柱花草种间亲缘关系SSR分析
2015-06-24丁西朋罗小燕张龙王文强白昌军
丁西朋,罗小燕,张龙,王文强,白昌军
(中国热带农业科学院热带作物品种资源研究所,海南 儋州 571737)
柱花草种间亲缘关系SSR分析
丁西朋,罗小燕,张龙,王文强,白昌军
(中国热带农业科学院热带作物品种资源研究所,海南 儋州 571737)
为研究柱花草(Stylosanthesspp.)种间的亲缘关系,本研究利用23个SSR标记对柱花草属14个种的33份材料的遗传关系进行了研究。结果表明,23个SSR标记在不同柱花草种间均具有多态性,共检测到等位基因138个,每个标记可检测到4~10个等位基因,平均6个;每个标记的多态性信息含量在0.501~0.830,平均为0.700。聚类分析和主成分分析结果表明, 33份供试柱花草种质可分为5类,根据包含的基因组类型分别命名为AB类、AC类、J类、DE类、G类。毛叶柱花草(S.subsericea)和有钩柱花草(S.hamata)(I)的亲缘关系最近,有钩柱花草(I)与头状柱花草(S.capitata)的遗传关系较远。
柱花草;SSR;亲缘关系;聚类分析;主成分分析
柱花草属(Stylosanthesspp.)为多年生豆科植物,原产于拉丁美洲,具有适应性强、耐贫瘠、抗旱、饲草产量高、营养价值高等优点,兼具改良土壤、水土保持的功效,是全球热带及亚热带地区广泛种植的优良牧草之一[1]。柱花草属于自花授粉,且在开花之前已经授粉[2]。柱花草属包含约50个种或亚种,常用的栽培品种主要来自圭亚那柱花草(Styosanthesguianensis)、有钩柱花草(S.hamata)、糙柱花草(S.scabra)、灌木柱花草(S.seabrana)、矮柱花草(S.humilis)和头状柱花草(S.macrocephala)等[3]。大部分柱花草为二倍体(2n=2x=20),也有部分为多倍体(2n=4x=40或2n=6x=60)[4]。Liu等[5]鉴定出柱花草属的10个基本基因组类型,并命名为A~J。柱花草传统的分类方法主要基于花果形态,早在1938年,Vogel根据荚果基部退化花轴的有无和小苞片的数量将柱花草划分为Styposanthes(包含二倍体和多倍体)和Stylosanthes(完全为二倍体)两大分支[6]。但是柱花草属中有些种和亚种间的形态特征非常相近,形态学鉴定非常困难,目前还存在许多同种异名或同名异种的现象,近年来,不断有新种被发现或重新定义[7-9]。国内外有学者利用STS、RFLP及ITS等分子标记和叶绿体DNA序列差异对柱花草属种间分类进行了研究,但由于所用手段和研究材料的不同,研究结果也存在一定差异[5,10-11],所以深入研究柱花草属种间的亲缘关系对柱花草种质资源研究与利用尤为重要。
SSR标记因具有数量丰富、多态性高、共显性、操作简单等多个优点,在遗传学研究中作为一种重要的分子标记技术,已经被广泛于植物研究的诸多领域[12]。在牧草上,SSR标记已经在柱花草[13]、苜蓿(Medicagosativa)[14]、黑麦草(Loliumperenne)[15]、羊草(Leymuschinensis)[16]、鸭茅(Dactylisglomerata)[17]、三叶草(Trifoliumpratense)[18]、百喜草(Paspalumnotatum)[16]等草种上进行了遗传图谱构建、倍性鉴定、遗传多样性分析、父系分析、QTL鉴定等方面的研究[16]。近年,柱花草SSR引物的不断开发和应用,为柱花草种质资源及遗传改良提供准确了可靠的标记[4]。本研究利用已报道的部分SSR标记,对柱花草种间的亲缘关系进行分析,以进一步为其种间的分类提供更多的分子水平上的依据。
1 材料与方法
1.1 试验材料
本研究所用33份柱花草种质来自柱花草属的14个种,均由中国热带农业科学院热带作物品种资源研究所牧草中心收集。其中7份来自澳大利亚联邦科学与工业研究组织(CSIRO),12份来自中国热带农业科学院(CATAS)热带作物品种资源研究所,14份引进于哥伦比亚国际热带农业中心(CIAT)(表1)。由于有钩柱花草包括四倍体和二倍体两种类型,所以将其分为两个种群,分别命名为有钩柱花草(I)和有钩柱花草(II)。所有供试材料均种植在中国热带农业科学院热带作物品种资源研究所热带牧草研究中心基地,按照日常大田管理办法进行管理。
1.2 DNA的提取及浓度检测
在柱花草营养生长期,采集柱花草新鲜幼嫩叶片置于液氮中,带回实验室于-80 ℃超低温冰箱保存备用。按照中国热带农业科学院热带作物品种资源研究所热带牧草研究中心实验室改良CTAB法提取柱花草总DNA[19]。利用NanoVue超微量分光光度计(GE Healthcare)检测完全溶解的DNA样品的浓度与A260/280、A260/230,然后将DNA样品浓度按照50 ng·μL-1配置好,同时利用1%琼脂糖凝胶电泳检测DNA样品。
1.3 SSR引物、PCR扩增及产物检测
选用中国热带农业科学院热带作物品种资源研究所热带牧草研究中心实验室已经筛选过在8个不同柱花草种中都能有效扩增,且条带清晰、存在多态性的已报道的SSR标记进行柱花草种间亲缘关系分析(表2)[19],引物由上海生工生物工程公司合成。试验中Taq DNA聚合酶、dNTPs、10 × PCR Buffer(含Mg2+)及DL2000均购自TaKaRa公司。
利用德国Biometra公司的TGRADient型PCR仪进行PCR扩增,采用20 μL反应体系,包括2.0 μL 10×PCR Buffer(含Mg2+),1.0 μL模板DNA(50 ng·μL-1),0.4 μL正向引物(10 μmol·L-1),0.4 μL反向引物(10 μmol·L-1),1.2 μL dNTPs(2.5 mmol·L-1),0.3 μL Taq DNA 聚合酶(5 U·μL-1),剩余体积用ddH2O补足。PCR反应扩增程序为94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火45 s,72 ℃延伸45 s,循环35次;最后72 ℃延伸10 min,4 ℃保存。扩增产物用8%聚丙烯酰胺凝胶电泳(Polyacrylamide Gel Electrophoresis,PAGE)电泳后,采用硝酸银法染色,最后照相记录[19]。
1.4 数据分析
扩增产物片段大小以DL2000 DNA marker作为分子量参考来读取,对大小合适、清晰且易于辨认的谱带采用“0,1”系统记录其位置,有条带记为“1”, 没有记为“0”。所记录的原始数据汇总到Excel表格中并形成二进制矩阵,用于扩增条带的多态性比较和聚类分析。利用NTSYS-pc 2.10e软件分析柱花草种质及种间的遗传距离和相似系数(Genetic Similarity,GS),然后利用GS值采用非加权组平均法(UPGMA)对供试材料进行聚类分析和主成分分析[20],并通过FreeTree软件[21]采用1 000次重抽样对聚类图进行bootstrap分析。利用POPGENE 1.32软件计算每个标记的遗传多样性参数,包括等位基因数(Number of Alleles,Na)、每个等位基因的频率、观察杂合度(Observed Heterozygosity,Ho)、期望杂合度(Excepted Heterozygosity,He)和Shannon多态性指数(I)[22]。根据Botstein等[23]方法,利用SSR标记每个等位基因频率计算其多态性信息含量(Polymorphic Information Content,PIC),计算公式如下:
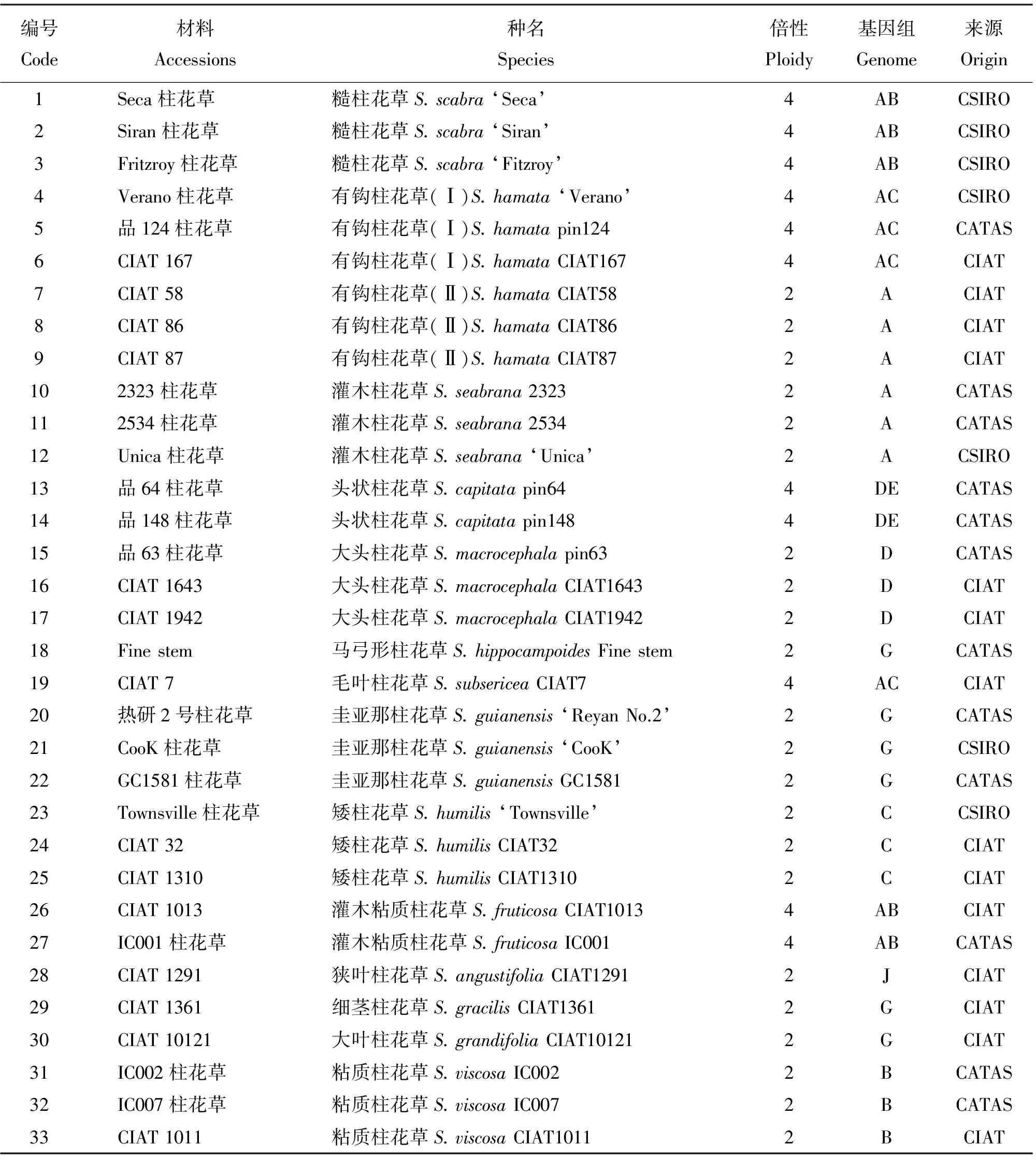
表1 供试的33份柱花草种质Table 1 33 Stylosanthes accessions used in this study
注:CSIRO, 澳大利亚联邦科学与工业研究组织;CATAS,中国热带农业科学院;CIAT,哥伦比亚国际热带农业中心。
Note: CSIRO, Commonwealth Scientific and Industrial Research Organisation; CATAS, Chinese Academy of Tropical Agricultural Sciences; CIAT, International Center for Tropical Agriculture.

式中,n表示每个标记检测到的等位基因数目,Pi、Pj表示第i、j个等位基因在供试材料中出现的频率。
2 结果与分析
2.1 柱花草种间SSR扩增特点
选用的23对SSR引物在33份柱花草材料中均具有多态性,共检测到138个等位基因,每个标记检测到的等位基因数为4~10个,平均6个(表2)。23个SSR标记的观察杂合度(Ho)的变化范围为0.000~0.667,平均值为0.225,而期望杂合度(He)的变化范围为0.572~0.861,平均值为0.750(表2)。标记SSR23的观察杂合度最高,标记SSR5的预期杂合度最大。各SSR标记的多态信息含量(PIC)值变化范围为0.501~0.830,平均为0.700。各SSR标记的Shannon多样性指数(I)变异范围为1.012~2.039,其平均值为1.516。多态信息含量(PIC)值和Shannon多样性指数(I)都以标记SSR5最大,标记SSR101最小,表明SSR5具有较高的多态性检测效率。
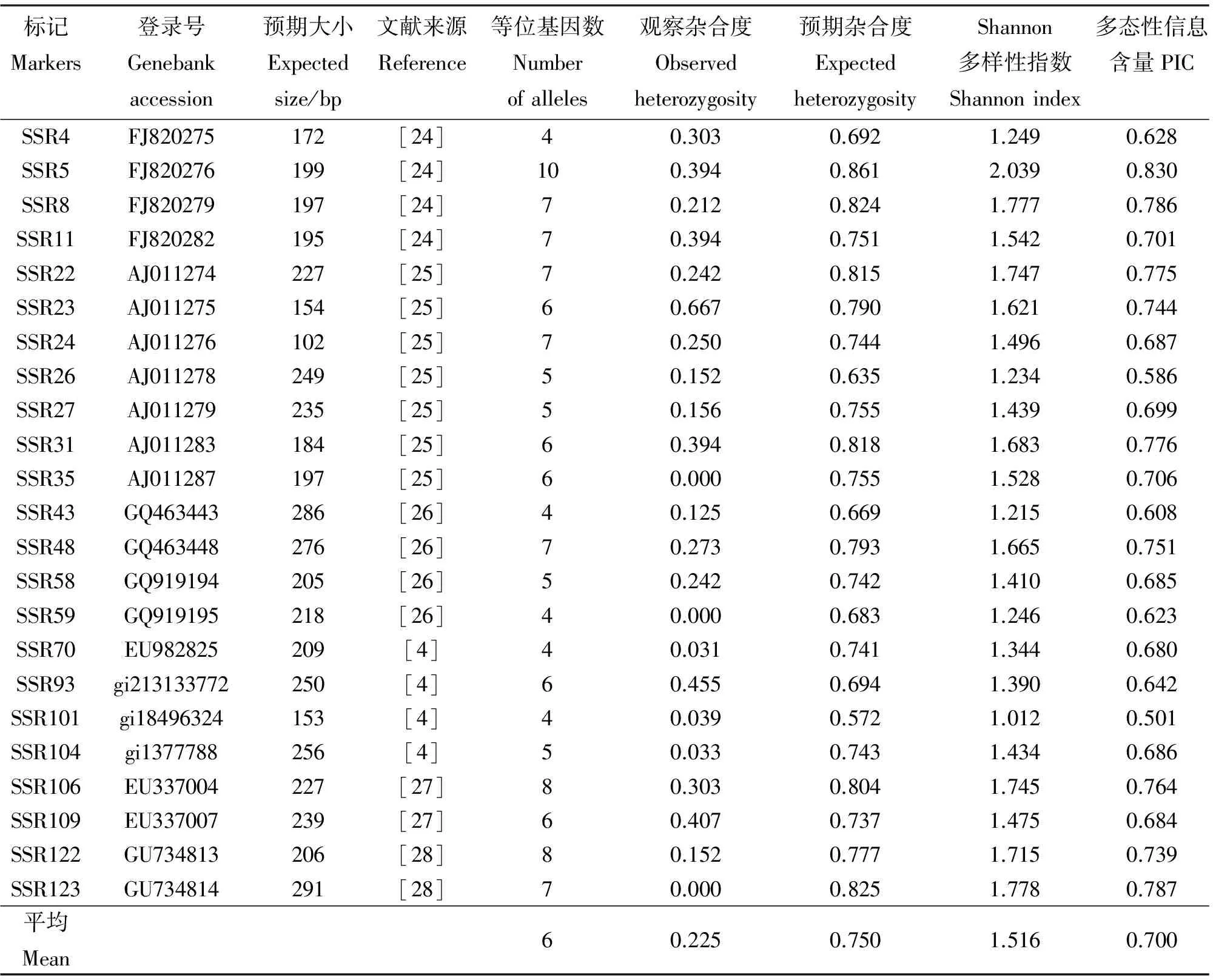
表2 23对SSR引物信息及特征分析Table 2 Information and characterization of 23 SSR markers
2.2 柱花草种质间的遗传多样性与聚类分析
利用NTSYS-pc 2.10e分析软件,对供试柱花草种质资源的进行遗传多样性,结果表明,33份种质资源间的遗传相似系数GS变化范围为0.557~0.993,平均为0.715。其中,CIAT 167有钩柱花草与品148头状柱花草的遗传相似性最小。基于23对SSR引物的UPGMA聚类分析表明,在相似系数0.760处,33份柱花草种质资源可以明显地被分为5类(图1):3份糙柱花草、2份灌木粘质柱花草、3份灌木柱花草和3份粘质柱花草被归为一类,由于他们的基因组类型为A、B或AB类,所以命名为AB类;6份有钩柱花草、1份毛叶柱花草和3份矮柱花草被归为一类,由于他们的基因组类型为AC或C类,所以命名为AC类;含有J类基因组的CIAT1291狭叶柱花草单独分为一类;3份头状柱花草(DD)和2份大头柱花草(DDEE)被归为DE类;剩下的3份圭亚那柱花草(GG)、Fine stem马弓形柱花草(GG)、CIAT10121大叶柱花草(GG)和CIAT1361细茎柱花草(GG)被归为G类。
2.3 主成分分析
基于SSR数据矩阵,对33份供试柱花草材料进行主成分分析,前3个主成分解释的变异分别为72.51%、6.32%和4.25%,合计占总变异的83.08%。根据前3个主成分制作33份供试材料的三维散点分布图(图2),位置相靠近者表示关系密切,远离者表示关系疏远。比较图2和图1发现,主成分分析和聚类分析结果基本一致,同样将33份柱花草种质分为5类,其中AC类和G类比较集中,而AB类和DE类相对分散。相对而言,主成分分析(图2)比聚类分析(图1)能更直观地揭示不同材料间的亲缘关系。

图1 柱花草种间SSR分析聚类图Fig.1 Dendrogram of Stylosanthes species based on SSR information

图2 33份材料第1、2、3主成分三维散点图Fig.2 3D-scatterplot based on the first, second and third principal components of 33 materials
注:图中数字表示材料编号同表1。
Note: The numbers for materials in the figure are corresponding to the numbers of accession name given in Table 1.
2.4 柱花草不同物种间的亲缘关系分析
如果将33份供试柱花草资源按种可分为14个不同的组,但考虑到不同倍性对亲缘关系的影响,将有钩柱花草分为四倍体有钩柱花草和二倍体有钩柱花草,分别命名为有钩柱花草(Ⅰ)和有钩柱花草(Ⅱ)。利用SSR数据矩阵对这15个种群进行相似系数和遗传距离分析发现(表3),有钩柱花草(Ⅰ)和毛叶柱花草的相似系数最大,为0.902,大于有钩柱花草(Ⅰ)和有钩柱花草(Ⅱ)的相似系数(0.872)。头状柱花草除了与大头柱花草的相似系数较高(0.824)外,与其他种群的相似系数均较低。有
钩柱花草(Ⅰ)和头状柱花草的相似系数最小,为0.575。遗传距离分析结果与相似系数分析结果相反,有钩柱花草(Ⅰ)和毛叶柱花草之间的遗传距离最小,为0.098,而有钩柱花草(Ⅰ)和头状柱花草之间的遗传距离最大,为0.425。
3 讨论与结论
分子标记是作物遗传学研究和分子标记辅助育种的重要工具,SSR标记相对于其他基于PCR的分子标记,具有数量丰富、多态性高、共显性、操作简单等多个优点,这使SSR技术在多个领域得以应用。目前,SSR技术主要应用于柱花草种内遗传多样性分析及和核心种质的建立。蒋昌顺等[29]利用18个SSR标记成功地将42份圭亚那柱花草(S.guianensis)种质分为6类,并证明其效果好于RAPD标记。Santos-Garcia等[30]利用20个SSR标记对150份圭亚那柱花草种质资源进行了遗传多样性分析。Chandra等[4]开发了133个柱花草SSR标记,并用其中41个对20份灌木柱花草进行了遗传多样性分析。Santos-Garcia等[31]用13个SSR标记对134份大头柱花草种质进行了基因型分析最终建立了含23份种质的大头柱花草核心种质,同时用15个SSR标记对192份头状柱花草种质进行了基因型分析并最终建立了含13份种质的头状柱花草核心种质。本研究通过分析来自柱花草属14个种的33份供试柱花草材料的遗传多样性,首次利用SSR标记分析了柱花草种间的亲缘关系。23个SSR标记在33份供试材料中均有多态性,共检测到的等位基因138个,每个标记可检测到4~10个等位基因,平均6个。利用SSR标记能将每份供试柱花草很好地分开,同种柱花草种质资源聚在一起,相互间相似系数较高,而不同柱花草种间相互分开,遗传距离较远(图1,表3),且该分类结果与植物学分类结果较一致。因此,SSR标记不仅能用于柱花草种质遗传多样性的分析,还是柱花草种间亲缘分析的有效方法。
柱花草传统的分类方法主要是基于花果形态的差异,但由于柱花草分布广泛、种类繁多、花果形态复杂多变,使得柱花草属内种和亚种的形态学上鉴
定非常难,种的概念及其特性目前还存在多种观点,分子标记方法对阐述物种间遗传关系提供了有力的手段[32]。国外有学者利用STS、RFLP及ITS等分子标记和叶绿体DNA序列差异对柱花草属种间亲缘关系进行了研究[5,10-11]。本研究利用23个SSR标记,对柱花草属14个种亲缘关系进行分析,结果表明有钩柱花草(Ⅰ)和毛叶柱花草的相似系数最大,为0.902,大于有钩柱花草(Ⅰ)和有钩柱花草(Ⅱ)的相似系数(0.872),和形态学上的分类不一致,这可能是因为有钩柱花草(Ⅰ)和毛叶柱花草拥有相同的倍性及基因组类型。有钩柱花草(Ⅰ)和头状柱花草的相似系数最小,为0.575,表明它们之间的亲缘关系最远。来自圭亚那柱花草复合体的圭亚那柱花草、大叶柱花草、马弓形柱花草和细茎柱花草亲缘关系较近,相互间相似系数较大,为0.785~0.846,同时被归为G类(图1、2),这一结果与用其他分子标记分析的结果一致[5,10-11]。头状柱花草和大头柱花草都含有D类基因组,两者相似系数较高,为0.824,但两者与其他种群的相似系数均较低,这与Gilles和Abbott[11]利用叶绿体DNA序列差异分析结果吻合。本研究的SSR分析结果还为判断四倍体柱花草的祖先提供了证据:糙柱花草(AABB)、灌木柱花草(AA)和粘质柱花草(BB)被归为AB类,进一步支持了灌木柱花草和粘质柱花草是糙柱花草祖先的观点[33]。同样,有钩柱花草(Ⅰ)(AACC)、有钩柱花草(Ⅱ)(AA)和矮柱花草(CC)被归为AC类支持了二倍体有钩柱花草和矮柱花草是四倍体有钩柱花草祖先的说法[34]。
[1] 蒋昌顺.柱花草的研究进展[J].热带作物学报,2005,26(4):104-108.
[2] 古艳,罗丽娟.热研2号柱花草传粉受精作用的研究[J].热带农业科学,2010,30(3):25-29.
[3] Maass B L,Sawkins M.High-yielding Anthracnose-resistantStylosanthesfor Agricultural Systems[M].Australia:Australian Centre for International Agricultural Research (ACIAR),2004:9-26.
[4] Chandra A,Tiwari K K,Nagaich D,Dubey N,Kumar S,Roy A K.Development and characterization of microsatellite markers from tropical forageStylosanthesspecies and analysis of genetic variability and cross-species transferability[J].Genome,2011,54(12):1016-1028.
[5] Liu C J,Musial J M,Thomas B D.Genetic relationships amongStylosanthesspecies revealed by RFLP and STS analyses[J].Theoretical and Applied Genetics,1999,99(7-8):1179-1186.
[6] Mannetje L T.The Biology and Agronomy ofStylosanthes[M].Sydney:Academic Press,1984:1-21.
[7] Sousa Costa N M,Van den Berg C.A new species ofStylosanthesSw.(Leguminosae-Papilionoideae) from Paraná,Brazil[J].Neodiversity,2009(4):9-13.
[8] Calles T,Schultze-Kraft R.Stylosanthes falconensis (Leguminosae,Papilionoideae,Dalbergieae),a new species endemic to Venezuela[J].Kew Bulletin,2010,65(1):73-76.
[9] Calles T,Schultze-Kraft R.Re-establishment ofStylosanthesgracilis(Leguminosae) at species level[J].Kew Bulletin,2010,65(2):233-240.
[10] Vander Stappen J,Marant S,Volckaert G.Molecular characterization and phylogenetic utility of the rDNA external transcribed spacer region inStylosanthes(Fabaceae)[J].Theoretical and Applied Genetics,2003,107(2):291-298.
[11] Gillies A C M,Abbott R J.Phylogenetic relationships in the genusStylosanthes(Leguminosae) based upon chloroplast DNA variation[J].Plant Systematics and Evolution,1996,200(3-4):193-211.
[12] Kalia R K,Rai M K,Kalia S,Singh R,Dhawan A K.Microsatellite markers:An overview of the recent progress in plants[J].Euphytica,2011,177(3):309-334.
[13] 韩蓉蓉,史亮涛,刘国道,何光熊,文亦芾,曾黎琼,金杰,王檀,罗富成.7个柱花草品种的AFLP和SSR遗传多样性分析[J].中国农学通报,2014,30(18):271-276.
[14] Wang Z,Yu G,Shi B,Wang X,Qiang H,Gao H.Development and characterization of simple sequence repeat (SSR) markers based on RNA-sequencing ofMedicagosativaand in silico mapping onto theM.truncatulagenome[J].PLoS One,2014,9(3):e92029.
[15] Farrell J D,Byrne S,Paina C,Asp T.De novo assembly of the perennial ryegrass transcriptome using an RNA-seq strategy[J].PLoS One,2014,9(8):e103567.
[16] 李志勇,孙启忠,李鸿雁,王小丽,师文贵,戴军.分子标记技术在牧草种质资源研究中的应用[J].草原与草坪,2010,30(5):91-96.
[17] Xie W,Zhang X,Cai H,Huang L,Peng Y,Ma X.Genetic maps of SSR and SRAP markers in diploid orchardgrass (DactylisglomerataL.) using the pseudo-testcross strategy[J].Genome,2011,54(3):212-21.
[18] Riday H,Krohn A L.Genetic map-based location of the red clover (TrifoliumpratenseL.) gametophytic self-incompatibility locus[J].Theoretical and Applied Genetics,2010,121(4):761-767.
[19] 张龙,丁西朋,严琳玲,胡珊娜,白昌军.8种柱花草属牧草SSR-PCR反应体系优化及引物筛选[J].草业科学,2014,31(2):233-242.
[20] Rohlf F J.NTSYS-pc Numerical taxonomy and multivariate analysis system[CP].version 2.10.Exeter Software,New York:Department of Ecology and Evolution State University of New York Stany Brook,2000.
[21] Hampl V,Pavlicek A,Flegr J.Construction and bootstrap analysis of DNA fingerprinting-based phylogenetic trees with the freeware program Free Tree:Application to trichomonad parasites[J].International Journal of Systematic and Evolutionary Microbiology,2001,51:731-735.
[22] Yeh F C,Yang R C,Boyle T.POPGENE,version 1.32:The User Friendly Software For Population Genetic Analysis[M].Edmonton,AB,Canada:Molecular Biology and Biotechnology Centre,University of Alberta,1999.
[23] Botstein D,White R L,Skolnick M,Davis R W.Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J].The American Journal of Human Genetics,1980,32(3):314-331.
[24] Santos M O,Karia C T,Resende R M,Chiari L,Jungmann L,Zucchi M I,Souza A P.Isolation and characterization of microsatellite loci in the tropical forage legumeStylosanthesguianensis(Aubl.) Sw.[J].Conservation Genetics Resources,2009,1:43-46.
[25] Vander Stappen J,Weltjens I,Volckaert G.Microsatellite markers inStylosanthesguianensis[J].Molecular Ecology,1999,8(3):514-517.
[26] Santos M O,Sassaki R P,Ferreira T H S,Resende R M S,Chiari L,Karia C T,Faleiro F G,Jungmann L,Zucchi M I,Souza A P.Polymorphic microsatellite loci forStylosanthesmacrocephalaFerr.et Costa,a tropical forage legume[J].Conservation Genetics Resources,2009,1:481-485.
[27] Santos M O,Sassaki R P,Chiari L,Resende R M,Souza A P.Isolation and characterization of microsatellite loci in tropical forageStylosanthescapitataVogel[J].Molecular Ecology Resources,2009,9(1):192-194.
[28] Billotte N,Lagoda P J R,Risterucci A M,Baurens F C.Microsatellite-enriched libraries:Applied methodology for the development of SSR markers in tropical crops[J].Fruits,1999,54:277-288.
[29] 蒋昌顺,马欣荣,邹冬梅,张义正.应用微卫星标记分析柱花草的遗传多样性[J].高技术通讯,2004(4):25-30
[30] Santos-Garcia M O,Karia C T,Resende R M,Chiari L,Vieira M L,Zucchi M I,Souza A P.Identification ofStylosanthesguianensisvarieties using molecular genetic analysis[J].AoB Plants,2012:doi:10.1093/aobpla/plsool.
[31] Santos-Garcia M O,Toledo-Silva G,Sassaki R P,Ferreira T H,Resende R M,Chiari L,Karia C T,Carvalho M A,Faleiro F G,Zucchi M I, Souza A P.Using genetic diversity information to establish core collections ofStylosanthescapitataandStylosanthesmacrocephala[J].Genetic and Molecular Biology,2012,35(4):847-861.
[32] 唐燕琼,吴紫云,刘国道,易克贤.柱花草种质资源研究进展[J].植物学报,2009,44(6):752-762.
[33] Liu C J,Musial J M.Stylosanthessp.aft.S.scabra:A putative diploid progenitor ofStylosanthesscabra(Fabaceae)[J].Plant Systematics and Evolution,1997,208:99-105.
[34] Curtis M D,Cameron D F,Manors J M.Molecular evidence that diploidStylosantheshumilisand diploidStylosantheshamataare progenitors of allotetraploidStylosantheshamata[J].Genome,1995,38:344-348.
(责任编辑 王芳)
Genetic relationships amongStylosanthesspecies revealed by SSR analysis
DING Xi-peng, LUO Xiao-yan, ZHANG Long, WANG Wen-qiang, BAI Chang-jun
(Institute of Tropical Crops Genetic Resources, CATAS, Danzhou 571737, China)
To investigate genetic relationships amongStylosanthesspecies, a total of 33 accessions from 14Stylosanthesspecies were studied using 23 simple sequence rpeats (SSR) markers. All 23 SSR markers were polymorphic among 33Stylosanthesaccessions. A total of 138 alleles were generated by 23 SSR markers among differentStylosanthesaccessions. The number of alleles per SSR marker ranged from 4 to 10 with an average of 6 alleles. The polymorphic information content per SSR marker varied from 0.501 to 0.830, with an average of 0.700. Based on the results of analysis of cluster and principal components, the 33Stylosanthesaccessions were divided into 5 separate clusters, which were named to be cluster AB, cluster AC, cluster J, cluster DE and cluster G based on genome contained. The highest and lowest genetic relations were presented betweenS.subsericeaandS.hamata(I),S.hamata(I) andS.capitata, respectively.
Stylosanthes; SSR markers; genetic relationship; cluster analysis; principal components analysis
BAI Chang-jun E-mail: baichangjun@126.com
10.11829j.issn.1001-0629.2015-0155
2015-03-19 接受日期:2015-06-01
海南省自然科学基金(313067);中央级公益性科研院所基本科研业务费专项资金(中国热带农业科学院热带作物品种资源研究所)资助项目(1630032013002);现代产业体系牧草岗位科学家经费(CRAS-35);中南美洲种质资源收集与合作研究(2011DFB31690);农业部“948”重点项目“重要热带作物特异种质资源的引进”[2011-G13(5)]
丁西朋(1982-),男,河南封丘人,助理研究员,博士,主要从事热带牧草遗传育种研究。E-mail:xipding@163.com
白昌军(1967-),男,甘肃民勤人,研究员,博士,主要从事热带牧草种质资源和栽培利用研究。E-mail:baichangjun@126.com
S32
A
1001-0629(2015)10-1594-09*
丁西朋,罗小燕,张龙,王文强,白昌军.柱花草种间亲缘关系SSR分析[J].草业科学,2015,32(10):1594-1602.
DING Xi-peng,LUO Xiao-yan,ZHANG Long,WANG Wen-qiang,BAI Chang-jun.Genetic relationships amongStylosanthesspecies revealed by SSR analysis[J].Pratacultural Science,2015,32(10):1594-1602.
