高脂饮食诱导的肥胖对颞叶癫痫小鼠海马损伤的影响
2015-08-29邓子辉张金英李建华颜光涛
冯 杰,邓子辉,张金英,薛 辉,梁 辰,李建华,颜光涛
解放军总医院,北京 1008531基础所生化研究室;2生化科
高脂饮食诱导的肥胖对颞叶癫痫小鼠海马损伤的影响
冯 杰1,邓子辉1,张金英1,薛 辉1,梁 辰1,李建华1,颜光涛2
解放军总医院,北京 1008531基础所生化研究室;2生化科
目的 探讨肥胖对颞叶癫痫发生、发展的影响。方法 高脂饲料喂养C57BL/6J小鼠建立肥胖小鼠模型,建模成功后单侧海马微量注射200 ng海人酸(kainic acid,KA)诱导颞叶癫痫,记录小鼠癫痫评分,测定血清肌酸激酶同工酶(creatine kinase isoenzyme,CK-MB)浓度,Western blot检测海马S-100β的表达。海人酸注射5周后,观察小鼠海马病理结构改变。结果 海人酸注射后,肥胖小鼠的癫痫行为评分较野生型鼠显著增高;海人酸注射24 h后,与野生型鼠相比,肥胖小鼠海马血清CK-MB浓度和海马S-100β表达升高;海人酸注射后,病理切片显示肥胖鼠海马CA3区神经元丢失和星形胶质细胞活化较野生型鼠更严重,肥胖鼠星形胶质细胞形态发生明显变化。结论 肥胖可能加重了小鼠对海人酸诱导颞叶癫痫的损伤。
肥胖;海人酸;星形胶质细胞;颞叶癫痫;C57BL/6J小鼠
网络出版时间:2015-05-05 14:17 网络出版地址:http://www.cnki.net/kcms/detail/11.3275.R.20150505.1417.002.html
肥胖症是21世纪人类面临的最严峻的公共卫生问题之一,不良的生活方式和不健康的饮食习惯是目前导致肥胖的最主要原因。肥胖症既是一种疾病,也是其他疾病的重要诱因,包括常见的糖尿病、心血管疾病和神经系统疾病等[1-2]。流行病学调查发现,在难治型癫痫患者中超重和肥胖患者所占比例高,肥胖可能是影响癫痫病理过程的重要因素。癫痫是第二大神经系统性顽症,是由于神经元的同步异常放电引起的一种短暂脑功能障碍疾病。其中颞叶癫痫(temporal lobe epilepsy,TLE)是最常见的局灶性癫痫综合征,异常放电来源于颞叶[3-4]。临床病理研究发现,颞叶癫痫患者具有异常的海马形态,并且伴有特征的病理改变包括神经元的丢失和星形胶质细胞活化,尤其是海马CA3区[5-6]。目前,C57BL/6J小鼠单侧海马微量注射海人酸是常见的颞叶癫痫的动物模型建模方法[7]。本研究通过高脂饲料喂养C57BL/6J小鼠建立肥胖模型[8],然后注射海人酸诱导颞叶癫痫小鼠模型,检测小鼠行为学、病理学上的变化,初步探讨肥胖与颞叶癫痫的关系。
材料和方法
1 动物和试剂 C57BL/6J小鼠(SPF级),90只,4周龄,雄性,体质量14 ~ 15 g,购自解放军总医院医学实验动物中心;标准饲料(10% kJ%fat,北京中兴饲料);高脂饲料(40% kJ%fat,上海实验动物中心);海人酸(Sigma Chemical Co. St Louis,MO,USA);脑立体定位仪(ZS-B/C,北京众实迪创科技发展有限责任公司);5 μl微量进样器Microliter Syringes(上海高鸽工贸有限公司);乙醚(北京益利精细化学品公司);甲苯胺蓝(Sigma Chemical Co. St Louis,MO,USA);一抗:胶质纤维酸性蛋白(GFAP,Cell Signaling Technology Inc. #3670),S-100β(Santa Cruz Biotechnology,SC-28533);二抗:山羊抗小鼠IgG/辣根酶标记、山羊抗兔IgG/辣根酶标记(北京中衫金桥公司);柠檬酸盐缓冲液(北京普利莱公司);DAB显色液(北京中衫金桥公司)。
2 高脂饮食诱导肥胖小鼠模型的建立 C57BL/6J饲养于无特定病原体动物房,温度22 ~ 24℃,维持12 h昼夜交替。小鼠用标准饲料适应性喂养1周,1周后称体质量。小鼠按随机数字表法分为2组,肥胖组给予高脂饲料饲养,标准饲料饲养组给予标准饲料饲养,均自由饮水和进食。体质量记录:称质量前1 d于下午18:00禁食,第2天9:00称量。饲养12周后,肥胖组小鼠体质量高于野生型组小鼠20%以上,肥胖小鼠模型构建成功。
3 颞叶癫痫小鼠模型的建立 将肥胖小鼠分为氯化钠注射组(OS组,15只)和海人酸注射组(OK组,30只),野生型小鼠分为氯化钠注射组(WS组,15只)和海人酸注射组(WK组,30只)。将小鼠乙醚麻醉后,置于脑立体定位仪上,以大脑Bregma为原点的海马定位注射坐标:-2.0、-1.8、-2.3(右侧海马),进行单侧注射海人酸200 ng(100 ng/μl,2 μl)后,速度2 μl/5 min,注射结束留注射器5 min,去除吸入麻醉机后,观察行为表现。氯化钠组以0.9%氯化钠注射液代替海人酸,其余步骤和注射方式同海人酸小鼠。
4 行为学观察 海人酸注射后,根据Racine分级标准[9]进行痫性评分:0分:无惊厥;1分:咀嚼运动、眨眼、胡须颤抖等面部肌肉抽搐;2分:颈部肌肉抽搐,节律点头;3分:单侧前肢震颤,颈部肌肉痉挛;4分:双前肢阵挛,后肢站立;5分:身体持续性强直,跌倒伴阵挛。
5 血清肌酸激酶同工酶的测定 海人酸注射24 h,小鼠麻醉后,摘眼球取血,静置4℃过夜,3 000 r/ min离心15 min后取上清液,采用比色法,通过Roche Cobas 8000 Modular Analyser仪器测定。
6 尼式染色 4 μm石蜡切片脱蜡至水,用磷酸盐缓冲溶液洗2 min,放1%甲苯胺蓝溶液中室温20 min,用梯度乙醇依次脱水,结果尼式小体呈深蓝色颗粒,细胞核呈淡蓝色,背景基本无色。
7 免疫组化法观察GFAP的表达和形态变化4 μm石蜡切片脱蜡至水,柠檬酸盐缓冲溶液高压修复5 min,37℃ 5%的山羊血清封闭20 min,GFAP(1:400) 4℃过夜,漂洗后,加辣根过氧化物酶标记的二抗37℃孵育40 min,DAB显示,苏木素染核,漂洗后封片,置于显微镜下观察并拍照,GFAP显黄色,细胞核显淡蓝色。
8 Western blot检测S-100β表达 小鼠处死后,取脑剥离右侧海马,放入RIPA蛋白裂解液,匀质离心提取总蛋白,BCA法蛋白定量。蛋白上样量为100 μg,在质量分数为12%的SDS-PAGE上进行垂直电泳,然后电转移到硝酸纤维素膜上(100 V,80 min),将硝酸纤维素膜放入含5%脱脂奶粉(TBST溶解)塑料袋中,放入摇床37℃封闭1 h,将稀释好的一抗,S-100β(1∶1 000)、β-actin(1∶500),4℃封闭过夜。取出后放室温30 min,TBS/T漂洗膜3次×10 min,加入稀释好的二抗(1∶3 000),37℃孵育30 min,TBS/T漂洗硝酸纤维素膜3次×10 min,加入发光液,暗室曝光。扫描后,使用Image Plus 6.0软件,分析其灰度值,3次独立实验求平均值。
9 统计学方法 采用GraphPad Prism软件进行统计分析,数据以±s表示,组间比较用t检验,与对照组比较采用LSD-t检验。P<0.05为差异有统计学意义。
结 果
1 高脂饮食诱导的肥胖模型建立 经过12周喂养,高脂饲料饲养组小鼠体质量比标准饲料饲养组高出20%以上(P<0.05),高脂饲料喂养的小鼠脂肪垫湿重是标准饲料喂养小鼠的2倍以上,肥胖小鼠模型建立成功(图1)。
2 颞叶癫痫肥胖小鼠行为学评分和血清肌酸激酶同工酶(creatine kinase isoenzyme,CK-MB)增高海人酸注射后4 h内,肥胖组每只小鼠Racine最高评分平均值显著高于野生型小鼠(P<0.05),OK组每只小鼠最高评分均值为4.6,WK组每只小鼠最高评分均值为3.2(图2A)。海人酸注射后,小鼠血清内肌酸激酶同工酶显著增加(P<0.05),肥胖鼠和标准饲料喂养鼠差异无统计学意义,但肥胖组血清肌酸激酶同工酶的平均浓度高于标准饲料喂养组(P>0.05)(图2B)。
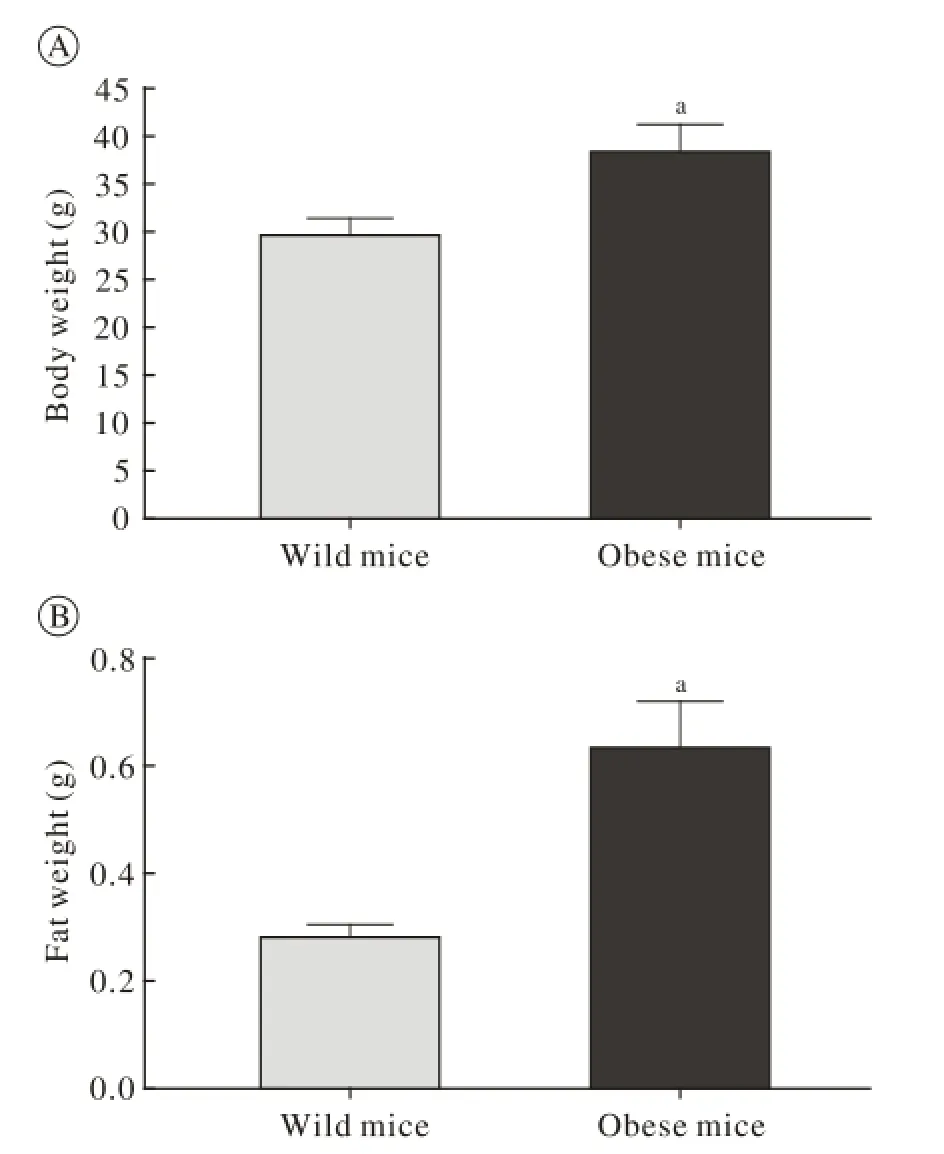
图 1 肥胖模型的建立(aP<0.05, vs wild mice group)Fig. 1 Establishment of obesity model (aP<0.01, vs wild mice group)
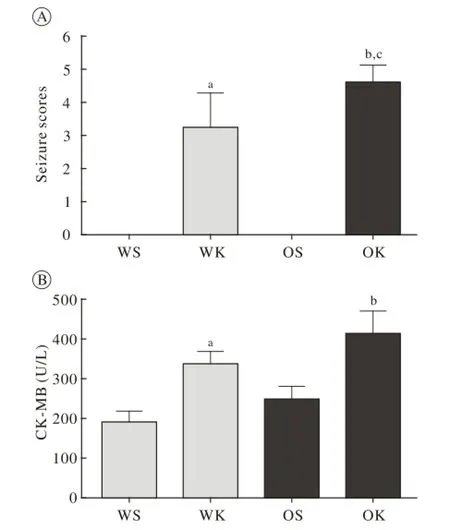
图 2 肥胖对海人酸诱导的颞叶癫痫小鼠抽搐评分与血清肌酸激酶同工酶的影响(aP<0.05, vs WS group;bP<0.05, vs OS group;cP<0.05, vs WK group)Fig. 2 Effect of obesity on seizure scores and serum CK-MB after KA-induced mice model of TLE (aP<0.05, vs WS group;bP<0.05, vs OS group;cP<0.05, vs WK group)
3 颞叶癫痫肥胖小鼠海马CA3区神经元丢失严重及星形胶质细胞过度活化 海人酸注射后继续饲养5周,小鼠脑冠状面切片尼式染色结果显示,与野生型小鼠海马注射侧CA3区相比,肥胖组小鼠神经元丢失更重,神经元弥散(图3A)。免疫组化结果显示,星形胶质细胞过度活化和星形胶质细胞形态改变(星形胶质细胞胞体变粗,细胞足加长,细胞聚集成簇)(图3B)。颞叶癫痫肥胖小鼠海马S-100β(活化星形胶质细胞分泌蛋白)表达升高(图4)。
讨 论
肥胖患者多为单纯性肥胖,占总肥胖人数90%以上,主要由不健康的饮食习惯和缺乏锻炼引起。本研究采用高脂饮食诱导方式建立肥胖模型很好的模仿单纯性肥胖,避免使用基因缺陷鼠如ob/ob或db/db,基因缺陷鼠会产生严重的代谢紊乱[10]。肥胖是高血压、冠心病、心肌梗死、糖尿病、脑梗死、阿尔茨海默病等多种疾病的主要危险因素。目前观点认为,癫痫患者发生体质量增加是抗癫痫药物如丙戊酸钠的不良反应[11-13],但流行病学调查发现,在未经药物治疗的难治性癫痫中肥胖者患病率高于正常体质量者[14-15],本文探究肥胖是否加重颞叶癫痫。
颞叶癫痫发病率高,治疗效果不佳,严重影响病人生活和工作。目前认为,神经元离子通道改变、突触传递异常、神经元微环境发生变化导致兴奋性与抑制性的不平衡,从而导致神经元异常同步放电[16-17]。本研究探讨肥胖与癫痫的关系可能会为颞叶癫痫发生、发展的研究提供一个新思路。病理研究发现,颞叶癫痫大部分由颞叶海马组织的变形引起的,神经元丢失和星形胶质细胞活化是海马变形的主要特征,因此本研究主要观察小鼠注射海人酸后海马组织的病理变化。研究发现,在颞叶小鼠模型中,肥胖小鼠的行为学Racine评分要明显高于野生型小鼠。研究表明,全身痉挛发作可引起血清肌酸激酶同工酶升高[18],肥胖小鼠血清肌酸激酶同工酶浓度平均值高于标准饲料饲养组小鼠,但无统计学差异,可能与小鼠数量少有关。病理切片显示,与野生型小鼠相比,肥胖小鼠神经元丢失和扩散加重;免疫组化显示,星形胶质细胞过度活化以及星形胶质细胞胞体加粗,星形胶质细胞足加长,细胞聚集。S100β是活化星形胶质细胞的分泌蛋白,在唐氏综合征、阿尔兹海默病、癫痫中都过表达,是判断脑损伤的常用指标[19],在肥胖鼠海马中大量表达,表明星形胶质细胞过度活化。活化的星形胶质细胞可以迁移聚集在损伤部位形成瘢痕,阻碍了营养物质向神经元的运输,同时活化的星形胶质细胞兴奋性氨基酸转运受体和谷氨酸合成酶活力降低,导致神经元外兴奋性氨基酸浓度升高,最终影响神经元的微环境[20],与癫痫的反复发作密切相关。因此,肥胖可能促进了颞叶癫痫的发生、发展。肥胖小鼠海马中神经元的大量丢失和星形胶质细胞的过度活化可能与肥胖小鼠血清相关因子变化有关,研究表明,肥胖小鼠具有高瘦素水平,JAK2/ STAT3是瘦素主要下游信号通路,JAK2/STAT3信号通路的激活可以促进星形胶质细胞的活化,星形胶质细胞活化可以促进癫痫的发生[21-23]。提示肥胖的颞叶癫痫患者减轻体质量可能有利于疾病的治疗,同时对了解颞叶癫痫的机制提供了一个新思路。因此,肥胖影响颞叶癫痫的机制值得进一步研究。
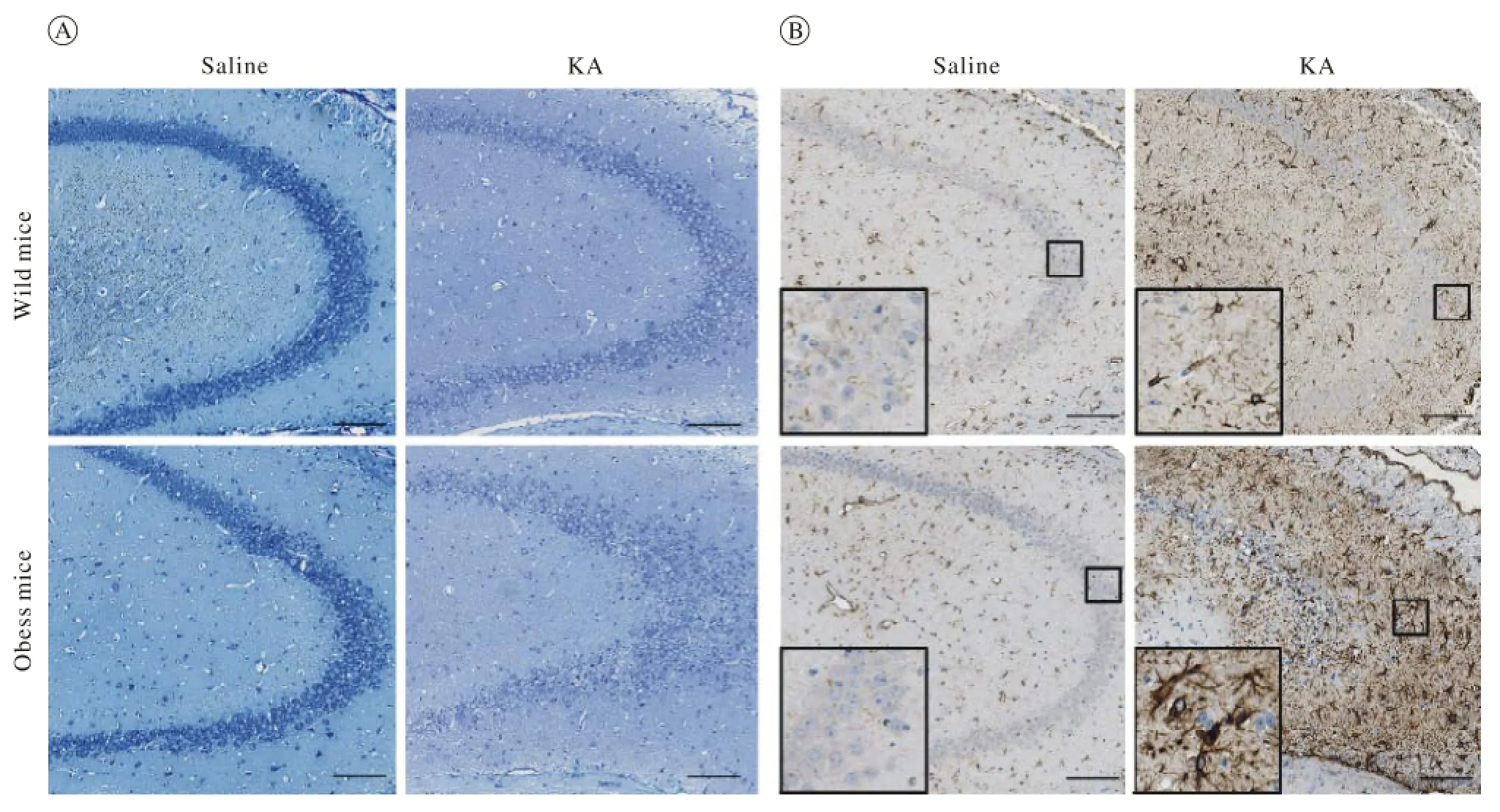
图 3 肥胖对海人酸诱导的颞叶癫痫小鼠海马CA3区神经元(A)和星形胶质细胞的影响(B)Fig. 3 Effect of obesity on neurons and astrocytes in hippocampal CA3 after KA-induced mice model of TLE

图 4 肥胖对海人酸诱导颞叶癫痫小鼠海马S-100β表达的影响Fig. 4 Effect of obesity on hippocampal S-100βexpression after KA-induced mice model of TLE (aP<0.05, vs WS group,bP<0.05, vs OS group,cP<0.05, vs WK group)
1 Spielman LJ, Little JP, Klegeris A. Inflammation and insulin/IGF-1 resistance as the possible Link between obesity and neurodegeneration[J]. J Neuroimmunol, 2014, 273(1/2): 8-21.
2 Jung UJ, Choi MS. Obesity and its metabolic complications: the role of adipokines and the relationship between obesity, inflammation,insulin resistance, dyslipidemia and nonalcoholic fatty liver disease[J]. Int J Mol Sci, 2014, 15(4):6184-6223.
3 Arabadzisz D, Antal K, Parpan F, et al. Epileptogenesis and chronic seizures in a mouse model of temporal lobe epilepsy are associated with distinct EEG patterns and selective neurochemical alterations in the contralateral hippocampus[J]. Exp Neurol, 2005, 194(1):76-90.
4 Jan MM, Sadler M, Rahey SR. Electroencephalographic features of temporal lobe epilepsy[J]. Can J Neurol Sci, 2010, 37(4):439-448.
5 Tang Y, Yu X, Zhou B, et al. Short-term cognitive changes after surgery in patients with unilateral mesial temporal lobe epilepsy associated with hippocampal sclerosis[J]. J Clin Neurosci, 2014,21(8):1413-1418.
6 Van Der Hel WS, Notenboom RG, Bos IW, et al. Reduced glutamine synthetase in hippocampal areas with neuron loss in temporal lobe epilepsy[J]. Neurology, 2005, 64(2): 326-333.
7 Sha LZ, Xing XL, Zhang D, et al. Mapping the spatio-temporal pattern of the mammalian target of rapamycin (mTOR) activation in temporal lobe epilepsy[J]. PLoS One, 2012, 7(6):e39152.
8 Murase T, Nagasawa A, Suzuki J, et al. Beneficial effects of tea catechins on diet-induced obesity: stimulation of lipid catabolism in the liver[J]. Int J Obes Relat Metab Disord, 2002, 26(11):1459-1464.
9 Racine RJ, Gartner JG, Burnham WM. Epileptiform activity and neural plasticity in limbic structures[J]. Brain Res, 1972, 47(1):262-268.
10 Pelleymounter MA, Cullen MJ, Baker MB, et al. Effects of the obese gene product on body weight regulation in ob/ob mice[J]. Science,1995, 269(5223): 540-543.
11 Verrotti A, La Torre R, Trotta D, et al. Valproate-induced insulin resistance and obesity in children[J]. Horm Res, 2009, 71(3):125-131.
12 Kanemura H, Sano F, Maeda Y, et al. Valproate sodium enhances body weight gain in patients with childhood epilepsy: a pathogenic mechanisms and open-label clinical trial of behavior therapy[J]. Seizure, 2012, 21(7):496-500.
13 Ayyagari M, Chitela SR, Kolachana V. Obesity, polycystic ovarian syndrome and thyroid dysfunction in women with epilepsy[J]. Ann Indian Acad Neurol, 2012, 15(2): 101-105.
14 Daniels ZS, Nick TG, Liu C, et al. Obesity is a common comorbidity for pediatric patients with untreated, newly diagnosed epilepsy[J]. Neurology, 2009, 73(9): 658-664.
15 Janousek J, Barber A, Goldman L, et al. Obesity in adults with epilepsy[J]. Epilepsy Behav, 2013, 28(3): 391-394.
16 Gorter JA, Zurolo E, Iyer A, et al. Induction of sodium channel Na(x)(SCN7A) expression in rat and human hippocampus in temporal lobe epilepsy[J]. Epilepsia, 2010, 51(9):1791-1800.
17 Soukupová M, Binaschi A, Falcicchia C, et al. Impairment of GABA release in the hippocampus at the time of the first spontaneous seizure in the pilocarpine model of temporal lobe epilepsy[J]. Exp Neurol, 2014, 257:39-49.
18 张亚东,郝苏怡,李洪亮.癫痫全身强直阵挛发作与自动症发作肌酶变化分析[J].神经损伤与功能重建,2014,03(3):251-252.
19 Yang L, Li FH, Zhang HJ, et al. Astrocyte activation and memory impairment in the repetitive febrile seizures model[J]. Epilepsy Res, 2009, 86(2/3): 209-220.
20 Eid T, Tu N, Lee T, et al. Regulation of astrocyte glutamine synthetase in epilepsy[J]. Neurochem Int, 2013, 63(7): 670-681.
21 Lynch JJ, Shek EW, Castagné V, et al. The proconvulsant effects of leptin on glutamate receptor-mediated seizures in mice[J]. Brain Res Bull, 2010, 82(1/2): 99-103.
22 Xu Z, Xue T, Zhang Z, et al. Role of signal transducer and activator of transcription-3 in up-regulation of GFAP after epilepsy[J]. Neurochem Res, 2011, 36(12): 2208-2215.
23 Koga S, Kojima A, Ishikawa C, et al. Effects of diet-induced obesity and voluntary exercise in a tauopathy mouse model: implications of persistent hyperleptinemia and enhanced astrocytic leptin receptor expression[J]. Neurobiol Dis, 2014, 71:180-192.
Effect of high fat diet induced-obesity on hippocampal damage of temporal lobe epilepsy in mice
FENG Jie1, DENG Zihui1, ZHANG Jinying1, XUE Hui1, LIANG Chen1, LI Jianhua1, YAN Guangtao2
1Bichemistry Laboratory of Basic Medical Institute;2Department of Clinic Biochemistry Chinese PLA General Hospital, Beijing 100853, China
Corresponding author: YAN Guangtao. Email: yan301@263.net
Objective To study the effect of obesity in pathogenesis of temporal lobe epilepsy. Methods A high-fat diet induced obesity model of C57BL/6J mice was established and then a normal dose of kainic acid (KA, 200 ng per mouse) were microinjected into the hippocampus of mice to induce temporal lobe epilepsy and seizures were scored with the Racine scores. The concentration of serum CK-MB was tested and the expression of S-100β was measured by Western blot. Then, the hippocampal pathogenesis of mice was observed 5 weeks after KA injection. Results The mean peak seizure scores of obese mice significantly increased compared with the wild mice after KA injection. After 24 h of injection of KA, the mean serum CK-MB concentration and hippocampal S-100βexpression of obese mice were higher than wild mice. The neuron loss and astrocyte activation significantly increased in hippocampal CA3 region of obese mice compared with these of the wild mice after KA injection, there were significantly morphologic change of reactive astrocyte in obese mice. Conclusion Obesity may be detrimental to KA-induced brain injury.
obesity; kainic acid; astrocyte; temporal lobe epilepsy; C57BL/6J mice
R 363.2
A
2095-5227(2015)07-0724-05
10.3969/j.issn.2095-5227.2015.07.023
2014-11-20
科技基础性工作专项项目(2011FY130100);国家科技支撑计划项目(2012BAK25B01)
Supported by National Basic Research Development Program of the Ministry of Science and Technology of China (2011FY130100);The National Key Technology R&D Program (2012BAK25B01)
冯杰,男,在读硕士。研究方向:神经退行性疾病的机制。Email: fengjie654321@126.com
颜光涛,男,研究员,主任,博士生导师。Email: yan3 01@263.net
