ARMARM结构的缺失对SPAG6SPAG6在哺乳细胞中定位的影响*
2015-08-04汪智琼张玲石玉琴李红飞张志兵武汉科技大学医学院公共卫生学院武汉430065华中科技大学同济医学院附属同济医院VirginiaCommonwealthUniversityRichmondVA
汪智琼张 玲石玉琴李红飞张志兵, 3**.武汉科技大学医学院公共卫生学院(武汉 430065) ;2.华中科技大学同济医学院附属同济医院; 3. Virginia Commonwealth University, Richmond, VA
·论 著·
ARMARM结构的缺失对SPAG6SPAG6在哺乳细胞中定位的影响*
汪智琼1, 2张 玲1石玉琴1李红飞1张志兵1, 3**
1.武汉科技大学医学院公共卫生学院(武汉 430065) ;2.华中科技大学同济医学院附属同济医院; 3. Virginia Commonwealth University, Richmond, VA
摘要目的 本研究构建精子相关抗原6(SPAG6)基因全长及6个不同长短ARM序列的真核表达载体pEGFPN2-SPAG6-△ARM,并使其在CHO细胞中表达,观察全长及6个ARM结构缺失后对真核细胞CHO细胞中SPAG6蛋白定位的影响。方法 利用NCBI数据库,寻找小鼠SPAG6蛋白的保守功能区,分析全长蛋白序列,检索出SPAG6有7个ARM区域。利用PCR技术扩增出6个缺失不同ARM结构的SPAG6 cDNA,测序后亚克隆至携带绿色荧光蛋白基因的pEGFP-N2真核表达载体中,对阳性克隆进行酶切和测序鉴定,将构建的重组质粒转染到CHO细胞中,分别提取细胞蛋白进行Western blot检测。利用共聚焦激光扫描显微镜观察7个不同SPAG6/GFP融合蛋白在CHO细胞内的定位。结果酶切和测序鉴定表明,全长及6个缺失不同长短ARM结构的真核表达质粒构建成功,转染实验发现重组质粒均能够在CHO细胞中表达,但仅全长表达产物定位于微管,缺失任何一个ARM区域都可影响在细胞中的微管定位。结论SPAG6在哺乳动物细胞内的正确定位依赖于全长。该研究为进一步研究SPAG6蛋白的结构与功能奠定基础。
关键词精子相关抗原6基因; ARM结构域; 不育, 男性
Key woorrddss Sperm- associated antigen 6; armadillo domains; infertility, male
男性不育是临床上常见的一种疾病,随着社会现代化进程的发展,空气质量、环境卫生、食品安全及个人婚育观等问题的不断凸显,近年来发病率有急剧上升趋势。全球目前有10%~15% 的育龄夫妇有生育障碍,其中20%~40%为男性,绝大部分是因为精子功能缺陷而导致不育[1]。精子发生障碍是导致男性不育的一个重要因素,其中遗传因素缺陷是一个重要机制[2-4]。 正常精子的生成是一个极为复杂的生理过程,许多基因参与这一过程[5],近年来随着小鼠基因敲除技术的广泛运用,越来越多调节精子发生的基因被发现[6],据报道目前有超过2 300个基因参与[7]。其中精子相关抗原6(Sperm- associated antigen 6,PAG6)是一个重要基因[8]。
SPAG6是最初使用患不育症的男性患者的高滴度抗体血清筛选睾丸基因表达文库而克隆出来的[9]。研究发现,SPAG6基因表达蛋白是精子鞭毛支架蛋白的一种,对维持精子鞭毛结构完整和精子鞭毛活动有重要作用。SPAG6基因缺失或沉默时,动物表现为精子活力损伤、精子畸形和雄性不育[10]。经SMART分析软件得到小鼠SPAG6蛋白由7个armadillo(ARM)区域组成,ARM可介导蛋白-蛋白相互作用从而可能影响蛋白在细胞中的定位及功能。为研究ARM序列对SPAG6定位及蛋白的功能,在构建的含全长SPAG6绿色荧光素报告基因的基础上,我们构建了6个携带不同ARM序列的SPAG6绿色荧光蛋白基因的真核表达载体,并在CHO细胞中表达定位,观察其与完整PAG6基因表达产物在细胞内定位的差异,探讨某一段ARM区域缺失影响了SPAG6在体外细胞中的定位,为进一步探讨SPAG6的生物学效应奠定基础。
材料与方法
一、菌株﹑细胞﹑质粒及主要试剂
小鼠SPAG6/pEGFP-N2 质粒、CHO细胞为本实验室保存;大杆菌肠DH5a感受态细胞、质粒小提试剂盒及TOP10感受态细胞购自TIANGEN公司;CR试剂盒购自Fermentas公司;限制性核酸内切酶coRI和BamHI购自Thermo公司;ECL试剂盒,TA克隆试剂盒购自Invitrogen公司;BCA蛋白测定试剂盒、T4DNA连接酶购自碧云天公司;DNA marker、rotein marker购自东盛生物科技有限公司;DNA凝胶回收试剂盒购自AXYGEN公司;胎牛血清购自杭州四季青公司;DMEM培养液及胰酶购自HYclone公司;FuGene®HD Transfection Reagent购自Roche公司;anti-ɑ-GFP抗体购自博奥森公司;抗actin抗体、抗HRP标记的羊抗鼠IgG购自飞羿科技有限公司;引物合成及DNA序列测定由上海生工生物工程有限公司完成;其他试剂为国产分析纯。
二、小鼠SPAG6SPAG6系列缺失ARMARM结构cDNAcDNA的扩增
以小鼠全长SPAG6/pEGFP-N2质粒为模板,用下列引物扩增出系列缺失ARM的SPAG6 cDNA,检索出SPAG6有7个重复的ARM保守结构区域,进行逐步缺失处理,设计扩增的PCR前后引物,并在引物中加入EcoR I 和BamH I 两个限制性酶切位点。
共用的上游引物5'-GAATTCAATGAGCCAGCGG CAGGTGCTGCAAG-3'
②缺ARM下游引物5'-GGATCCGAGCAGCCAC GTAACCCAGCATCATG-3'
③缺ARM下游引物5'-GGATCCGTTTTGCAATC TCTCTGATTAAGGTG-3'
④缺ARM下游引物5'-GGATCCGCTTTGCAATC TGACTGAGAGCTGAG-3'
⑤缺ARM下游引物5'-GGATCCGCTTTGAGATA TCACTGAGGGCCGAG-3'
⑥缺ARM下游引物5'-GGATCCGGTGTCTTGCA ATGTATCCGAGTGC-3'
⑦缺ARM下游引物5'-GGATCCGGTAATTGGCC AGTCTCCCCAGAGC-3'
(一)目的基因片段的扩增及回收
PCR扩增,将PCR产物进行1%琼脂糖凝胶电泳,紫外灯下切割含有目的条带的凝胶块,利用凝胶回收试剂盒进行目的片段回收。
(二)TA克隆及鉴定
将回收产物和PCR 2.1载体在T4DNA连接酶作用下进行TA克隆,4℃连接过夜。取2μl TA clone产物转化至50μl TOP10感受态细胞中,涂布于含氨苄青霉素的LB培养基上,每板预处理涂有IPTG(异丙基-β-D-硫代吡喃半乳糖苷) 40μl(1 mol/L)和X-gal 80μl(20 mg/mL),37℃过夜培养。观察培养基上有白蓝菌落生长,挑取白色单克隆菌落适量扩增后提质粒DNA,经EcoRⅠ和BamH Ⅰ限制性核酸内切酶双酶切后,电泳观察酶切结果。
(三)重组质粒测序
将阳性克隆质粒送公司进行测序,获得6段缺失ARM区域编码序列。
三、SPAG6/pEGFP-N2FP-N2表达载体的构建
将pEGFP-N2载体和6个系列缺失ARM的SPAG6/ PCR2.1 TA克隆质粒分别用EcoRⅠ和 BamHⅠ双酶切后,凝胶回收目的片段,将两个片段用T4DNA连接酶4℃连接过夜,取5μl连接产物转化到DH-5ɑ大肠杆菌感受态中,37℃培养过夜。挑取单克隆菌落,接种于每毫升培养基含有5μl卡那霉素的LB培养基中,37℃震荡过夜。用质粒小提试剂盒提取质粒DNA,用EcoRⅠ和BamHⅠ双酶切鉴定外源基因的插入,阳性质粒送公司进一步测序分析。
四、SPAG6/pEGFP-N2FP-N2转染哺乳细胞
CHO细胞用DMEM高糖培养基培养,细胞铺于六孔板至70%~80%融合,按Roche公司提供的FuGene®HD Transfection Reagent转染六孔板和chambered slides的说明书进行操作。
五、蛋白质提取与Western Blot Blot鉴定
转染48h后,弃掉培养基,用PBS清洗两遍。向6孔板中的每个板中加入200μl含有蛋白酶抑制剂的RIPA裂解液,用无菌枪头缓慢吹打细胞数次。收集所有液体到新的离心管中,冰上放置30min,裂解细胞。13 000×g,4℃离心10min,将上清转到新的离心管中,取上清5μl进行定量。
用BCA蛋白测定试剂盒将蛋白定量后,取40μg总蛋白经10% SDS-PAGE凝胶分离,电泳转膜至NC膜上,5%脱脂奶粉封闭2h,TBST洗膜15min,3次。用anti-GFP抗体(1:500稀释),4℃过夜,TBST洗膜3次。再用辣根过氧化物酶标记的羊抗鼠(1:5000稀释)二抗孵育1h,TBST洗膜15min,3次。ECL显色,压片。
六、共聚焦激光扫描显微镜观察SPAG6/GFP6/GFP融合蛋白在细胞内定位
将构建的绿色荧光蛋白表达载体SPAG6/ pEGFP-N2转染到CHO细胞中48h后,应用OLYMPUS FV 1000S-SIM/IX81激光共聚焦扫描显微镜进行扫描,激发光波长为488nm。
结 果
一、小鼠SPAG6SPAG6由7个ARMARM序列组成
为寻找小鼠SPAG6蛋白的保守功能区,用SMART软件分析全长蛋白序列, 发现SPAG6有7个ARM区域,结果见图1。
二、6个缺失ARMARM结构的小鼠SPAG6 cDNA cDNA的扩增
为研究ARM区域对SPAG6定位的影响,以全长SPAG6/pEGFP-N2 质粒为模板,6对特异性引物进行聚合酶连反应扩增6个缺失ARM结构基因。扩增的条件为95℃5min预变性;以95℃ 30s,60℃ 45s,72℃ 90s这3个温度进行17个循环;最后72℃10min,温度降至4℃。PCR产物经1%水平琼脂糖凝胶电泳、溴化乙锭染色后,在凝胶成像仪下照相观察结果(图2)。

图1 蛋白序列示意图及各段ARM区域对应的氨基酸起点和终点
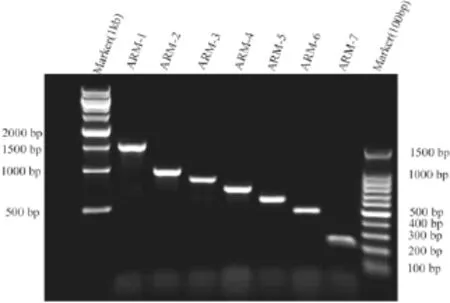
图2 PCR CR扩增出的SPAG6SPAG6的全长及6个缺失ARMARM结构的基因序列
三、SPAG6SPAG6全长及缺失不同长短ARMARM区域的SPAG6真核表达质粒的构建和鉴定
将重组质粒SPAG6/pEGFP-N2和pEGFP-N2质粒经EcoRⅠ和BamHⅠ进行限制性核酸内切酶酶切后得到了约4.7kb的pEGFP-N2和不同分子量的7条Spag6条带,条带在相应的分子量处(图3)。

图3 对重组融合质粒DNADNA,行EcoRⅠEcoR 和BamHⅠBamH 限制性核酸内切酶双酶切
四、Western blot blot检测7个不同SPAG6/GFP6/GFP融合蛋白在CHOCHO细胞中的表达
将构建的7个真核表达质粒SPAG6/pEGFP-N2转染到CHO细胞中,48h以后将提取的蛋白经10%SDSAGE凝胶电泳,Western blot检测到了融合绿色荧光蛋白GFP的7个SPAG6蛋白表达(图4)。
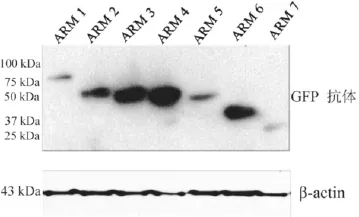
图4 对融合蛋白SPAG6/GFP6/GFP的表达进行Western blot lot 检测
五、利用共聚焦激光扫描显微镜观察7个不同PAG6/GFP6/GFP融合蛋白在CHOCHO细胞内的定位
将构建的7个带GFP的真核表达质粒转染到CHO细胞中,48h以后通过共聚焦激光扫描显微镜观察PAG6/GFP在细胞内定位。观察7个SPAG6/GFP融合蛋白在细胞内定位,由图5可看出,仅全长SPAG6在微管中表达,缺失最C端的一个或两个ARM结构的PAG6,即实验命名的ARM2和ARM3表达定位于胞浆,当从C端更多的缺失其他ARM结构的SPAG6,即实验命名的ARM4-7在整个细胞中表达。
讨 论
SPAG6是双鞭毛衣藻PF16(paralysed fl agella 16)的同源体,该同源蛋白衣藻PF16位于鞭毛中央轴突的中心管内参与调节鞭毛的运动[11, 12]。小鼠SPAG6不仅存在于精子鞭毛,在具有运动纤毛的组织,如肺、脑室膜中也有表达,在运动纤毛内,SPAG6定位于调节纤毛运动的中心轴突上,并且决定其他蛋白,如SPAG16L、SPAG17在中心轴突的定位[13, 14]。 SPAG6基因敲除小鼠模型已经建立,SPAG6缺失小鼠由于运动纤毛功能障碍而表现出雄性不育。此外,近一半SPAG6缺失小鼠生长较正常小鼠显著减慢,提示SPAG6可能和细胞生长有关,最近研究表明SPAG6在肺癌等肿瘤组织中表达显著增加,表明该基因跟肿瘤形成有关[15]。
过去的研究表明,在体外转染的哺乳细胞中,SPAG6定位于微管系统[16],但其微管定位的结构基础并不清楚。微管是真核细胞普遍存在的一种纤维结构,由微管蛋白和少量微管结合蛋白的聚合作用形成,是细胞骨架的主要成分之一[17]。微管在体内起着十分重要的作用,主要参与细胞运动、细胞器的定位、胞内物质的运输、真核细胞的有丝分裂过程及细胞信号传递等[18-22]。SPAG6定位于微管,该基因可能也参与微管的一些相关作用,为进一步了解其功能,探讨其微管定位的结构基础是非常有必要的。
SPAG6蛋白全长序列中有7个ARM序列。ARM重复序列是介导蛋白和蛋白之间相互作用的一种重要结构域,每个ARM区域都是由42个氨基酸构成,该氨基酸重复序列的特点是高度保守,并且仅有右手超螺旋结构,普遍存在于真核细胞中,相关研究发现ARM区域参与生物的多种重要生理过程,例如参与了细胞信号的转导、调节胞质内的物质转换、协助细胞核的物质转运,还包括调节遗传物质的转录和泛素化过程,即泛素对靶蛋白进行特殊性修饰过程等等[23]。我们设想SPAG6定位于微管依赖于这些ARM序列。为研究这些ARM区域对SPAG6在细胞内定位的影响,在携带GFP的表达全长SPAG6真核表达质粒的基础上,我们又构建了依次缺失ARM序列的6个携带绿色荧光蛋白基因的真核表达载体,并在CHO细胞中表达定位,结果发现仅全长SPAG6定位于微管,其他6个缺失ARM的蛋白都不能定位于微管。由于即使缺失最C端ARM的蛋白都不能定位于微管,因此,该ARM序列有可能是介导SPAG6定位于微管的关键区域。缺失C端6个ARM的最短的SPAG6/GFP融合蛋白即实验命名的ARM7,在大部分细胞主要表达在整个细胞中,有些仅在细胞特点部位局部表达,提示最N端的ARM可能有特殊功能。该ARM序列对SPAG6的功能有待于进一步研究。

图5 SPAG6/GFPP 在CCHHOO细胞内定位
本研究初步探讨了SPAG6结构与在细胞内定位的关系,为进一步研究SPAG6的生物学特性及与其他蛋白相互作用奠定了初步基础。本研究今后将更深入的利用分子生物学的相关技术,在分子水平和细胞水平的基础上,尝试从整体水平去分析,利用RNA干扰、基因敲除、酵母双杂交等手段全面而系统研究该蛋白的更多功能。不仅可进一步的探讨SPAG6基因的功能,而且可深入的研究此基因ARM区域的功能,可以为SPAG6基因引起的一类疾病的诊断和治疗提供新的思路和方法。
参 考 文 献
1 董金金, 苗晋华, 王鹏飞. 精子形态学分析在男性不育诊治中的应用. 山西医药志•上半月 2012; 41(6): 575-576
2 徐晓博, 阿周存. 无精症因子与男性不育症的研究概况.中国临床医学 2011; 18(6): 883-884
3 Hann MC, Lau PE, Tempest HG. Meiotic recombination and male infertility: from basic science to Clinical reality? Asian J Androl 2011; 13(2): 212-218
4 傅耀文, 常喜化, 蔡露. 睾丸生精过程中的细胞凋亡及影响因素. 中国男科学杂志 1999; 13(2): 120-123
5 宋博, 蔡志明. 精子形成的调控机制及其进展. 现代泌尿外科杂志 2011; 16(6): 575-577
6 余振东, 桂耀庭, 唐爱发, 等. 精子发生基因表达的调控研究进展. 中国实验诊断学 2006; 10(1): 104-107
7 Tamowski S, Acton KI, Carrell DT. The use of transyenic mouse models in the study of male infertility. Syst Biol Reprod Med 2010; 56(3): 260-273
8 Zhang Z, Jones BH, Tang W, et al. Dissecting the axoneme interactome: the mammalian orthologue of Chlamydomonas PF6 interacts with sperm-associated antigen 6, the mammalian orthologue of Chlamydomonas PF16. Mol Cell Proteomics 2005; 4(7): 914-923
9 Neilson LI, Schneider PA,Van Deerlin PG, et al. cDNA cloning and characterization of a human sperm antigen(SPAG6) with homologyto the product of the chlamydomonas PF16 locus. Genomics 1999; 60(3): 272-280
10 Sapiro R, Kostetskii I, Olds-Clarke P, et a1. Male infertility impaired sperm motility,and hydrocephalusin mice deficient in sperm-associated antigen 6. Mol Cell Biol 2002; 22(17): 6298-6305
11 Zhang Z, Sapiro R, Kapfhamer D, et al. A Sperm-associated WD repeat protein orthologous to Chlamydomonas PF20 associates with Spag6,the mammalian orthologue of Chlamydomonas PF16. Mol Cell Biol 2002; 22(22): 7993-8004
12 Smith EF, Lefebvve PA. The role of central apparatus components in flagellar motility and microtubule assembly. Cell Motil Cytoskeleton 1997; 38(1): 1-8
13 Zhang Z, Tang W, Zhou R, et al. Accelerated mortality from hydrocephalus and pneumonia in mice with a combined defi ciency of SPAG6 and SPAG16L reveals a functional interrelationship between the two central apparatus proteins. Cell Motil Cytoskeleton 2007; 64(5): 360-376
14 Zhang Z, Jones BH, Tang W, et al. Dissecting the axoneme interactome: the mammalian orthologue of Chlamydomonas PF6 interacts with sperm-associated antigen 6, the axoneme interactome:the mammalian orthologue of Chlamydomonas PF16. Mol Cell Proteomics 2005; 4(7): 914-923
15 Silina K, Zayakin P, Kalnina Z, et al. Sperm-associated antigens as targets for cancer immunotherapy: expression pattern and humoral immune response in cancer patients. J Immunother 2011; 34(1): 28-44
16 Sapiro R, Tarantino LM, Velazquez F, et al. Sperm antigen 6 is the murine homologue of the Chlamydomonas reinhardtii central apparatus protein encoded by the PF16 locus. Biol Reprod 2000; 62(3): 511-518
17 Akhmanova A, Steinmetz MO. Microtubule +TIPs at a glance. J Cell Scl 2010; 123(Pt 20): 3415-3419
18 Jolly AL, Kim H , Srinivasan D, et al. Kinesin-1 heavy chain mediates microtubule sliding to drive changes in cell shape. Proc Natl Acad Sci USA 2010; 107(27): 12151-12156
19 Loiseau P, Davies T ,Williams LS, et al. Drosophila PAT1 is required for Kinesin-1 to transport cargo and to maximize its motility. Development 2010; 137(16): 2763-2772
20 Tanenbaum ME, Medma RH. Mechanisms of centre-some separation and bipolar spindle assembly. Dev Cell 2010; 19(6): 797-806
21 Miller AL, Bement WM. Cell division: the need for speed. Curr Biol 2009; 19(23): R1071-R1073
22 Adachi A, Kano F, Tsuboi T, et al. Golgi-associated GSK3-beta regulates the sorting process of post-Golgi membrane trafficking. J Cell Sci 2010; 123(Pt 19): 3215-3218
23 Samuel M A, Salt J N, Shiu S H, et a1. Multifunctional arm repeat domains in plants. Int Rev Cyto1 2006; 253: 1-26
(2014-03-28收稿)
**通讯作者, Email: zhangzhibing@hotmail.com
doi:10.3969/j.issn.1008-0848.2015.01.001
中图分类号R 698.2; R 394.33
*基金项目资助: 国家自然科学基金(81172462)、(81300536)、(81302401)及湖北省自然科学基金(2012FFB04904)、(2013CFB331)资助
Effect of ARM domain deletion on SPAG6 subcellular localization in mammalian cells*
Wang Zhiqiong1, 2, Zhang Ling1, Shi Yuqin1, Li Hongfei1, Zhang Zhibing1, 3**
1. School of Public Health,Wuhan University of Science and Technology, Wuhan 430065, Hubei, China; 2. Department of Obstetrics and Gynecolgy and Biochemistry; 3. Virginia Commonwealth University, Richmond Corresponding author: Zhang Zhibing, Email: zhangzhibing@hotmail.com
AbstractObjective To examine the effect of armadillo (ARM)domains on sperm- associated antigen 6 (SPGA6) localization in CHO cells. Metthhooddss The sequences coding different ARM domains were amplifi ed by polymerase chain reaction (PCR), and were subcloned into the pEGFP-N2 vector. These plasmids were transfected into CHO cells. The expression of the GFP-tagged SPAG6 proteins was proved by Western blot analysis, and localization of GFP-tagged full length and truncated SPAG6 proteins was observed by using laser scanning confocal microscopy. Ressuullttss Truncated SPAG6 proteins with series deletion of ARM domains were successfully constructed into the mammalian expressing vector carrying the green fl uorescent protein gene, pEGFP-N2. The expression of the fusion protein was confi rmed by Western blot analysis. The full length GFP-tagged SPAG6 protein was 1ocalized on the microtubule in the transfected CHO cells, but none of the truncated proteins was localized on the microtybule. Concluussiioonn Microtubule localization of mouse SPAG6 protein is dependent on full length sequence. Construction of the plasmids expressing full length and truncated SPAG6 proteins provides an important tool for further study SPAG6 function.
