华南地区推广应用大豆品种对疫霉根腐病的抗性评价
2015-07-12程艳波马启彬牟英辉谭志远
程艳波,马启彬,牟英辉,谭志远,吴 鸿,年 海,
(1 国家大豆改良中心 广东分中心/华南农业大学 农学院,广东 广州510642;2 亚热带农业生物资源保护与利用国家重点实验室,华南农业大学,广东 广州510642)
在大豆Glycine max(L.)Merrill 生产中存在很多种病害,如大豆花叶病毒病、锈病、白粉病、大豆疫霉根腐病等病害,其中大豆疫霉根腐病是由大豆疫霉菌Phytophthora sojae 引起的严重危害大豆生产的一种世界性病害.虽然它的起源相对较晚,但它却是大田作物唯一的疫霉病害[1].大豆疫霉根腐病常年可造成大豆10%~30%的减产,严重甚至可达60%~90%以上.给全球大豆生产每年造成的损失达10亿~20 亿美元[2].使用抗病品种是防治大豆疫霉根腐病最经济有效的方法,筛选和评价华南地区大豆品种对大豆疫霉根腐病的抗性以及热带亚热带地区大豆生产极其重要.我国大豆主产区在东北和黄淮海地区,由于大豆多年连作种植,大豆疫霉根腐病发生较重,对大豆疫霉根腐病原菌的分离、鉴定和抗性资源筛选前人[3-10]已做了大量的工作,并筛选了一批抗大豆疫霉根腐病的品种或资源.我国东北和黄淮海大豆品种或资源中具有较丰富的抗大豆疫霉根腐病的双抗或多抗品种或资源,存在多抗的大豆品种和资源,但抗病品种数量和抗性水平存在地区间差异[11-19].来源于东北和黄淮海大豆种植区的抗性品种不能直接在热带亚热带地区利用.本研究系统地评价华南地区主要推广应用的大豆品种及其骨干亲本对大豆疫霉根腐病的抗性,筛选出抗病品种,以期为华南地区抗病育种和热带亚热带地区大豆生产提供重要抗性材料.
1 材料与方法
1.1 材料
67 个大豆品种(表1)分别来自广东、广西、福建、江西、四川等地.
PGD1 由华南农业大学国家大豆改良中心广东分中心在广东大豆疫霉根腐病病株中发现并分离得到,Pm14、Pm28、PNJ1、PNJ3、PNJ4、P6497 由南京农业大学国家大豆改良中心邢邯教授惠赠.培养基是V8 蔬菜汁-碳酸钙培养基[10].
1.2 方法
将健康的大豆种子播于以蛭石为基质的直径为9 cm 的花盆中,每品种种植2 盆,每盆种12 粒,留10株苗,于播种当天、第3 和第7 天各浇水1 次.大豆培养在光照培养室中,温度控制在20~30 ℃,光照14 h.播种2 d 后取保存在10 ℃冰箱中的大豆疫霉菌,用接种环挑取0.5 cm2左右的边缘菌落,转接于具有培养基的培养皿中央,25 ℃温箱中倒置培养5~6 d.
取美国进口的V8 蔬菜汁120 mL,加入碳酸钙1.2 g,在4 000 r·min–1、25 ℃条件下离心8 min,取上清液100 mL,加超纯水定容到1 L,再加入15 g 琼脂;将培养基和培养皿在121 ℃灭菌锅灭菌20 min后,将培养基和培养皿取出,放置于超净工作台冷却,倒入直径为9 cm 的培养皿备用,培养基的厚度以0.3 cm 为宜.播种7 d 后第1 对真叶平行时接种.采用下胚轴伤口接种法[6,19-20],略有修改,用消毒后的刀片在大豆子叶节下约1 cm 处划一伤口,伤口以能看到组织液为宜,一般深度不超过大豆茎粗的1/3,然后取25 ℃恒温箱中培养5 d 的大豆疫霉菌,在菌株菌落外缘割取带有菌丝体的培养基(边长约3 mm 的方块)嵌入伤口中,菌丝面接触伤口,接种后把大豆放在塑料薄膜架子内,向膜内喷水,保湿膜内相对湿度在90%以上,保湿膜内不开灯,保湿12 h,打开保湿膜,同时打开日光灯.
接种5 d 后进行病情调查.一般接种后第3 天感病植株在接种部位折倒,第5 天死亡.抗病植株接种后无变化或接种部位局部褐变,整株继续正常生长.抗感标准参照文献[10,15,18]等方法:70%或以上的植株死亡为感病(Susceptible,S);30%或以下植株死亡为抗病(Resistant,R);死亡植株率在31%~69%的品种为中间型(Intermediate,I).试验以Williams(rps)、华春2 号和华夏3 号为感病对照,粤夏2011-4 和桂夏1 号为抗病对照.表现中间型和抗病材料重复鉴定2 次.
死亡率=死亡株数/接种总株数×100%,
毒力频率=接种后感大豆疫霉菌的个数/接种疫霉菌的个数×100%,
侵染率=接种某一疫霉菌感病品种个数/接种品种个数×100%.
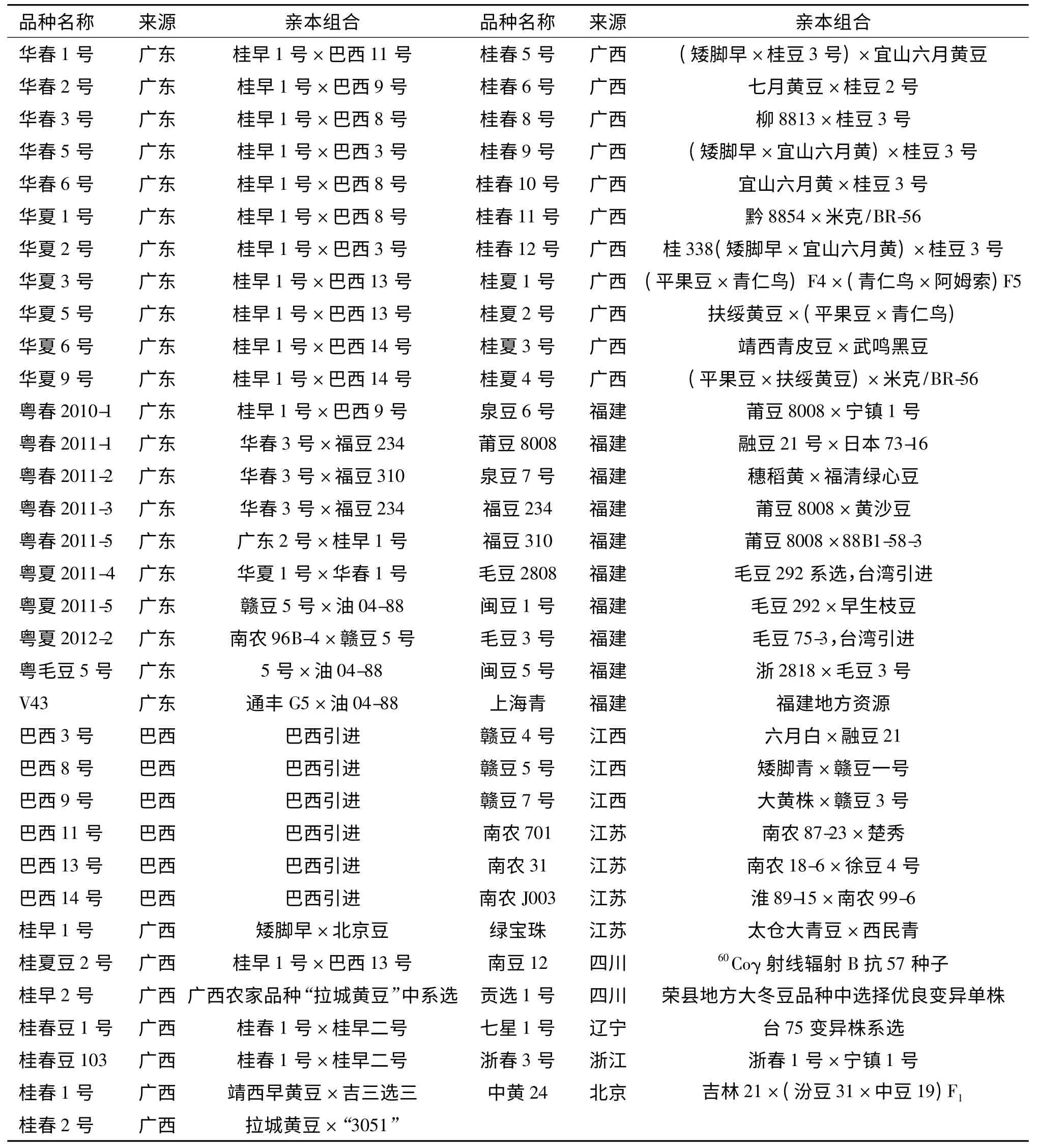
表1 大豆品种(系)名称及来源Tab.1 Names and origins of soybean cultivars or lines used in this study
2 结果与分析
2.1 大豆抗病基因的鉴定
所选用的7 个大豆疫霉菌株具有不同的毒性基因或毒性基因组合(表2),其中大豆疫霉菌Pm14 的毒性最强,其次是Pm28.PGD1 是在广东发现并分离出的大豆疫霉菌,其毒力公式为:1a,1b,1k,2,3a,3c,4,5,6,7.
2.2 华南地区大豆品种及骨干亲本对大豆疫霉菌的抗性评价
用7 个不同毒力的大豆疫霉菌株接种鉴定67个华南地区推广应用的大豆新品种(系)及其亲本,抗性鉴定结果如下:
67 个大豆品种对7 个不同毒力菌株PGD1、Pm14、Pm28、PNJ1、PNJ3、PNJ4、P6497 的侵染率不同,侵染率分别为86.6%、80.6%、85.1%、61.2%、77.6%、74.6% 和80.6%,其中接种大豆疫霉菌PGD1 的侵染率最高,其次是接种Pm28 和Pm14,接种PNJ1 和PNJ4 的侵染率较低(表3).所以华南地区推广应用的大豆品种(系)及骨干亲本对7 个大豆疫霉菌的抗性品种较少.
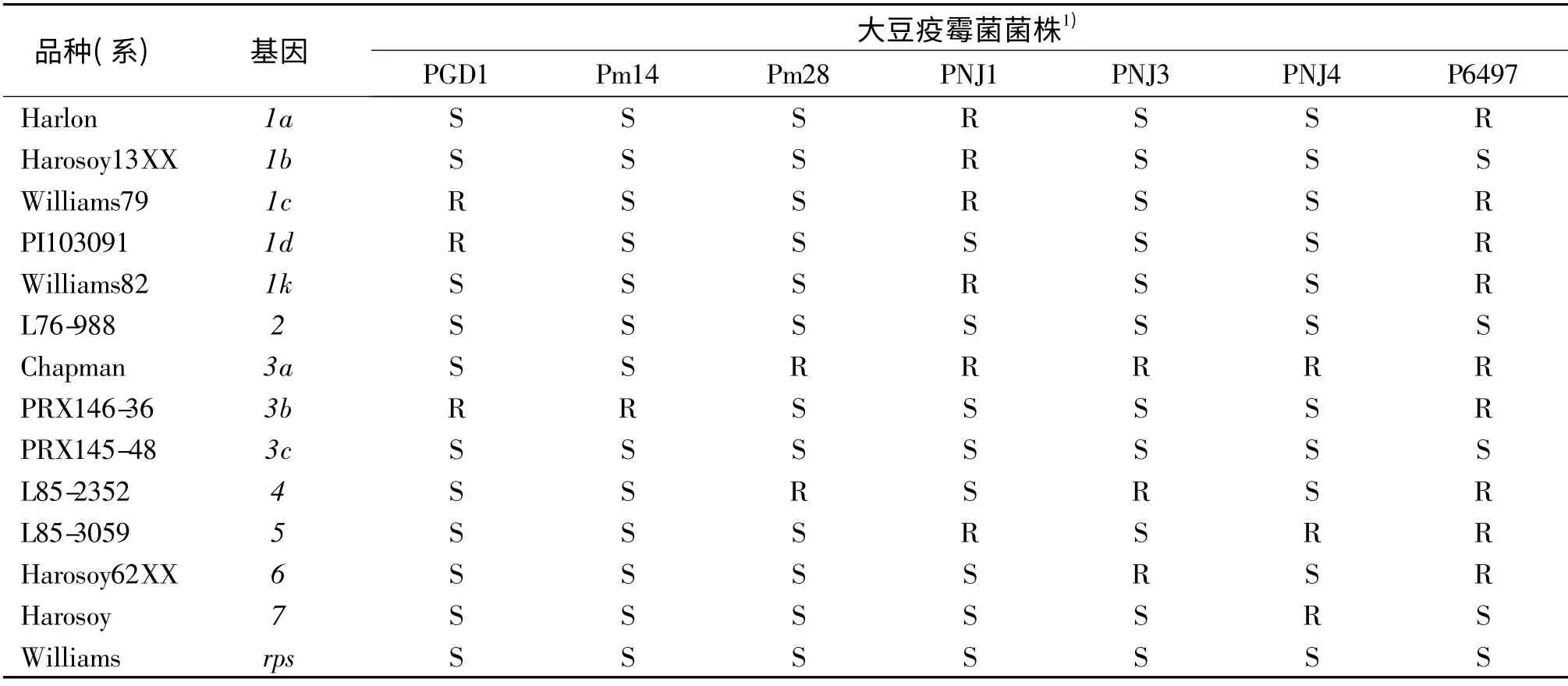
表2 含已知抗病基因大豆品种或品系对7 个大豆疫霉根腐菌鉴别菌株的反应Tab.2 Differential reactions of soybean cultivars or lines possessing Phytophthora resistance genes after being inoculated with 7 strains of Phytophthora sojae
对于同一大豆品种,不同菌株对其毒力频率也存在差异,毒力频率最高为100%,即感7 个大豆疫霉菌株的品种有29 个,占鉴定品种的43.3%;毒力频率最低的为0,仅有桂夏1 号,该品种接种大豆疫霉菌PGD1、Pm14、PNJ1、PNJ4 和P6497 表现抗病,接种大豆疫霉菌Pm28 和PNJ3 表现为中间型(表3),桂夏1 号可作为抗病育种的重要抗源.
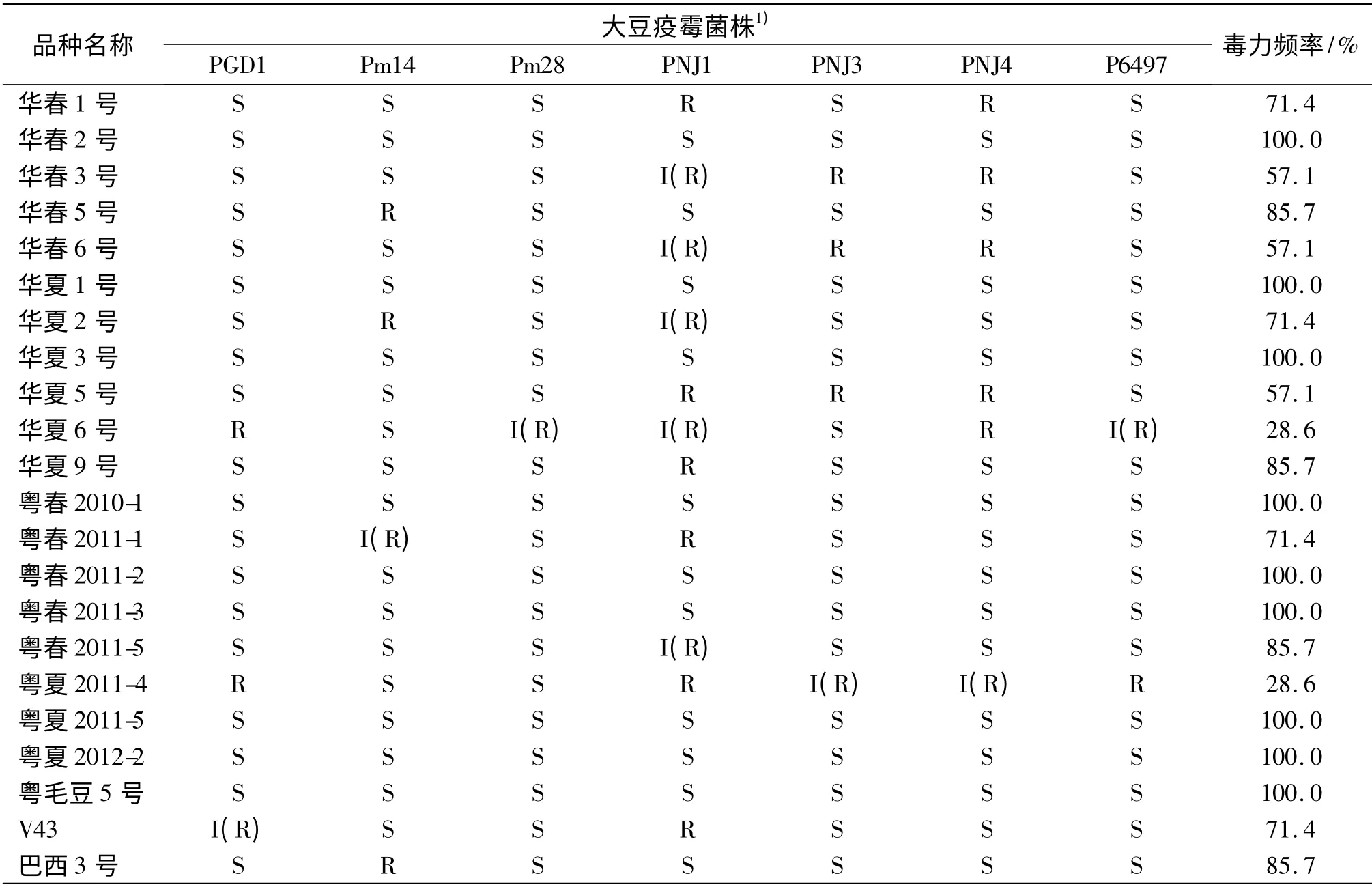
表3 华南地区推广应用大豆品种(系)及骨干亲本对7 个大豆疫霉菌株的反应Tab.3 Responses of different soybeans and partial parents inoculated with 7 strains of Phytophthora sojae in South China
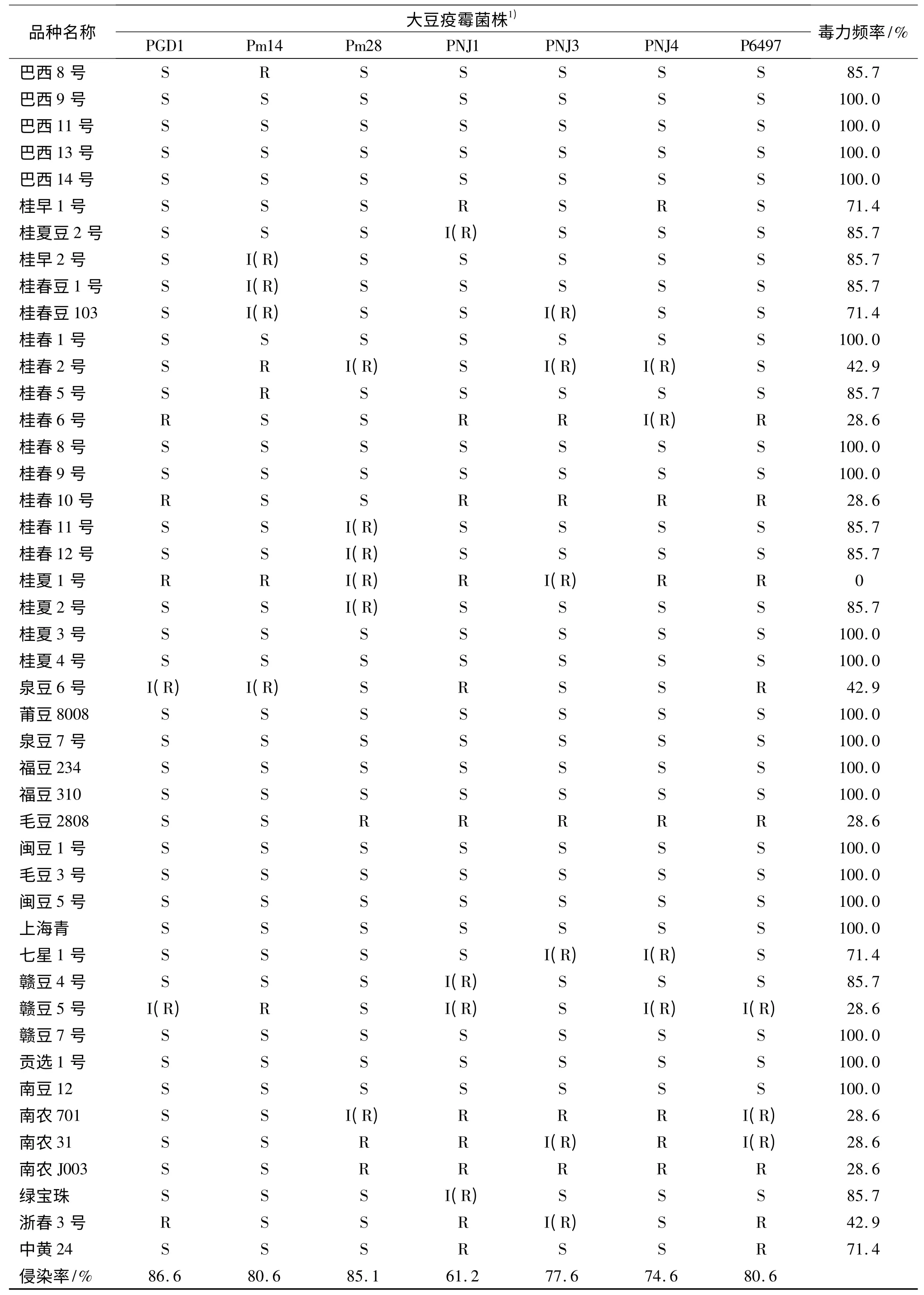
续表3 Tab.3 continued
由表3 还可知,广东选育的大豆品种或品系多抗疫霉菌的品种较少,这些品种具有共同的母本桂早1 号,父本多为从巴西引进的高产耐酸铝品种,由于选用的亲本对7 个大豆疫霉菌的毒力频率较低,这可能是广东所选育的品种多抗性品种较少的主要原因.
3 讨论
本研究利用7 个不同毒力大豆疫霉菌对华南地区67 个推广应用的大豆品种和骨干亲本进行抗疫霉根腐病鉴定,其中大豆疫霉菌PGD1 的侵染率最高.大豆疫霉菌PGD1 是华南农业大学在广东分离的新的大豆疫霉菌株,该小种可能是华南地区大豆疫霉菌优势小种.对7 个大豆疫霉菌都表现为感病的品种有29 个,占鉴定品种的43.3%;毒力频率为0的品种仅有桂夏1 号.所以华南地区生产上应用的大豆品种抗大豆疫霉根腐病的品种较少,其中抗大豆疫霉菌Pm14 研究结果与任海龙等[10]利用大豆疫霉菌Pm14 对华南地区大豆品种和育种亲本进行抗性鉴定结果较一致.
华南地区目前生产上主要应用的品种遗传背景相似,造成这些品种抗大豆疫霉根腐病较少的原因可能与选用的育种亲本有很大关系.巴西3 号、巴西8 号、巴西13 号等品种在巴西大面积推广应用,是巴西主要推广品种,具有高产耐酸铝特性.利用本地高蛋白品种桂早1 号与巴西高产品种杂交,选育的适合华南地区种植的大豆品种产量显著提高,但由于引进的巴西品种几乎全感7 个大豆疫霉根腐菌.所以华南农业大学选育的大豆品种多抗大豆疫霉菌品种较少.
华春5 号和华夏2 号抗大豆疫霉菌Pm14,这2个品种都是利用“桂早1 号×巴西3 号”杂交选育而成,桂早1 号感大豆疫霉菌Pm14,巴西3 号抗大豆疫霉菌Pm14,说明选育的抗病品种的抗病基因可能是由巴西3 号遗传而来;华春1 号(桂早1 号×巴西11号)、华夏5 号(桂早1 号×巴西13 号)、华夏9 号(桂早1 号×巴西14 号)、粤春2011-1(华春3 号×福豆234)、粤夏2011-4(华夏1 号×华春1 号)这些品种接种大豆疫霉菌PNJ1 表现抗病,巴西8 号、巴西11 号、巴西13 号、巴西14 号和福豆234 接种大豆疫霉菌PNJ1 表现感病,华春3 号为中间型.抗病品种与桂早1 号具有亲缘关系,同时桂早1 号抗大豆疫霉菌PNJ1,所以这些品种的抗病基因可能由桂早1 号遗传而来.华春1 号、华春3 号、华春6 号、华夏5号、华夏6 号都是桂早1 号衍生品种,桂早1 号抗大豆疫霉菌PNJ4,这些品种抗大豆疫霉菌PNJ4 可能也是由桂早1 号遗传而来.齐黄1 号、鲁豆4 号、早熟18、铁荚四粒黄和吉林20 号是我国黄淮海大豆育成品种的主要亲本[19-20],齐黄1 号于1962年由山东省农业科学院系统选育而成,已衍生出92 个大豆品种[20],陈晓玲等[15]研究发现,豫豆系列、周豆系列、科新系列大豆品种在黄淮海地区生产上具有广谱的抗性,可能是含有这些抗病品种的抗病基因.豫豆25号抗多个不同毒力的大豆疫霉菌[21],该品种的原始亲本中含有齐黄13 的血缘,齐黄13 也是齐黄1 号的衍生品种[20],因此豫豆25 号具有广谱抗病基因可能来源于齐黄1 号.所以,筛选抗不同毒力的大豆疫霉菌株的种质资源对抗病育种非常重要.
很多研究表明,大豆疫霉根腐病基因由1 对单显性基因控制[22-23],然而,在筛选评价品种的抗性过程中发现,一些品种的亲本对某疫霉菌株接种鉴定表现感病,而在杂交后代选育的品种中表现抗病.如华夏6 号和粤夏2011-4 接种鉴定抗大豆疫霉菌PGD1,华夏6 号和粤夏2011-4 的亲本及亲本的亲本都感大豆疫霉菌PGD1;华春3 号(桂早1 号×巴西8号)、华春6 号(桂早1 号×巴西8 号)和华夏5 号(桂早1 号×巴西13 号)接种大豆疫霉菌PNJ3 表现抗病,而亲本都感大豆疫霉菌PNJ1;粤夏2011-4(华夏1 号×华春1 号)抗大豆疫霉菌P6497,而亲本及亲本的亲本都感大豆疫霉菌P6497;出现感病品种相互杂交选育出抗病品种的现象,不符合单基因遗传规律,所以这些新品种的抗病性可能是由于基因重组所致.很多研究表明大豆品种对大豆花叶病毒病多数株系的抗性由1 对显性基因控制[24-26],华春5号高抗SC3、SC8、SC15 和SC18 生理小种,发病率几乎为0,而亲本都为高感.这些品种的选育只从产量方面决选,而对抗病性未做接种鉴定,按照现有遗传理论无法解释,而且出现抗病的概率较高,所以在常规育种中,应以产量提高为选种标准,兼顾品种的抗病性鉴定.
4 结论
大豆疫霉菌PGD1 是在广东发现并分离出的新的大豆疫霉菌株,是华南地区大豆疫霉菌优势小种,抗大豆疫霉菌PGD1 的品种有华夏6 号、粤夏2011-4、桂春6 号、桂春10 号、桂夏1 号、浙春3 号,占9.0%,这些品种可作为华南地区抗大豆疫霉菌PGD1 病育种的重要亲本.29 个品种同时感7 个大豆疫霉菌,占43.3%,所以华南地区生产上应用的抗大豆疫霉根腐病品种较少,需要加大抗疫霉根腐病种质资源筛选和抗病品种选育力度.
致谢:南京农业大学邢邯教授惠赠鉴别寄主和菌株.广西农业科学院陈渊研究员、陈怀珠研究员、梁江研究员,福建省农业科学院林国强研究员,江西省农业科学院王瑞珍研究员,四川省自贡市农业科学研究所杨华伟高级农艺师等为本研究提供大豆品种(系).华南农业大学宋恩亮同学、任海龙同学和实验技术员张文珊提供技术支持和帮助,在此表示衷心感谢!
[1]SCHMITTHENNER A F.Problems and progress in control phytophthora root rot of soybean[J].Plant Dis,1985,69(4):362-368.
[2]TYLER B M.Phytophthora sojae:Root rot pathogen of soybean and model oomycete[J].Mol Plant Pathol,2007,8(1):1-8.
[3]朱振东,王晓鸣,常汝镇,等.黑龙江省大豆疫霉菌生理小种鉴定及大豆种质的抗性评价[J].中国农业科学,2000,33(1):62-67.
[4]李长松,路兴波,刘同金,等.大豆疫霉根腐病菌生理小种的鉴定及品种抗病性筛选[J].中国油料作物学报,2001,23(2):60-62.
[5]许修宏,吕慧颖,曲娟娟,等.大豆疫霉根腐病菌分离方法及发病地区初步调查[J].大豆科学,2002,21(2):147-150.
[6]徐鹏飞,吴俊江,范素杰,等.大豆疫霉根腐病菌的分离鉴定及种质资源对3 号生理小种的抗性评价[J].大豆科学,2010,29(2):272-275.
[7]王晓鸣,朱振东,王化波,等.中国大豆疫霉根腐病和大豆种质抗病性研究[J].植物病理学报,2001,31(4):324-329.
[8]吕慧颖,孔凡江,许修宏,等.东北三省大豆种质资源对大豆疫霉根腐病的抗性表现[J].中国油料作物学报,2001,23(4):16-18.
[9]任龙翚,张宝强,武晓玲,等.大豆种质对大豆疫霉菌株Pm8 的抗性分析[J].大豆科学,2010,29(1):77-79.
[10]任海龙,马启彬,杨存义,等.华南地区大豆育种材料抗疫霉根腐病鉴定[J].大豆科学,2012,31(3):453-456.
[11]LOHNES D G,NICHELL C D,SCHMITTHENNER A F.Origin of soybean alleles for Phytophthora resistance in China[J].Crop Sci,1996,36(6):1689-1692.
[12]KYLE D E,NICHELL C D,NELSON R L,et al.Response of soybean accessions from provinces in Southern China to Phytophthora sojae[J].Plant Dis,1998,82:555-559.
[13]DORRANCE A E,SCHMITTENNER A F.New sources of resistance to Phytophthora sojae in the soybean plant introductions[J].Plant Dis,2000,84:1303-1308.
[14]朱振东,王化波,王晓鸣,等.黑龙江省主要栽培大豆品种(系)对大豆疫霉根腐病的多抗性评价[J].植物遗传资源学报,2004,5(1):22-25.
[15]陈晓玲,朱振东,王晓鸣,等.大豆品种(系)抗疫霉根腐病基因推导[J].中国农业科学,2008,41(4):1227-1234.
[16]孙石,赵晋铭,武晓玲,等.黄淮地区大豆种质对疫霉根腐病的抗性分析[J].大豆科学,2008,27(3):465-470.
[17]隋喆,黄静,马振川,等.吉林、辽宁大豆品种(系)对大豆疫病的抗病基因鉴定[J].中国油料作物学报,2010,32(1):94-98.
[18]唐庆华,崔林开,李德龙,等.黄淮地区大豆种质资源对疫霉根腐病的抗病性评价[J].中国农业科学,2010,43(11):2246-2252.
[19]夏长剑,张吉清,王晓鸣,等.大豆资源抗疫霉根腐病基因分析[J].中国油料作物学报,2011,33(4):396-401.
[20]徐冉,时传娥,张礼凤,等.黄淮海大豆优异种质齐黄1号的育种应用[J].植物遗传资源学报,2004,5(2):170-175.
[21]SUN S,WU X L,ZHAO J M,et al.Characterization and mapping of RpsYu25,a novel resistance gene to Phytophthora sojae[J].Plant Breeding,2011,130:139-143.
[22]武晓玲,周斌,孙石,等.大豆对大豆疫霉菌株Pm14抗性的遗传及基因定位[J].中国农业科学,2011,2011,44(3):456-460.
[23]BURHAM K D,DORRANCE A E,FRANCIS D M,et al.Rps8,a new locus in soybean for resistance to Phytophthora sojae[J].Crop Sci,2003,43(1):101-105.
[24]LIM S M.Resistance to soybean mosaic virus in soybeans[J].Phytopathology,1985,75(2):199-201.
[25]李海朝,智海剑,白丽,等.大豆对SMV 株系SC-11 的抗性遗传及抗病基因的等位性研究[J].大豆科学,2006,25(4):365-368.
[26]王大刚,马莹,刘宁,等.大豆花叶病毒(SMV)株系SC4和SC8 的抗性遗传分析[J].作物学报,2012,38(2):202-209.
