自发性眨眼对瞳孔反应和注视位置的影响
2015-09-16杨开富颜红梅尧德中李永杰
高 鑫 杨开富 颜红梅 尧德中 李永杰
(电子科技大学 生命科学与技术学院, 成都 610054)
自发性眨眼对瞳孔反应和注视位置的影响
高 鑫 杨开富 颜红梅 尧德中#李永杰*
(电子科技大学 生命科学与技术学院, 成都 610054)
瞳孔反应和注视位置是眼动研究中重要的监测指标,但在实验过程中,常常会受到自发性眨眼的干扰。目前,多数研究者只关注到眨眼过程中短暂的数据丢失和波动,而忽略了眨眼对这些变量更广泛的影响。因此本文系统地分析了眨眼对瞳孔反应和注视位置的影响。本研究采用简单的长时间注视任务,使用光学眼动仪记录22个人类被试的瞳孔和注视位置的数据并在其中精确地提取眨眼。瞳孔反应和注视位置的时间序列以眨眼开始和结束对齐。结果表明,在眨眼结束后,瞳孔反应发生长达4 s的大幅度波动(约12%),其中在300~1 700 ms间,瞳孔值显著地低于基线水平(P 眨眼; 注视误差; 瞳孔;注视位置 随着光学眼动仪(video eye tracker)的普及,越来越多的研究者通过记录瞳孔反应和注视位置来探究人的心理活动。瞳孔是人眼内虹膜中心的孔状结构,是光进入人眼的门户。瞳孔除了对进入人眼内的光线反应,还可以反映人类的心理活动。早期研究发现,瞳孔的大小可以反映出人类的唤起水平以及认知负荷[1]。但对瞳孔反应的研究随着脑电等技术的兴起而逐渐没落。近年来,由于瞳孔大小的数据可以很容易地通过眼动仪获取到,因此越来越多的研究者重新转向了瞳孔反应在认知活动中的研究,例如,近年来研究者发现,瞳孔的变化和人的预测、推理、决策以及知觉切换等高级认知活动有关[2-4]。注视位置也是反应心理活动的重要指标。由于注视位置和视觉注意紧密相连[5-6],通过记录人的注视位置,可以推测出当前的注意目标。但在这些研究过程中,瞳孔和眼动数据常常会受到自发眨眼的干扰。虽然眨眼有很重要的生理功能,例如保持眼内的湿润、防止异物进入眼睛等。但它发生的频率要远远高于它在生理功能上的需要。有研究者指出,人处于眨眼的时间可以达到人类清醒时间的10%左右[7]。因此,频繁的眨眼会阻断视觉输入,并造成瞳孔和眼动位置数据的丢失和波动。 然而,目前的研究者都只关注到眨眼过程中短暂的数据丢失和波动,而忽略了眨眼可能产生的持续影响。例如,眨眼会引起进入眼内的光线变化,这可能会使对光强度敏感的瞳孔产生持续的反应。眨眼还会造成视觉目标的短暂丢失;并且眨眼本身会伴随着眼动[8-9],这也可能造成眨眼后注视位置的偏移。但目前针对眨眼对瞳孔和注视位置影响的研究却很少,并且研究结果相互矛盾。例如,Fukuda[10]和Hupé[11]在认知加工任务中测量了眨眼后的瞳孔反应,前者发现了小幅度的瞳孔扩大,而后者则观察到了大幅度的瞳孔收缩。由于以往的众多研究表明瞳孔和眨眼反应都和认知活动紧密相关[1, 12],因此,在认知加工任务中去考察眨眼后的瞳孔反应会造成结果的混淆。同样的,对于眨眼对注视位置影响,之前只在动物研究中(猴子)测量过[13],目前还没有针对人类的精确测量。因此,本研究使用更为单一的注视任务,以排除认知加工因素对瞳孔反应和注视位置的影响。同时,通过对眨眼前后的瞳孔反应和注视位置的时间序列的精确分析,发现眨眼对瞳孔反应会产生持续约4 s、最大幅度约为12%的波动。同时,眨眼会造成持续时间约200 ms、幅度约为1°的注视位置的偏移。此外,眨眼后瞳孔变化也会引起由光学眼动仪记录到的注视位置的变化,其最大幅度为0.26°。 1.1实验被试和实验装置 实验被试共有22名在校大学生,其中男13名,女9名。被试的年龄为22~26岁。实验采用Eyelink 2000型眼动仪(SR Research Ltd.,加拿大)记录被试双眼的瞳孔面积和注视位置。数据采样频率为1 000 Hz。显示器的分辨率为1 024像素×768像素。实验过程中,被试眼睛正对屏幕中心,离屏幕的距离为71 cm。为减少头动引起的记录误差,被试的下颚和前额放在特制的支架上。实验的刺激程序由Matlab的心理学工具箱编写(Psychotoolbox)[14-15]。 1.2试验流程 实验采用简单的注视任务。在每次测试中,被试需要注视屏幕中心的注视点(黑色、0.4°)45 s。在下一次测试之前,被试有8 s的休息时间,期间呈现空白灰屏。每个被试需要做50次实验,这些实验分成5组,每组10次,每做完一组实验,被试可以根据自身状态自由休息。 1.3数据分析 1.3.1数据获取 数据通过眼动仪自带软件导出(DataView)。其导出的数据包括:双眼在每一时间点上(每毫秒)注视位置的水平和垂直坐标,以及双眼在每一时间点上(每毫秒)的瞳孔面积。 1.3.2注视误差 使用注视误差的大小来表示眨眼对注视位置的影响。注视误差定义为:当前注视点相对于注视目标的欧氏距离。 1.3.3基线选取 采用眨眼前2 000 ms的瞳孔反应以及注视误差的平均值作为基线水平,并以此作为基准值和眨眼后的相应指标做对比(图2和图3)。 1.3.5误差范围 使用标准误来表示平均数的误差范围,即平均数±标准误。在绘制瞳孔反应及注视误差的时间过程中,也在每个时间点的被试间平均数(n=22)的上下标记出了一个标准误的误差范围,所有时间点的误差值连接起来形成了标准误包络线。其中,标准误表示为 (1) 式中,s代表被试间标准差,n代表样本数量,本研究中n=22。 1.3.6检测眨眼 光学眼动仪(video eye tracker)是通过计算瞳孔的中心和角膜反射位置的差值来确定注视的位置。在眨眼过程中,由于眼睑遮挡了瞳孔,会造成瞳孔数据的丢失,因此,目前的研究都是通过检测瞳孔的丢失值来确定眨眼。这种方法虽然能检测出眨眼的发生,但并不精确。因为瞳孔丢失的部分只代表眼睑闭合(eye closure)时的状态。一次完整的眨眼还包括眼睑向下和向上运动的过程。这一过程会造成很大的数据波动(见图1),但由于瞳孔没有完全被遮挡,因此不会产生数据的丢失,进而无法被常规的方法检测到。本研究将结合瞳孔变化速度和瞳孔丢失值来近似地检测眨眼的完整过程,进而更加精确计算眨眼对瞳孔和注视位置影响的时间过程。具体方法如下(见图1)。首先检测出瞳孔丢失值的区间(灰色部分)。为了减小记录噪声,用Savitzky-Golay数据平滑方法(Matlab函数:sgolayfilt)[16]对丢失瞳孔之前和之后200 ms的数据做平滑处理,其中阶数为1,带宽为41个采样点,并计算其瞳孔变化速度(求导数)。最后,在得到的速度曲线中,最小值之前最后一个趋近于0(±0.5之间)和最大值后第一个趋近0(±0.5之间)的采样点被定义为眨眼的开始和结束。需要说明的是, 在图1中,眨眼过程中记录到的瞳孔面积和注视位置的快速变化主要是由于眼睑运动引起的,例如眼睑对瞳孔的遮挡,并不必然意味着真正的瞳孔和注视位置的变化。 图1 定义眨眼的开始和结束(灰色部分代表瞳孔数据丢失的区间。从上至下分别为瞳孔变化速度、瞳孔面积、垂直注视位置和水平注视位置)Fig.1 The definition of blink onset and blink end (The light grey area represents the period where the pupil information is missing; panels from top to bottom denote the pupil changing velocity, the pupil area, vertical eye position and horizontal eye position, respectively) 1.4数据准备和统计检验 由于本研究记录被试双眼的瞳孔面积以及注视位置,以下的关于瞳孔面积及注视位置分析均为双眼平均的结果。眨眼开始前2 s和眨眼结束后4 s的瞳孔反应和注视误差数据分别以眨眼开始和结束点对齐,如果在这期间有其它的眨眼发生,或出现注视误差超过3°的数据点,那么这次眨眼的数据将被剔除。在参加实验的所有被试中(n=22)共检测出10 705(486.59±100.40)次眨眼,根据此数据剔除标准,有2 478(112.64±14.13)次眨眼的数据保留到后续的眨眼和注视位置的分析中。括号内为被试间均值±标准误,下文相同。为了检测每一时间点的数据与基线水平的差异是否达到显著性水平,对每一时间点(n=6 000)的数据(瞳孔大小和注视误差)与基线水平做t检验。由于这样在同一分析中多次使用t检验(多重比较,multi-comparison)可能会造成一定程度的虚假检测,即把原本不显著的结果错误地判定为显著,因此为消除由多重比较产生的虚假检测率(false discovery rate,FDR),对t检验的显著阈值在0.05水平上进行了校正(FDR0.05),即只有当某一时间点上t检验得到的P值小于矫正后的阈值时(P 图2 眨眼前后瞳孔面积的变化(较细的两条线表示标准误形成的包络;两条垂直实线间表示瞳孔值显著差别于基线的点(P 2.1眨眼对瞳孔反应的影响 由于瞳孔的绝对大小存在着很大的个体差异,以基线为标准对瞳孔值做归一化处理,并对所有22个被试在每个时间点上(n=6 000)求平均值。 分析发现,眨眼开始之前,瞳孔处于相对稳定的水平。但在眨眼结束之后,瞳孔反应出现了较大的波动。具体表现为: 眨眼结束后的200 ms内,瞳孔出现小幅度的放大(约2%),但未达到显著水平(P>FDR0.05)。在这之后,瞳孔发生大幅度的收缩,在眨眼结束后约700 ms达到最小值,并在眨眼结束后约4 000 ms恢复到基线水平,收缩最大幅度约12%。从图2中可以看出(浅灰色水平虚线),在眨眼结束后约300~1 700 ms间,被试间(n=22)的平均瞳孔值显著低于基线水平(P 2.2眨眼对注视位置的影响 图3(a)展示了22个被试平均注视误差的时间过程。在眨眼开始前,注视误差处于相对稳定的水平。但在临近眨眼开始前约30 ms内,注视误差快速增加到约0.7°,但未达到显著水平(P> FDR0.05)。这可能是有于伴随眨眼的眼动引起的(blink-associated eye movements),因为眨眼眼动会先于眨眼而发生[18]。在眨眼结束后,注视误差增加到1°左右,且显著地大于基线(P 图3 眨眼对注视位置的影响。(a)眨眼前后被试间(n=22)平均注视误差变化的时间过程(水平的虚线代表基线,在粗实线上下的两条细线代表标准误包络,眨眼结束到垂直细线之间的区域代表注视误差显著高于基线的点(P 由于光学眼动仪是根据计算得到的瞳孔中心和角膜反射的差值来确定注视的位置,因此,存在着一种无法避免的伪迹,即瞳孔大小的变化会引起由计算得到的瞳孔中心位置的改变,进而引起记录到的注视位置的偏移,而这种偏移实际上并不存在[19-20]。由于本研究发现在眨眼后瞳孔出现了大幅度的波动,因此也对由眨眼后瞳孔变化引起的注视偏移伪迹做出了详细的测量和分析。由于这种伪迹存在着很大的个体差异,因此,采用多人平均的分析方法很可能将这种伪迹淹没(见图3(a))。为了衡量眨眼后瞳孔变化和注视位置变化的相关程度,对每个被试计算了眨眼后瞳孔反应和注视误差的相关系数(200~4 000 ms)。分析表明,所有被试(n=22)相关系数绝对值的均值为 0.58±0.06(平均数±标准误),最大相关系数和最小相关系数(绝对值)分别为 0.95 和0.08。所有22个被试中,有19个(86%)表现为为正相关,3个(14%)为负相关,这说明眨眼后瞳孔的变化会引起记录到的注视位置的偏移,并且偏移的方向和程度因人而异。除了相关程度,也计算了注视位置随瞳孔变化的幅度。为此,对于眨眼后瞳孔面积和注视误差相关系数大于0.5(绝对值)的被试(n=15),计算了用眨眼后注视误差的最大值和最小值的差值来表示瞳孔变化对注视位置影响的幅度。计算结果表明,在15个相关系数大于0.5的被试中,这一幅度的均值为0.13°±0.01°,最大幅度和最小幅度分别为0.26°和0.04°。图3(b)展示了瞳孔变化对注视位置影响幅度最大的一名被试的结果,其幅度为0.26°,相关系数为0.91(P<0.000 1)。这再次说明,注视位置随瞳孔大小变化只是由于光学眼动仪在记录过程中产生的伪迹,并不是真正的注视位置的偏移。 3.1眨眼后瞳孔反应的生理机制以及在实验方法中的意义 本研究通过单一的长注视实验,排除了认知因素对眨眼和瞳孔反应的影响,发现眨眼对瞳孔反应可以产生持续约长达4 s的影响,并且眨眼后瞳孔的波动幅度高达12%。不同于以往的结果[10-11],发现了眨眼结束后小幅度的瞳孔放大,同时也发现了大幅度的瞳孔收缩。以往对瞳孔光反应的研究发现[1],当出现光亮刺激时,瞳孔会在刺激发生后200 ms左右产生收缩反应,并在500~1 000 ms达到最大收缩(瞳孔达到最小值)。这些研究结果和本研究发现的眨眼后的瞳孔反应在时程上十分一致,说明眨眼后的瞳孔收缩反应很可能是由于眨眼后瞳孔对眼内进光量增加(相对于眨眼过程中)的反应。但对于眨眼后瞳孔小幅度的扩大反应(约2%),其原因还尚不清楚。其中一种可能的解释是:小幅度的瞳孔扩张是眨眼中瞳孔暗反应过程的延续。因为在眨眼过程中,眼内进光量会大幅度减少,这会引起瞳孔的扩大。但发现眨眼结束后,瞳孔的面积低于基线水平,并在持续约200 ms的扩张反应后达到基线水平。并且,在眨眼过程中,由于眼睑对瞳孔的遮挡,无法获取到瞳孔的数据,因此,这种瞳孔暗反应的解释并不能被当前的实验结果证实。另一种可能的解释是:由于在眨眼的眼睑上升阶段(眼睛重新睁开),眼睑运动的加速度随时间而降低,因此在临近眨眼结束时,眼睑运动的速度是极低的。尽管本研究定义的眨眼结束时间点是在眨眼开始后瞳孔变化速度趋近于0的点(±0.5之间),但在这之后的短时间内,瞳孔反应仍然可能会受到眼睑运动的影响。因此,为更清楚地探究本研究发现的眨眼后短暂的瞳孔放大的机制,需要后续研究同时记录瞳孔反应和眼睑运动,并做更加精确的分析。 另外,更为重要的是,笔者的研究结果还可以为瞳孔反应的相关研究提供方法上的参考。由于在眨眼结束后,瞳孔仍然会发生持续的大幅度波动,因此在这些研究中,眨眼应被当做重要的干扰变量加以考虑。以往的大多数研究仅仅剔除了瞳孔丢失值或用插值的方法补充丢失的瞳孔数据,这并不能完全排除眨眼对实验结果造成的干扰。尤其是当眨眼本身与实验中考察的自变量密切相关时,例如在研究瞳孔反应与知觉状态切换的关系时[4, 11],眨眼可能对实验结果解释造成更大的混淆。 3.2眨眼对注视位置的影响 在研究中,发现眨眼会对注视位置造成偏差,尤其在很多研究中眨眼仅被定义为瞳孔丢失的区间时(图1中阴影部分,也是大部分眼动仪厂商提供的默认方法),这种偏差尤为明显(见图1)。即使本文采用了更为合理的眨眼检测方法,即以眼睑不再遮挡瞳孔作为眨眼结束的标记,在眨眼后依然存在1°以内、持续约200 ms的注视误差。虽然这样的误差相对较小,但对于一些对精度要求较高的研究来说,依然可能会对实验结果造成干扰。此外,在使用光学眼动仪记录注视位置时,眨眼后瞳孔的变化会引起注视位置的小幅度偏移。但这种偏移不只局限于眨眼后,任何可能会对瞳孔大小造成影响的因素,例如亮度变化以及认知加工负荷的变化等都会引起类似的注视位置偏移的伪迹。因此以注视位置为主要因变量的相关研究,应根据其精度要求来衡量眨眼以及上述其它因素对其实验结果的影响。 本课题旨在研究眨眼对瞳孔反应和注视位置的精确影响,为相关研究提供参考。通过记录并分析22个被试的瞳孔反应和注视位置的数据,作者发现,即使在眨眼结束后,眨眼对瞳孔和注视位置仍然存在着不可忽视的影响。眨眼对可以造成持续约4 s、最大幅度为12%的波动,并且会造成最大幅度约1°的注视位置的偏移。此外,对基于光学眼动仪的研究,由眨眼产生的瞳孔变化会引起记录到的注视位置的同步偏差,尽管其幅度微小,但仍然会对某些高精度的研究造成影响。因此,本研究认为,眨眼对瞳孔和注视位置的影响应当引起更多研究者的重视,并在结果解释中充分考虑到眨眼的影响。 [1] Beatty J, Lucero-Wagoner B. The pupillary system [J]. Handbook of Psychophysiology, 2000,2:142-162. [2] Nassar MR, Rumsey KM, Wilson RC,etal. Rational regulation of learning dynamics by pupil-linked arousal systems [J]. Nature Neuroscience, 2012,15(7):1040-1046. [3] Einhauser W, Koch C, Carter O. Pupil dilation betrays the timing of decisions [J]. Frontiers in Human Neuroscience, 2010,4:18. [4] Einhäuser W, Stout J, Koch C,etal. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry [J]. Proceedings of the National Academy of Sciences, 2008,105(5):1704-1709. [5] Sheliga B, Riggio L, Rizzolatti G. Orienting of attention and eye movements [J]. Experimental Brain Research, 1994,98(3):507-522. [6] Sheliga B, Riggio L, Rizzolatti G. Spatial attention and eye movements [J]. Experimental Brain Research, 1995,105(2):261-275. [7] Nakano T, Yamamoto Y, Kitajo K,etal. Synchronization of spontaneous eyeblinks while viewing video stories [J]. Proceedings of the Royal Society B: Biological Sciences, 2009,276: 3635-3544. [8] Collewijn H, Van der Steen J, Steinman R. Human eye movements associated with blinks and prolonged eyelid closure [J]. J Neurophysiol, 1985,54(1):11-27. [9] Evinger C, Manning KA, Pellegrini JJ,etal. Not looking while leaping: the linkage of blinking and saccadic gaze shifts [J]. Experimental Brain Research, 1994,100(2):337-344. [10] Fukuda K, Stern JA, Brown TB,etal. Cognition, blinks, eye-movements, and pupillary movements during performance of a running memory task [J]. Aviation, Space and Environmental Medicine, 2005, 76(Supplement 1):C75-C85. [11] Hupé JM, Lamirel C, Lorenceau J. Pupil dynamics during bistable motion perception [J]. Journal of Vision, 2009,9(7):10. [12] Siegle GJ, Ichikawa N, Steinhauer S. Blink before and after you think: blinks occur prior to and following cognitive load indexed by pupillary responses [J]. Psychophysiology, 2008,45(5):679-687. [13] Costela F, Otero-Millan J, McCamy M,etal. Microsaccades correct fixation errors due to blinks [J]. Journal of Vision, 2013,13(9):1335-1335. [14] Brainard DH. The psychophysics toolbox [J]. Spatial Vision, 1997,10:433-436. [15] Pelli DG. The VideoToolbox software for visual psychophysics: Transforming numbers into movies [J]. Spatial Vision, 1997,10(4):437-442. [16] Savitzky A, Golay MJ. Smoothing and differentiation of data by simplified least squares procedures [J]. Analytical Chemistry, 1964,36(8):1627-1639. [17] Genovese CR, Lazar NA, Nichols T. Thresholding of statistical maps in functional neuroimaging using the false discovery rate [J]. Neuroimage, 2002,15(4):870-878. [18] Bour L, Aramideh M, De Visser BO. Neurophysiological aspects of eye and eyelid movements during blinking in humans [J]. Journal of Neurophysiology, 2000,83(1):166-176. [19] Wyatt HJ. The human pupil and the use of video-based eyetrackers [J]. Vision Research, 2010,50(19):1982-1988. [20] Drewes J, Masson GS, Montagnini A. Shifts in reported gaze position due to changes in pupil size: Ground truth and compensation[C]//Proceedings of the Symposium on Eye Tracking Research and Applications. Santa Barbara: ACM, 2012: 209-212. The Influence of Spontaneous Blinks on the Pupil Response and Fixation Position Gao Xin Yang Kaifu Yan Hongmei Yao Dezhong#Li Yongjie* (School of Life Science and Technology, University of Electronic Science and Technology of China, Chengdu 610054, China) The pupil response and fixation position are important variables in eye movement researches. However, they are susceptive to the disturbance of eye blinks. While most studies only focus on the data missing and fluctuation during blinks, but neglect the more extensive influence of blinks on pupil response and fixation position. Thus, present study aims to precisely examine the impacts of blink on pupil response and fixation position. We recorded the pupil size and fixation position using a video eye tracker, and indentifying the blinks based on these data in a simple prolonged fixation task. The temporal sequence was relined to the onset and end of the detected blinks. We observed a large fluctuation (approximately 12%) in the pupil response, which last approximately 4 seconds after the ending of eye blink. During this period, the pupil size was significantly smaller than the baseline from 300 to 1700 ms (P blinks; fixation error; pupil; fixation position 10.3969/j.issn.0258-8021. 2015. 03.005 2014-12-20, 录用日期:2015-03-21 国家重点基础研究发展计划(973计划)(2013CB329401);国家自然科学基金(91420105,61375115) R339.14 A 0258-8021(2015) 03-0290-07 # 中国生物医学工程学会会员(Member, Chinese Society of Biomedical Engineering) *通信作者(Corresponding author), E-mail: liyj@uestc.edu.cn引言
1 研究方法
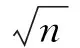
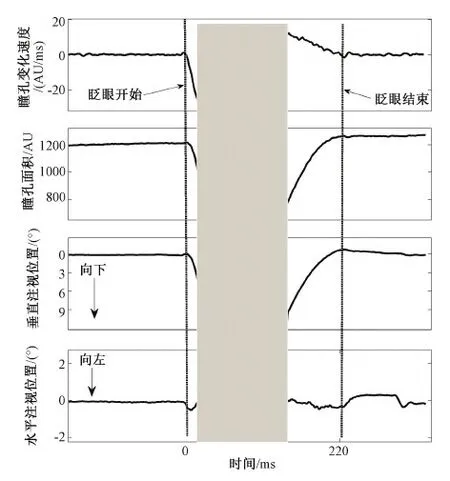
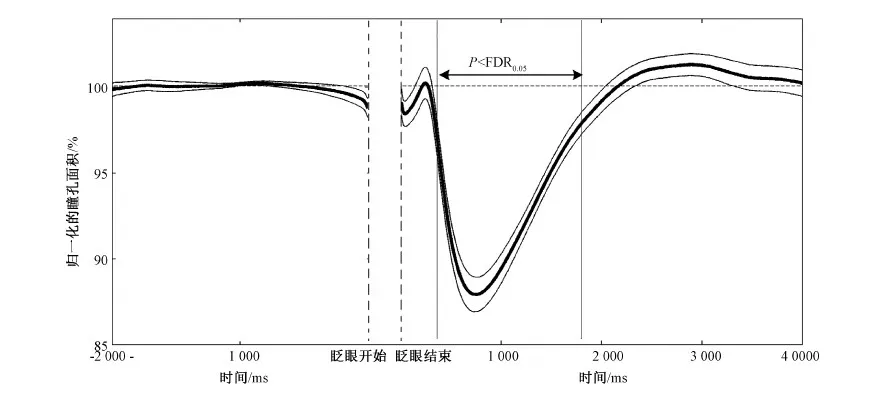
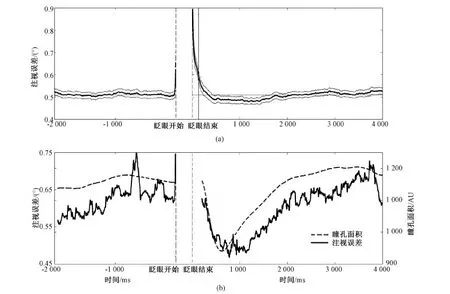
2 结果
3 讨论
4 结论
