fodingshan to Leigongshan,in which consisted of forest land as connecting factor and cultivated and construction land as blocked factors.The characteristics of landscape on bio-corridors and the connectivity for species dispersal among these three neighboring Nature Reserves were discovered in this paper,which will be referenced to build an effective bio-corridors and Nature Reserve network regionally.
landscape patterns;bio-corridor;connectivity;nature reserves;Guizhou Province
2014-03-10;
2014-04-21
国家教育部科技重点项目(210198);国家科技支持计划项目子专题(2011BAC02B0205-01、2011BAC02B0202);贵州省国际科技合作项目(黔科合外G字[2012]7013号);贵州省林业厅佛顶山保护区综合科考项目资助
粟海军(1978—),男,贵州锦屏人,贵州大学副教授,博士,从事生物多样性保护与自然保护区学研究。E-mail:haijun_su@163.com。
10.13428/j.cnki.fjlk.2015.01.012
S759.9
A
1002-7351(2015)01-0055-07
