Cytological Observations on the Fertilization of the Fern Pteridium aquilinum var.latiusculum
2015-07-04CAOJianguoDAIXilingWANGYaoWANGQuanxi
CAO Jianguo,DAI Xiling,WANG Yao,WANG Quanxi
(College of Life and Environmental Sciences,Shanghai Normal University,Shanghai 200234,China)
The fertilization processes in embryophytes,compared to invertebrate animals and algae,is still poorly understood because the fertilization events are internal and inaccessible[1-3].For the ferns,the knowledge about fertilization is limited to a few species,includingPteridium[4-6],Marsilea[7],Athyrium[8],Ceratopteris[2,9,10].InCeratopteris,a sac containing numerous small opaque vesicles is thought to be a barrier to prevent polyspermy[9-10].InPteridiumandAthyrium,spermatozoids are considered to be engulfed firstly into a“fertilization cavity”,from which a single sperm fuse with the egg cytoplasm[4,8].Thereafter,Bell and Duckett indicated that whether the fertilization cavity is an authentic feature of the viable eggs was still unclear[5].And the mechanism of preventing polyspermy was still unknown[5].In present inves-tigation,the cytological processes of fertilization,including the egg penetration,nuclear fusion and polarity establishment of zygote,are described in detail.The mechanism of preventing polyspermy is discussed based on observations.
1 Materials and methods
Spores ofPteridiumaquilinumvar.latiusculumwere collected from plants in Hezhang County,Guizhou Province,China.The culture of the gametophytes has been described in our previous paper[11].For observing the processes of the fertilization and zygote formation,the gametophytes containing near matured archegonium were plunged into the suspension of spermatozoids.The viable archegonia need at least 20-30 min flooding.When the spermatozoids were seen crowding into the archegonium through the open neck(under stereomicroscope),the time was recorded.The specimens were moved out and fixed at the time of within 3min,10 min,30 min,3h,12h,24hafter fertilization respectively.About five to ten fertilized archegonia were examined for each time.The specimens were picked up and fixed with 3%glutaraldehyde in 0.1mol/L phosphate buffer at room temperature for 6-8h.These specimens were subsequently washed with the same buffer for three times and post-fixed in osmic acid(1%solution in phosphate buffer)for 2hand then washed with the same buffer three times.After dehydration in a graded acetone series,the materials were infiltrated with a mixture of acetone and spurr’s resin and then embedded in pure spurr’s resin.Specimens were thick sectioned first for the presence of the fertilized egg and thin-sectioned with a diamond knife on a Leica UC7ultramicrotome(Germany).The thin sections were stained with uranyl acetate and lead citrate and observed with an H-600electron microscope(Hitachi,Japan).
2 Results
2.1 Egg penetration
In present investigation,eggs before and after fertilization are examined.It is discovered that a typical feature of the viable mature egg is numerous vesicles occurring in the upper part of the egg cytoplasm just beneath the fertilization pore(PlateⅠ,1).The nuclear evaginations,containing osmiophilic bodies in the basal region of the evagination,can be identified near the nucleus surface(PlateⅠ,1).
In order to obtain the moment of fertilization,more than 10 mature archegonia,which are fertilized within 3 min,were examined.A spermatozoid which is penetrating the egg is captured(PlateⅠ,2,3).The spermatozoid enters the egg through the fertilization pore(PlateⅠ,2,3).A fertilization cavity is formed most likely by the spiral motion of the spermatozoid(Plate Ⅰ,2,3).The diameter of the fertilization cavity is about 6.6μm,which is just equal to the transverse diameter of the spermatozoid(PlateⅠ,2,3).Serial sections showed that the fertilization cavity contains only one spermatozoid and no other sperms enter the cytoplasm at the moment.The organelles in the egg cell,including the mitochondria,endoplasmic reticula,are still recognizable(Plate Ⅰ,4).The egg nucleus is highly irregular with nuclear evaginations(PlateⅠ,4).
Regrettably,we can not capture the moment that the spermatozoid fused with the egg cytoplasm.However,as soon as the spermatozoid enters the egg cytoplasm,the fertilized egg shrinks markedly(PlateⅠ,5).The volume of the fertilized egg decreases to almost one-half to that of the unfertilized egg.The cytoplasm of the fertilized egg becomes dense and opaque(PlateⅠ,5).The membranous organelles of the egg,including the mitochondria and endoplasmic reticula of the egg are barely identifiable.Only plastids which contain the starch grains remain recognizable in the egg cytoplasm(PlateⅠ,6).
The fertilization cavity remains to lie beneath the fertilization pore consistently.Repeat experiments showed that such a fertilization cavity can not prevent the other spermatozoids from entering into the fertilization cavity.PlateⅠ,5showed that another spermatozoid has entered the fertilization cavity.Our observations showed that at least 2-6 spermatozoids can enter the cavity after the first spermatozoid fused with the egg cytoplasm.And the fertilization cavity increases its transverse diameter to 12μm(PlateⅡ,1).But all the observations showed that the spermatozoids in the fertilization cavity can not enter the egg cytoplasm.Penetration by the supernumerary spermatozoid into the egg cytoplasm was not encountered in all sections examined.After the spermatozoid entered the egg cytoplasm,the male organelles and the male nucleus can be identified obviously.The male organelles generally lie near the upper surface of the egg nucleus(PlateⅠ,6).
2.2 Nuclear fusion
At approximately 30min after fertilization,the fertilized egg was still shrunken and the cytoplasm kept dense and opaque.The fertilization cavity is obvious and contains at least 3 supernumerary spermatozoids.At this time,the nucleus of the spermatozoid begins to fuse with the egg nucleus(Plate Ⅱ,1).The multilayered structure of the male gamete anchored to the egg nuclear surface.The male nucleus,once a solid rod of chromatin,becomes decondensed.Most of the male nucleus has entered the egg nucleus.Sometimes,vesicle can be seen between the chromatin and the microtubular ribbon(Plate Ⅱ,2,3).The male nuclear envelope reappears around the male chromatin and connects with the egg nuclear envelope(PlateⅡ,3,arrows).In contrast to the egg chromatin,the male chromatin still shows electronic opaque and easily recognizable in the egg chromatin(PlateⅡ,2).
2.3 Egg expanding
At approximately 3 h after fertilization,the fertilized egg expanded fully to fill the venter cavity of the archegonium.The fertilization cavity now has been squashed by the expended zygote and the unfertilized sperms remained near the fertilization pore(Plate Ⅱ,4).The zygote now had a clear structure resembling that of the mature egg.The organelles in the zygote cytoplasm become abundant and easily recognizable.The ERs are abundant in the periphery cytoplasm.The mitochondria,showing spherical,oval and dumb-shaped,distribute evenly around the zygote nucleus(PlateⅡ,5).It is worthy to indicate that numerous vesicles(cytoplasm vacuolization)occur in a side of the egg cytoplasm.The original irregular zygotic nucleus now shows a relatively regular shape in outline.A side of the zygotic nucleus is expanded most likely by fusion of the male chromatin(Plate Ⅱ,4).The nuclear evaginations are still connect with the nucleus(Plate Ⅱ,5).And the electronic opaque bodies are seen at the base of the nuclear evaginations(Plate Ⅱ,5,arrows).The MTR and the flagella are present in the cytoplasm above the nucleus(Plate Ⅱ,5).However,the flagella and the MTR lose the membrane covering the microtubules(PlateⅡ,5,insert).
2.4 The zygote polarity establishment(12-24h after fertilization)
At approximately 12 h after fertilization,a conspicuous feature of the zygote is that the original cup-shaped nucleus now contracts to a relatively round shape.The upper surface of the nucleus still shows irregular outline(Plate Ⅲ,1).At this stage,the nuclear evaginations have detached from the nucleus bodies(Plate Ⅲ,2,3).No male organelles can be identified in the zygote cytoplasm.Some clusters of lipid droplets occur in the cytoplasm of zygote(Plate Ⅲ,1,2).The mitochondria become well developed in the zygote cytoplasm(Plate Ⅲ,3).Cell membrane reoccurs outside the cytoplasm(Plate Ⅲ,3,arrow).Two or more large vacuoles occur at a side of the zygote.Thus,the zygote establishes a horizontal polarity(PlateⅢ,1).
Approximately 24hafter fertilization,a large vacuole,which is most likely formed by coalescence of the small vesicles,always lies in the posterior part of the zygote.The nucleus,with 2-3conspicuous nucleoli,becomes spherical(Plate Ⅲ,4).The mitochondria,some of them are dumb-shaped,are abundant in the zygote cytoplasm(Plate Ⅲ,5).Plastids are still lack internal lammela.Sometimes,residues of metabolites or degenerated organelles are seen entering into a vacuole(Plate Ⅲ,6).The unfertilizing spermatozoids outside the zygote can still be seen to block the fertilization pore(PlateⅢ,1,4).But they were compressed into a small a-mount above the zygote(PlateⅢ,4).
3 Discussion
Although some aspects of the fertilization in the fernPteridiumaquilinumhave been reported[5-6],many details of the fertilization,including the syngamy fashion,prevention of polyspermy,digestion of the male organelles in the egg cytoplasm and fate of the nuclear evaginations,are still unclear.The present investigation gives some new observations about the fertilization processes.
3.1 Syngamy and fertilization cavity formation
In previous studies onP.aquilinum,it was believed that the spermatozoid breaks the egg membrane(egg envelope)using the flagella or screwlike motion and enters the egg cell[5-6].Fasciatiet al.also considered that the gyrations of a spermatozoid at the receptive spot,which last up to 15 min,may serve to open a pore in the venter coat through which the male gametes can penetrate into the egg cell in the fernAthyriumfilix-femina[8].In our previous study,it is found that the mature eggs ofPteridiumhave a fertilization pore[11].The present investigation showed that the spermatozoid penetrate the egg cell through the fertilization pore,which is similar to the fernCeratopteristhalictroides[2-3,10].However,the spermatozoid ofP.aquilinumalways prepares a fertilization cavity before the sperm fuses with the egg cytoplasm.
Duckett and Bell firstly considered that the fertilization cavity was a normal feature of the mature eggs ofPteridium[4].But in a subsequent report,they indicated that whether it was an authentic feature of viable egg was still not clear[5].The present investigation revealed that all the fertilized eggs possess the fertilization cavity and all the fertilized eggs can develop into an embryo and seedling.So the fertilization cavity is undoubtedly the normal structure formed by penetration of the spermatozoid.Similar structure was observed inLygodiumunder observations of light microscope and was described as the hyaline area[12].InAthyriumit was called fertilization vesicle[8].So far,how the fertilization cavity is formed is still un-clear.The present observations show that the formation of the fertilization cavity is accompanied by disappearance of the vesicles in the mature egg cytoplasm.So it is reasonable to infer that the fertilization cavity is formed by the coalescence of numerous vesicles of the mature egg.Certainly,stirring of the spermatozoid with spiral motion undoubtedly promote the coalescence of the vesicles and formation of the fertilization cavity.Moreover,collision of the spermatozoids on the protoplast of the egg may help the formation and expanding of the fertilization cavity.
3.2 Prevention of polyspermy
Organisms with sexual reproduction have developed the cytological mechanisms for preventing polyspermy[13-15].Although some aspects of fertilization in the fernP.aquilinumhave been observed,they can not explain the mechanism for preventing polyspermy[4-6].In present investigation,the possible mechanism for preventing polyspermy ofPteridiummay attribute to three respects.For the first aspect,the prolonged spiral motion of the first spermatozoid penetrating the egg blocks supernumerary spermatozoids entering the egg.On the other hand,the pyknosis of the egg protoplasm at the moment of the fertilization is undoubtedly unfavorable for the entrance of supernumerary spermatozoids.And the pyknosis of the egg protoplasm is most likely the key reason for preventing polyspermy.Moreover,the membranous structures around the fertilization cavity may form a barrier,which contributs to prevent polyspermy.These membranous structures may be derived from the membrane of the vesicles,which are commonly seen in the upper part of the mature egg.After the first spermatozoid enters the egg cytoplasm,the fertilization cavity still remains beneath the fertilization pore.Although the supernumerary spermatozoids can enter the fertilization cavity,they have no chance to fuse with the egg cytoplasm.
The supposed mechanism of preventing polyspermy in the fernP.aquilinumseems differs from the two species,MarsileavestitaandCeratopteristhalictroides.In the heterosporous fernM.vestita,apolysac-charide wall and a newly formed extracellular layer were believed to be used as barriers to polyspermy[7].In the fernC.thalictroides,a sac containing numerous opaque vesicles was discovered to block the fertilization pore persistently,which is considered to be cytological mechanism for preventing polyspermy inC.thalictroides[3,10].
3.3 Nuclear fusion
The male nuclear behavior in the egg cytoplasm and karyogamy are similar to previous observations inPteridium[5]andAthyrium[8].The male chromatin is dissociated from the MTr,and a layer of envelope reappears around the male chromatin.It is considered that the reappearance of the nuclear envelope is ready for the nuclear fusion[3].In present investigation,it is observed that the envelopes of male and female nucleus are connected with each other.And most male nucleus has entered the egg nucleus.The cytoskeletal apparatuses of the spermatozoid,including multilayer structure and microtubular ribbon,remain in the egg cytoplasm after nuclear fusion.Bell and Fasciatietal.suggested that the cytoskeletal apparatuses served as a guiding structure inside which the male nucleus twisted into the egg nucleus[6,8].InAthyriumthe presumptive actin filaments in the anterior end of the spermatozoid[8]were not seen in present investigation.
3.4 Organelles and nuclear evagination in the zygote
The male organelles of the spermatozoid including the microtubular apparatuses and the male mitochondria,undoubtedly enter the egg cytoplasm[6-10].Only the microtubular apparatuses of the spermatozoid,including flagella and microtubular ribbon,were easily recognized in the egg cytoplasm.They per-sist more that 3hand become much clearer after karyogamy as observed in previous reports[6-10].Hoffman and Vaughn indicated that the acetylation and post-translational modifications possibly make the microtubules the most stable structure in the cytoplasm[16].The male mitochondria in the zygote ofCeratopterisare digested between 6-9hafter fertilization[3].But,the male mitochondria can not be identified in present observations.
The egg nucleus ofPteridiumproduces extensive evaginations containing osmiophilic bodies[5,11].The fate of the nuclear evaginations is puzzling[5].The present investigation shows that the nuclear evaginations once become undistinguishable during karyogamy,but become clearly visible again after karyogamy.The nuclear evaginations persist about 12 h of fertilization,then most of them are separated from the nuclear body and enter the zygote cytoplasm.At about 24hafter fertilization,no visible evaginations are seen in the zygote cytoplasm.Bell and Duckett suggested that the nuclear evaginations either disintegrate or differentiate into some component of the cytoplasm[5].A further investigation is needed for illustrating the fate of the nuclear evaginations.
In the angiosperm it was also suggested that the polarity is important or prerequisite for the development of the zygote[17].InC.thalictroides,the zygote changes its vertical polarity into a horizontal polarity by the rearrangement of the organelles[3].The present investigation shows that a large vacuole lies in the lateral side of the zygote.Foundation of the polarity of the zygote is undoubtedly prepared for the first division which is always obliquevertical to the apical notch of the gametophyte.
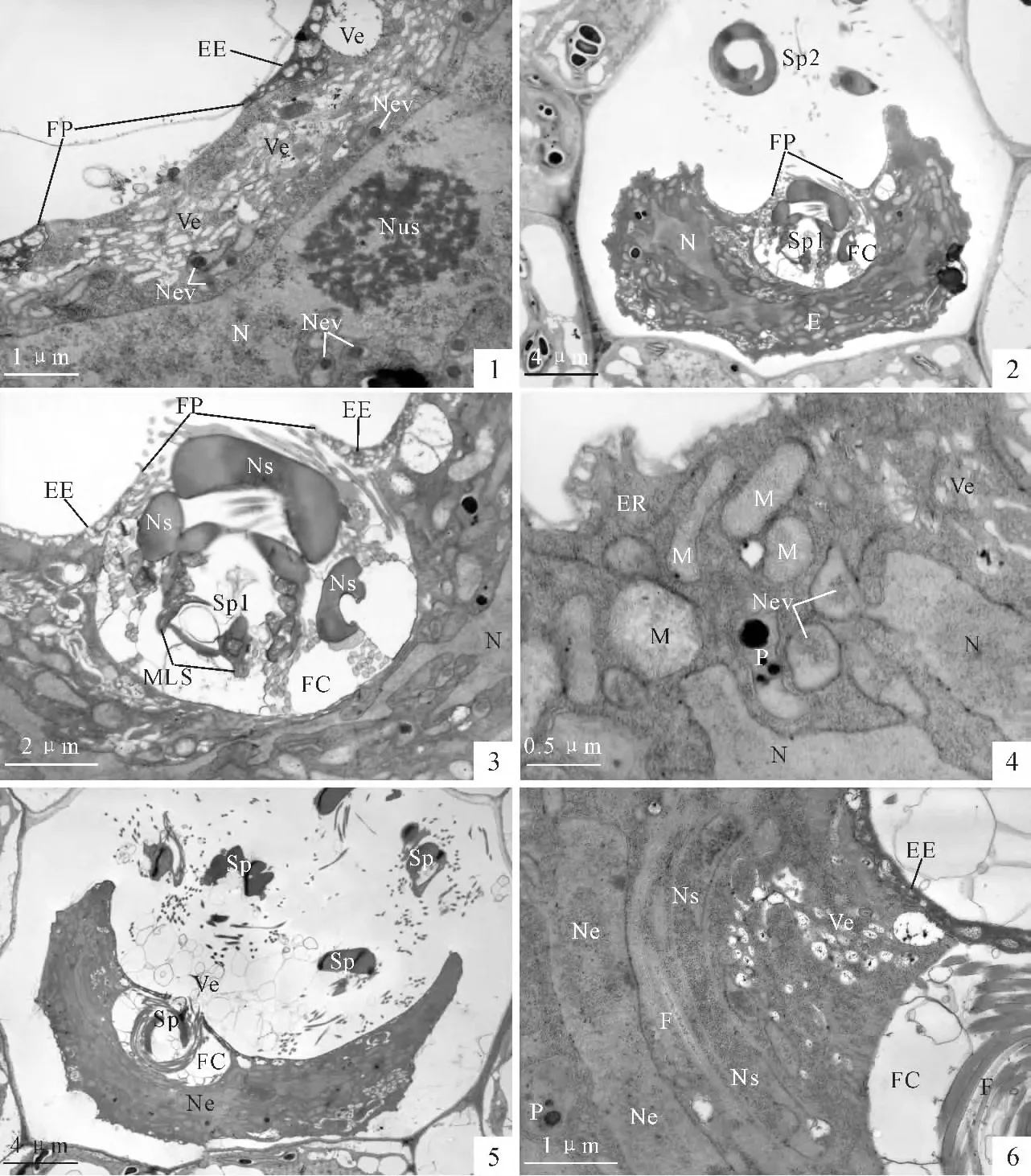
PlateⅠ Egg penetration in Pteridium aquilinum (within 10min of fertilization)
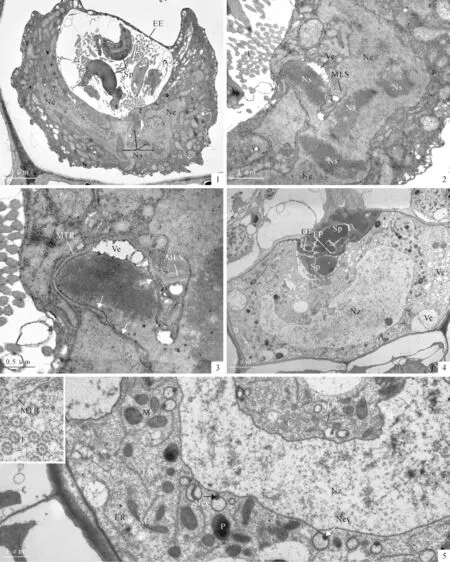
PlateⅡ Nuclear fusion and egg expanding(30min-3hafter fertilization)
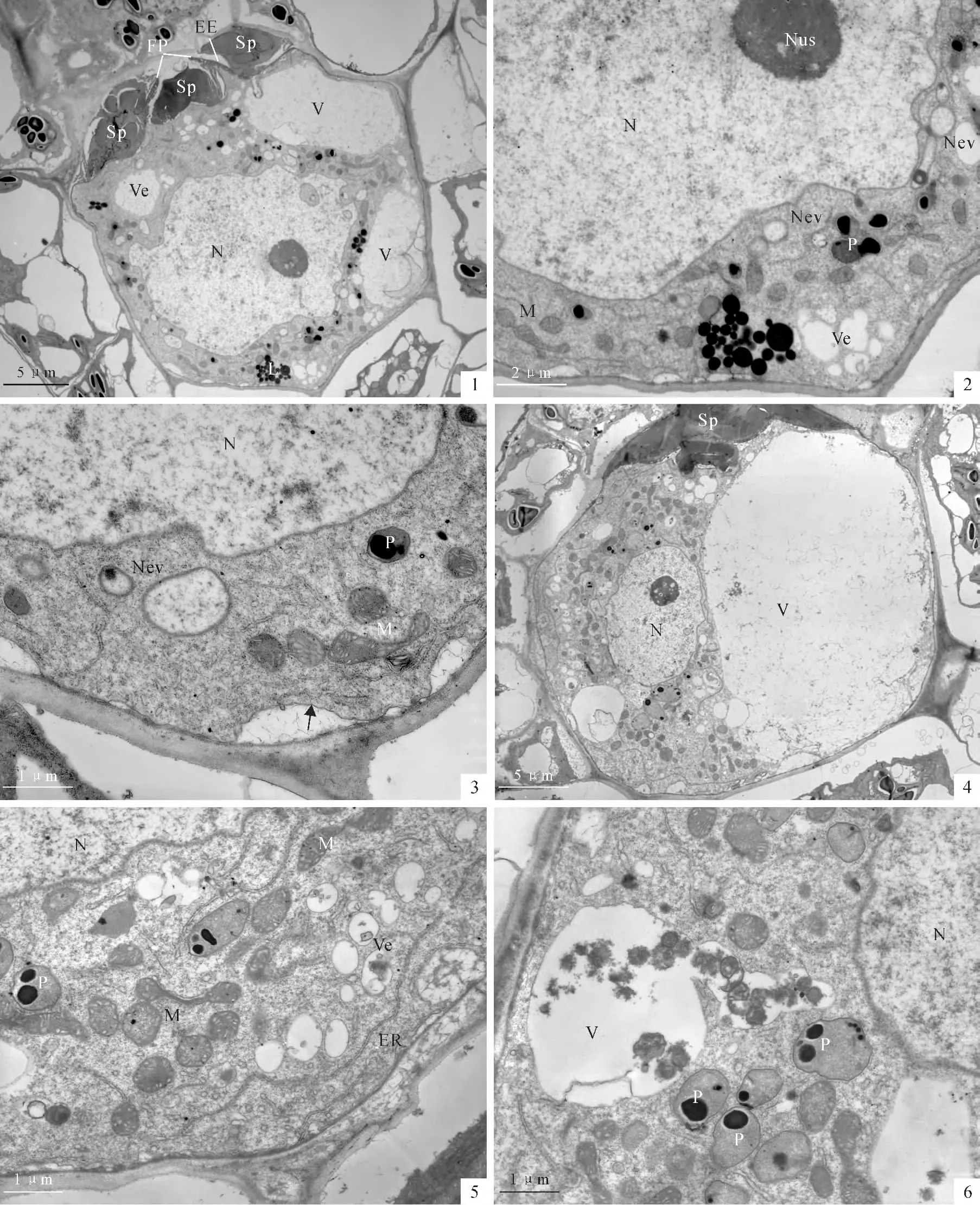
PlateⅢ Zygote polarity establishment(12-24hafter fertilization)
[1] ANTOINE A F,DUMAS C,FAURE J E,etal.Egg activation in flowering plants[J].SexualPlantReproduction,2001,14:21-26.
[2] CAO J G,WANG Q X,BAO W M.Formation of the fertilization pore during oogenesis of the fernCeratopteristhalictroides[J].Journal ofIntegrativePlantBiology,2010,52(6):518-527.
[3] CAO J G,WANG Q X,YANG N Y,etal.Cytological events during zygote formation of the fernCeratopteristhalictroides[J].Journalof IntegrativePlantBiology,2010,52(3):254-264.
[4] DUCKETT J G,BELL P R.Studies on fertilization in archegoniate plants.Ⅱ.Egg penetration inPteridiumaquilinum(L.)Kuhn[J].Cytobiologie,1972,6(1):35-50.
[5] BELL P R,DUCKETT J G.Gametogenesis and fertilization inPteridium[J].BotanicalJournaloftheLinneanSociety,1976,73:47-78.
[6] BELL P R.Observations on the male nucleus during fertilization in the fernPteridiumaquilinum[J].JournalofCellScience,1975,17:141-153.
[7] MYLES D G.The fine structure of fertilization in the fernMarsileavestita[J].JournalofCellScience,1978,30:265-281.
[8] FASCIATI R,SCHNELLER J,JENNI V,etal.Fertilization in the fernAthyriumfilix-femina(Pterophyta):Ⅱ.Ultrastructure[J].CryptogamicBotany,1994,4:356-367.
[9] LOPEZ-SMITH R,RENZAGLIA.Sperm cell architecture,insemination,and fertilization in the model fern,Ceratopterisrichardii[J].SexualPlantReproduction,2008,21:153-167.
[10] CAO J G,YANG N Y,WANG Q X.Ultrastructure of the mature egg and fertilization in the fernCeratopteristhalictroides(L.)Brongn[J].JournalofIntegrativePlantBiology,2009,51(3):243-250.
[11] CAO J G,DAI X L,WANG Q X.Ultrastructural and cytochemical studies on oogenesis of the fernPteridiumaquilinum[J].Sexual PlantReproduction,2012,25:147-156.
[12] ATKINSON L.R.Cytology[M]//VERDOOM F.Manual of Pteridology.Martinus Nijhoff.The Hague,1938:196.
[13] GARDNER A J,EVANS J P.Mammalian membrane block to polyspermy:new insights into how mammalian eggs prevent fertilization by multiple sperm[J].Reproduction,FertilityandDevelopment,2006,18(1-2):53-61.
[14] MISAMORE M J,GUPTA S,SNELL W J.TheChlamydomonasFus1protein Is present on the mating type plus fusion organelle and required for a critical membrane adhesion event during fusion with minus gametes[J].MolecularBiologyoftheCell,2003,14(6):2 530-2 542.
[15] SPIELMAN M,SCOTT R J.Polyspermy barriers in plants:from preventing to promoting fertilization[J].SexualPlantReproduction,2008,21:53-65.
[16] HOFFMAN J C,VAUGHN K C.Post-translational tubulin modifications in spermatogeneous cells of the PteridophyteCeratopterisrichardii[J].Protoplasma,1995,186(3-4):169-182.
[17] MANSFELD S G,BRIARTY L G.Early embryogenesis inArabidopsisthaliana.Ⅱ.The developing embryo[J].CanadianJournalof Botany,1991,69(3):461-476.
